Phenotypic Variation Patterns in Oecomys catherinae (Rodentia: Sigmodontinae): Craniodental Morphometric Analysis and Its Relationship with Latitudinal Variation in the Atlantic Forest and Cerrado Biomes
Paola Santos da Mata, Thiago dos Santos Cardoso, Cibele Rodrigues Bonvicino, Roberto do Val Vilela

TL;DR
This study explores how the skull shape of a rodent species varies across different Brazilian biomes and latitudes, suggesting environmental influences on physical traits.
Contribution
The study identifies biome-specific cranial adaptations in Oecomys catherinae and links them to ecological and latitudinal factors.
Findings
Cranial morphology differs significantly between the Atlantic Forest and Cerrado biomes.
Southern populations have slightly larger cranial dimensions, aligning with Bergmann’s rule.
Bony palate length is the most discriminating trait between biomes, possibly linked to diet.
Abstract
Understanding how animals adapt to natural environments reveals essential physical and behavioral traits for survival. We examined skull shape variation in the arboreal rodent Oecomys catherinae across two Brazilian biomes, the Atlantic Forest and Cerrado, and its relationship with latitude. Through cranial and dental measurements of 45 specimens from scientific collections, no significant differences were observed between males and females. However, cranial morphology differed between biomes, likely due to dietary adaptations and ecological pressures. Southern populations exhibited slightly larger cranial dimensions than northern counterparts, consistent with Bergmann’s rule (which posits that larger body sizes occur in colder climates). These results demonstrate how environmental pressures shape physical traits in small mammals. Further research is required to determine whether these…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
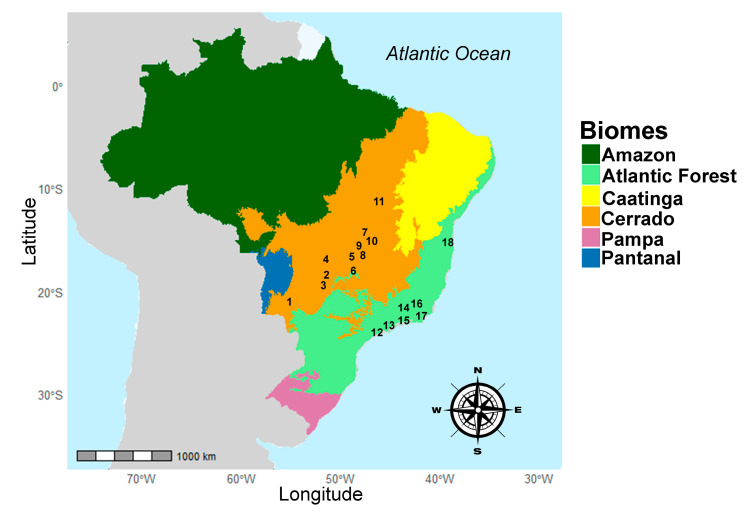 Figure 1
Figure 1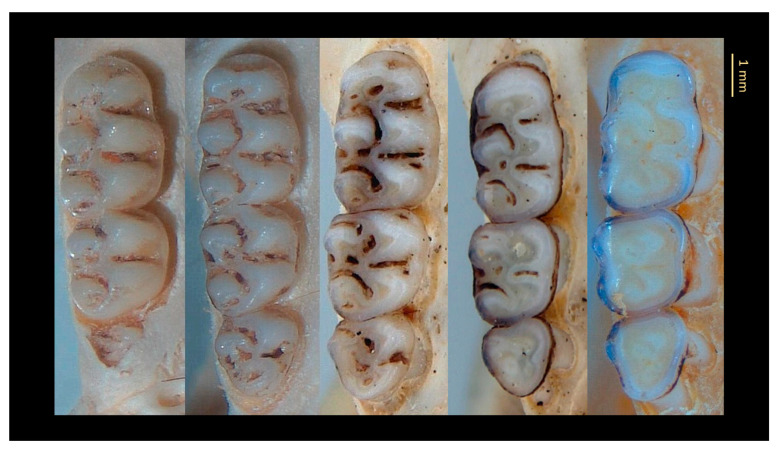 Figure 2
Figure 2 Figure 3
Figure 3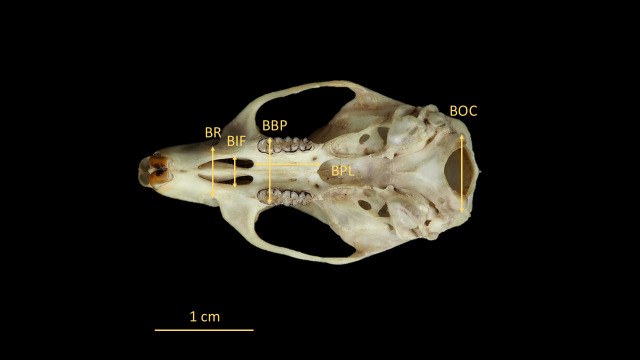 Figure 4
Figure 4 Figure 5
Figure 5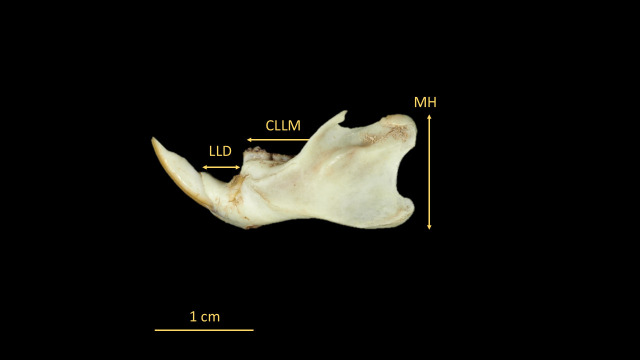 Figure 6
