Intestinal Alkaline Phosphatase Expression in Response to Escherichia coli Infection in Nursery Pigs
Sireethon Maksin, Attapon Kamlangdee, Alongkot Boonsoongnern, Prapassorn Boonsoongnern

TL;DR
This study shows that infection with E. coli in young pigs increases intestinal alkaline phosphatase (IAP), which may help defend against gut infections.
Contribution
The study demonstrates that IAP is upregulated in response to E. coli infection in pigs, suggesting a potential role as a gut health biomarker.
Findings
IAP levels increased significantly in the small intestine of pigs infected with E. coli K88.
IAP expression was upregulated on the apical surface of intestinal villi following bacterial challenge.
The increase in IAP is likely triggered by lipopolysaccharides from Gram-negative bacteria.
Abstract
Intestinal alkaline phosphatase (IAP) is an enzyme produced by cells lining the small intestine that plays a key role in maintaining gut health. In this study, we investigated how IAP responds in young pigs infected with Escherichia coli (E. coli) K88, a bacterium known to cause diarrhea. After exposure to E. coli, we found that IAP levels were significantly increased in the small intestine, particularly on the surface of the intestinal villi. This increase was observed using laboratory techniques such as immunohistochemistry and molecular analysis. The results suggest that IAP is activated in response to bacterial infection and may serve as a natural defense mechanism against harmful substances such as lipopolysaccharides (LPSs) from bacteria. Our findings highlight the potential use of IAP as a marker for gut health and a tool for detecting intestinal infections in animals.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
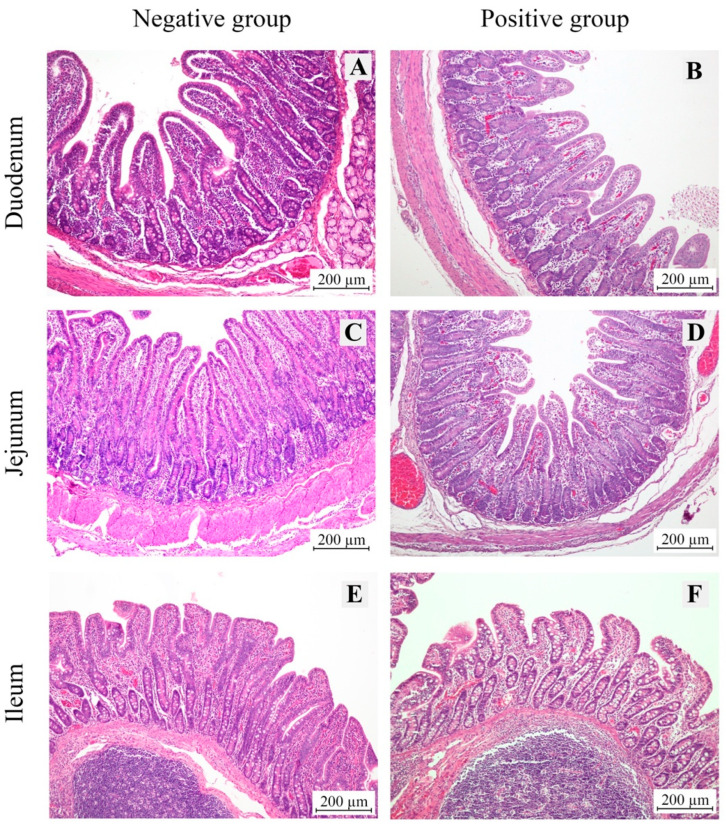 Figure 1
Figure 1