Figure 6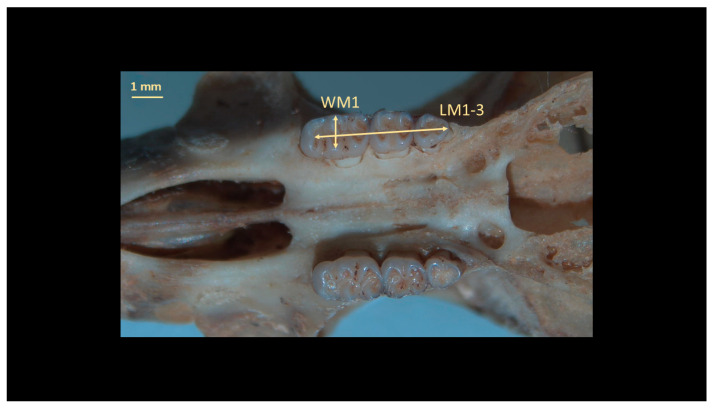 Figure 7
Figure 7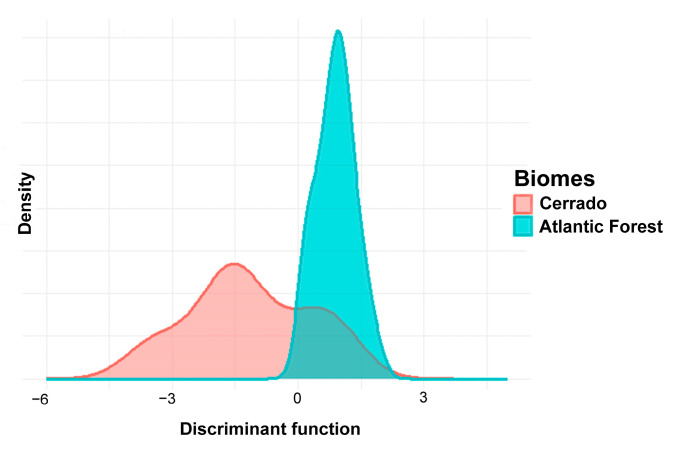 Figure 8
Figure 8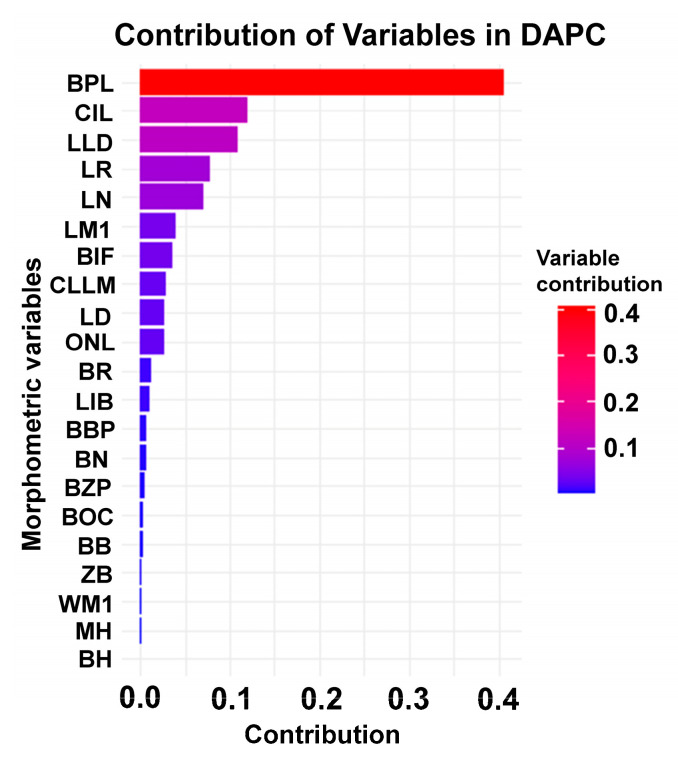 Figure 9
Figure 9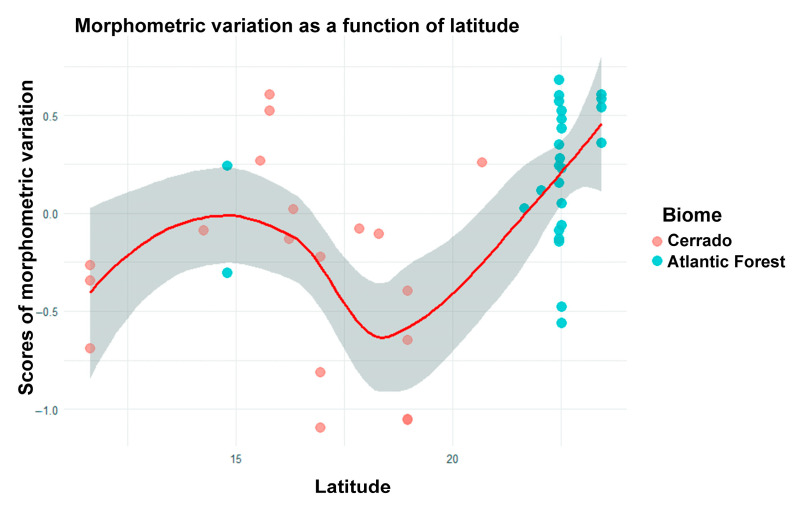 Figure 10
Figure 10- —Brazilian Coordination for the Improvement of Higher Education Personnel (CAPES)
- —Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro-FAPERJ
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEvolution and Paleontology Studies · Morphological variations and asymmetry · Wildlife Ecology and Conservation
1. Introduction
The genus Oecomys (Rodentia: Cricetidae) comprises semi-arboreal rodents with morphological and behavioral adaptations for efficiently utilizing arboreal strata in humid tropical forests [1]. These nocturnal species rely on vegetation for locomotion, feeding, and nesting, occupying both arboreal strata and the ground [2]. Morphological traits such as reduced limb proportions reflect this arboreal specialization, making them the most anatomically adapted Oryzomyini to this lifestyle [3].
The taxonomic classification of Oecomys is challenging due to high interspecific similarity, requiring integrative analyses of morphological and genetic traits [4,5]. This complexity has left many species unresolved, underscoring the need for further studies [6,7,8]. Additionally, the lack of long-term autecological studies represents a significant gap in understanding their biology and environmental interactions [2]. Among the species of Oecomys, widely distributed across South America, Oecomys catherinae (Thomas, 1909) [9] stands out due to its broad geographic range in Brazil, occurring along the eastern region from the state of Santa Catarina to Paraíba [2,10,11]. Oecomys catherinae occupies multiple biomes, including the Amazon, Atlantic Forest, and Cerrado, being one of the most widely distributed taxa in Brazil [12].
Phenotypic variation in small mammals, such as those of the genus Oecomys, can be measured through linear or geometric morphometric analyses of craniodental structures [13,14]. When correlated with environmental gradients (e.g., latitude, habitat heterogeneity), these variations may reflect mechanisms like phenotypic plasticity or local adaptations [15]. Although studies have demonstrated these associations in Amazonian Sigmodontinae rodents [1], the underlying mechanisms (plasticity versus adaptation) remain poorly investigated in widely distributed Oryzomyini such as O. catherinae.
The Atlantic Forest and Cerrado, as biodiversity hotspots [16], provide contrasting environmental gradients for O. catherinae. The Atlantic Forest exhibits remarkable altitudinal (0–2700 m) and latitudinal (3–30° S) heterogeneity, encompassing humid forests and semi-humid formations across diverse soils [17,18]. In contrast, the Cerrado combines seasonal savannas (cerrado sensu stricto), gallery forests, veredas, and rocky outcrop vegetation (campos rupestres) on acidic, oligotrophic soils under a seasonal tropical climate [19,20].
We hypothesize that craniodental morphology in O. catherinae varies significantly along latitudinal variation and between biomes, reflecting both local adaptations and phenotypic plasticity to environmental variation. To test this, we conducted morphometric analyses on specimens from the Atlantic Forest and Cerrado biomes, correlating morphological variation with environmental parameters (latitude and biome type) using multivariate statistical approaches. Understanding these patterns advances knowledge of O. catherinae biology and the mechanisms of adaptation and phenotypic plasticity in neotropical small mammals. Furthermore, this study reinforces the importance of integrating morphometric and ecological approaches to elucidate how environmental factors may shape phenotypic diversity across geographic scales and ecological contexts.
2. Materials and Methods
2.1. Sampling and Specimens Localities
Morphometric studies were conducted using small rodent specimens from the scientific collections of the Museu Nacional, Universidade Federal do Rio de Janeiro (MN/UFRJ), and the Coleção Integrada de Mamíferos Reservatórios Silvestres, Instituto Oswaldo Cruz, Fundação Oswaldo Cruz (COLMASTO/IOC/FIOCRUZ). Forty-five specimens of O. catherinae were selected based on their documented geographic origin, availability of complete biometric data, and proper craniodental preservation (Table S1: Geographic location, sex, and age classification of O. catherinae specimens).
Our sampling encompassed 18 localities, distributed across two biomes (Figure 1), The first is the Atlantic Forest biome, including the states of Bahia (Ilhéus), Rio de Janeiro (Cachoeiras de Macacu, Casimiro de Abreu, Guapimirim, Sumidouro), and São Paulo (Ubatuba), presenting habitats ranging from coastal lowland forests to montane formations. The second is the Cerrado biome, including the state of Tocantins (Dianópolis), Goiás (Anápolis, Aporé, Caiapônia, Caldas Novas, Luziânia, Planaltina, Niquelândia, Serranópolis), Distrito Federal (Brasília), Minas Gerais (Pirapetinga), and Mato Grosso do Sul (Dois Irmãos do Buriti), covering vegetation types from open grasslands to forested savannas.
2.2. Age Classification and Morphometric Measurements
Specimens were classified into five age categories based on eruption stage and wear of the third molar (M3), following [21] (Figure 2, Table 1); only specimens from Classes 2 to 5 were included to avoid ontogenetic bias in morphometric analyses, excluding juveniles (Class 1).
For the identification of O. catherinae, the key anatomical structures were measured following protocols derived from previous rodent studies [21,22,23] with methodological adaptations specific to Oecomys morphology [2]. Measurements were taken in dorsal, ventral, and lateral views using a ZAAS digital caliper (150 mm), recording 21 craniodental parameters (Figure 3, Figure 4, Figure 5, Figure 6 and Figure 7, Table 2, Table S2: Mean values and standard deviation (SD) of morphometric variables measured from specimens of O. catherinae collected in different locations of the Brazilian Atlantic Forest and Cerrado). All measurements were taken three times by the same observer, and the mean value was used in analyses.
2.3. Data Analysis
Sexual dimorphism was analyzed to verify morphometric differences between male and female specimens. This was performed using PERMANOVA (Permutational Multivariate Analysis of Variance), based on a Bray–Curtis distance matrix [24], calculated from the measured morphometric variables. This determined whether females and males could be pooled for more robust sampling of phenotypic variation analyses.
Interbiome variation between Cerrado (N = 19) and Atlantic Forest (N = 26) populations was examined using discriminant analysis of principal components (DAPC) [25]. DAPC was selected for its ability to maximize between-group variation while minimizing within-group variation [25]. The optimal number of principal components was determined via cross-validation [26]. The statistical significance of the difference between biomes was also tested with PERMANOVA on the morphometric distance matrix. Furthermore, the effect of latitude on the morphometric variation in the species was investigated using distance-based redundancy analysis (db-RDA), with significance assessed by permutation testing [27]. This analysis aimed to investigate whether there is an influence of a given latitudinal gradient on the morphometry of O. catherinae.
In PERMANOVA and db-RDA, the F-statistic is the ratio of explained to residual mean squares. Its first degree of freedom represents the number of parameters tested, while the second corresponds to the residual degrees of freedom (calculated as the number of observations minus the number of estimated parameters). R^2^ is the proportion of variance explained by the model, and the p-value is the probability of obtaining such an F under the null hypothesis [28].
Analyses were conducted in R v4.4.1 [29]. DAPC used the ‘adegenet’ package v2.1.10 [30]. PERMANOVA, distance matrices, permutation tests, and db-RDA employed the ‘vegan’ package v2.6-10 [31]. A 5% significance threshold was applied [32].
3. Results
3.1. Sexual Dimorphism
The PERMANOVA results revealed no statistically significant sexual dimorphism (R^2^ = 0.06; F_1,46_ = 2.73; p = 0.08), indicating that male and female O. catherinae do not exhibit significant differences in morphometric traits. Consequently, all specimens (males and females) were included in subsequent analyses.
3.2. Comparison Between Biome Populations
DAPC comparing O. catherinae populations from the Cerrado and Atlantic Forest biomes retained six principal components, which explained approximately 96% of the total variation. The analysis correctly classified 72% of specimens into their respective biome groups. Significant morphometric differences were detected between the two biomes (R^2^ = 0.18; F_1,46_ = 10.36; p < 0.01; Figure 8). The analysis identified BPL as the most influential morphometric variable in distinguishing the groups (Figure 9).
3.3. Latitude Effect
A statistically significant, although weak, influence of latitude on the morphometric variation in O. catherinae was recorded (F_1,46_ = 3.63; adjusted R^2^ = 0.05; p = 0.03). This indicates that the morphometry of the specimens varies with latitude, with a general tendency to increase at higher latitudes (Figure 10). Specifically, specimens from southern localities (farther from the Equator) tended to exhibit larger morphometric measurements (e.g., cranial or dental dimensions), while those from northern localities (closer to the Equator) showed smaller values. However, this latitudinal pattern was non-linear, as evidenced by the scatterplot, indicating that the rate of morphometric change does not follow a consistent gradient across latitudes.
3.4. Oecomys catherinae Craniodental Morphometric Comparison Between Atlantic Forest and Cerrado Biomes
In general, skulls of O. catherinae from the Atlantic Forest tend to be wider and longer, while those from the Cerrado show a somewhat more reduced and variable morphology. In this regard, Atlantic Forest specimens presented slightly larger skulls, with higher mean values for characteristics such as bony palate length (BPL: 6.89 ± 0.46 mm vs. 6.03 ± 0.92 mm), condylo-incisive length (CIL: 28.83 ± 1.41 mm vs. 28.06 ± 2.34 mm), and length of nasals (LN: 11.54 ± 0.91 mm vs. 9.64 ± 2.16 mm). Additionally, the lower length of diastema (LLD) was also greater in the Atlantic Forest (3.03 ± 0.31 mm) compared to in the Cerrado (2.71 ± 0.53 mm). On the other hand, length of rostrum (LR) showed slightly higher mean values in Cerrado specimens (9.28 ± 1.60 mm) than in Atlantic Forest specimens (9.16 ± 0.74 mm) (Table S2: Mean values and standard deviation (SD) of morphometric variables measured in O. catherinae specimens from Atlantic Forest and Cerrado locations).
4. Discussion
The absence of significant sexual dimorphism in O. catherinae corroborates previously documented patterns for the genus [33,34] and may reflect selective pressures for locomotor efficiency in arboreal habitats, which stabilize traits associated with this mode of movement [35]. This pattern contrasts with terrestrial rodents, where intense male competition for access to females leads to pronounced sexual dimorphism [36]. Despite this intersexual conservatism, O. catherinae exhibits significant morphological variation among populations from different biomes. Our DAPC analysis revealed morphometric divergence between Atlantic Forest and Cerrado individuals, with detectable latitude effects suggesting phenotypic plasticity along ecological gradients. This pattern supports our hypothesis and mirrors findings in other neotropical rodents: Holochilus brasiliensis showed biome-associated morphometric differences linked to vegetation structure [37], while four species (Akodon cursor, Cerradomys subflavus, Oligoryzomys nigripes, and Oxymycterus dasythrichus) demonstrated cranial morphology shifts along temperature and humidity gradients [38]. These cases reinforce the fundamental role of environmental factors in shaping morphological changes in small mammals.
Although latitude showed only modest effects on morphometric variation, this pattern aligns with both Bergmann’s rule (which predicts larger body size at higher latitudes) and latitudinal diversity patterns [39]. These trends may reflect adaptations to climatic pressures or resource availability, with environmental humidity—through its effect on heat exchange—representing one potentially significant factor influencing mammalian body size variation [40]. The macroecological trends identified here, while consistent with theoretical predictions [41,42], require future studies combining morphological time series and in situ ecological data. Such a multidimensional approach would prove particularly valuable for assessing latitudinal variation, where morphological patterns may emerge from contemporary ecological pressures, evolutionary legacies, or their interactions—not only in O. catherinae, but as a model for neotropical small mammal diversification.
Smaller skulls in the Cerrado compared to in the Atlantic Forest may reflect thermal adaptation, aligning with the trend of size reduction in warmer climates [43]. The longer snout in Cerrado individuals could be an adaptation to drier climates, facilitating thermoregulation and reducing water loss, similarly to patterns observed in other rodents where humidity and temperature influence rostral development [44]. The greater morphological variability in the Cerrado may reflect an adaptive response to environmental heterogeneity and seasonal resource fluctuations, while the uniformity in the Atlantic Forest may be associated with more stable ecological conditions, since body size is shaped by multiple ecological pressures (e.g., resource availability vs. competition) that act in concert—studying them in isolation may lead to misinterpretations [45].
Skull shape differences may also relate to diet: in the Cerrado, a longer rostrum may improve foraging efficiency in an environment with more dispersed resources and harder seeds, while in the Atlantic Forest, a more predictable diet may favor more uniform skulls [46,47]. These morphological differences in O. catherinae between the Atlantic Forest and Cerrado follow patterns observed in other rodents, where climate, resource availability, and competition shape cranial morphology [38]. The smaller size and greater variability in the Cerrado may represent adaptive responses to a hotter, drier, and more heterogeneous environment, while the Atlantic Forest’s uniformity reflects more stable ecological conditions.
Our spatially representative sampling across Atlantic Forest and Cerrado biomes, combined with detailed morphological analyses, provides a robust foundation for macroevolutionary inferences (i.e., large-scale evolutionary patterns across time/space) [48,49]. However, as this study focused solely on cranial variation, locomotor effects remain speculative. Larger samples would enhance resolution for detecting subtle interpopulation variation, while integrating ecological data (e.g., microclimate measurements) with molecular data (e.g., selection signatures in candidate genes) could clarify whether observed variations reflect phenotypic plasticity, local adaptation, gene–environment interactions, or cryptic speciation.
To advance this research, three complementary approaches appear particularly promising: (1) application of biomechanical models [50] to test form–function relationships across different contexts; (2) genomic integration [51] to discriminate plastic vs. genetic components of variation; and (3) temporal analyses of museum specimens [52] to assess historical responses to environmental change. Additionally, future studies could investigate postcranial skeletal variation (e.g., limb proportions, pelvic morphology) to test hypotheses about locomotor efficiency and its role in biome-specific adaptations.
5. Conclusions
Our study reveals that O. catherinae exhibits no significant sexual dimorphism, consistent with stabilizing selection for arboreal locomotion, while significant cranial divergence exists between Atlantic Forest and Cerrado populations. The biome-specific morphological differences likely reflect adaptations to warmer, drier climates and a more heterogeneous resource distribution in the Cerrado compared to in the stable Atlantic Forest environment. The influence of latitude on cranial morphometry was modest but aligned with ecogeographic trends, such as Bergmann’s rule.
To fully disentangle the roles of phenotypic plasticity and genetic adaptation, future studies should integrate genomic analyses with biomechanical modeling. Investigating postcranial morphology could further clarify how locomotor specialization interacts with environmental pressures. These findings underscore the importance of biome-driven selection in shaping morphological diversity in neotropical rodents, offering a framework for understanding adaptive responses to habitat variation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Patton J.L. da Silva M.N.F. Malcolm J.R. Mammals of the Rio Juruá and the evolutionary and ecological diversification of Amazonia Bull. Am. Mus. Nat. Hist.2000244130610.1206/0003-0090(2000)244<0001:MOTRJA>2.0.CO;2 · doi ↗
- 2Carleton M.D. Musser G.G. Order Rodentia Mammals of South America: Volume 2—Rodents Patton J.L. Pardiñas U.F.J. D’Elía G. University of Chicago Press Chicago, IL, USA 2015393417
- 3Hershkovitz P. Mice, land bridges, and Latin American faunal interchange Ectoparasites of Panama Wenzel R.L. Tipton V.J. Field Museum of Natural History Chicago, IL, USA 1960725751
- 4Pardiñas U.F.J. Teta P. Salazar-Bravo J. A new species of arboreal rat, Genus Oecomys (Rodentia: Cricetidae) from Chaco J. Mammal.2016971177119610.1093/jmammal/gyw 070 · doi ↗
- 5Saldanha J. Rossi R.V. Integrative Analysis Supports a New Species of the Oecomys catherinae Complex (Rodentia: Cricetidae) from Amazonia J. Mammal.2021102698910.1093/jmammal/gyaa 145 · doi ↗
- 6Weksler M. Phylogenetic Relationships of Oryzomyine Rodents (Muroidea: Sigmodontinae): Separate and combined analyses of morphological and molecular data Bull. Am. Mus. Nat. Hist.2006296114910.1206/0003-0090(2006)296[0001:PROORM]2.0.CO;2 · doi ↗
- 7Rocha R.G. Fonseca C. Zhou Z. Leite Y.L.R. Costa L.P. Taxonomic and conservation status of the elusive Oecomys cleberi (Rodentia: Sigmodontinae) from Central Brazil Mamm. Biol.20127741441910.1016/j.mambio.2012.02.004 · doi ↗
- 8Suárez-Villota E.Y. Carmignotto A.P. Brandão M.V. Percequillo A.R. Silva M.J.J. Systematics of the Genus Oecomys (Sigmodontinae: Oryzomyini): Molecular phylogenetic, cytogenetic and morphological approaches reveal cryptic species Zool. J. Linn. Soc.201818418221010.1093/zoolinnean/zlx 095 · doi ↗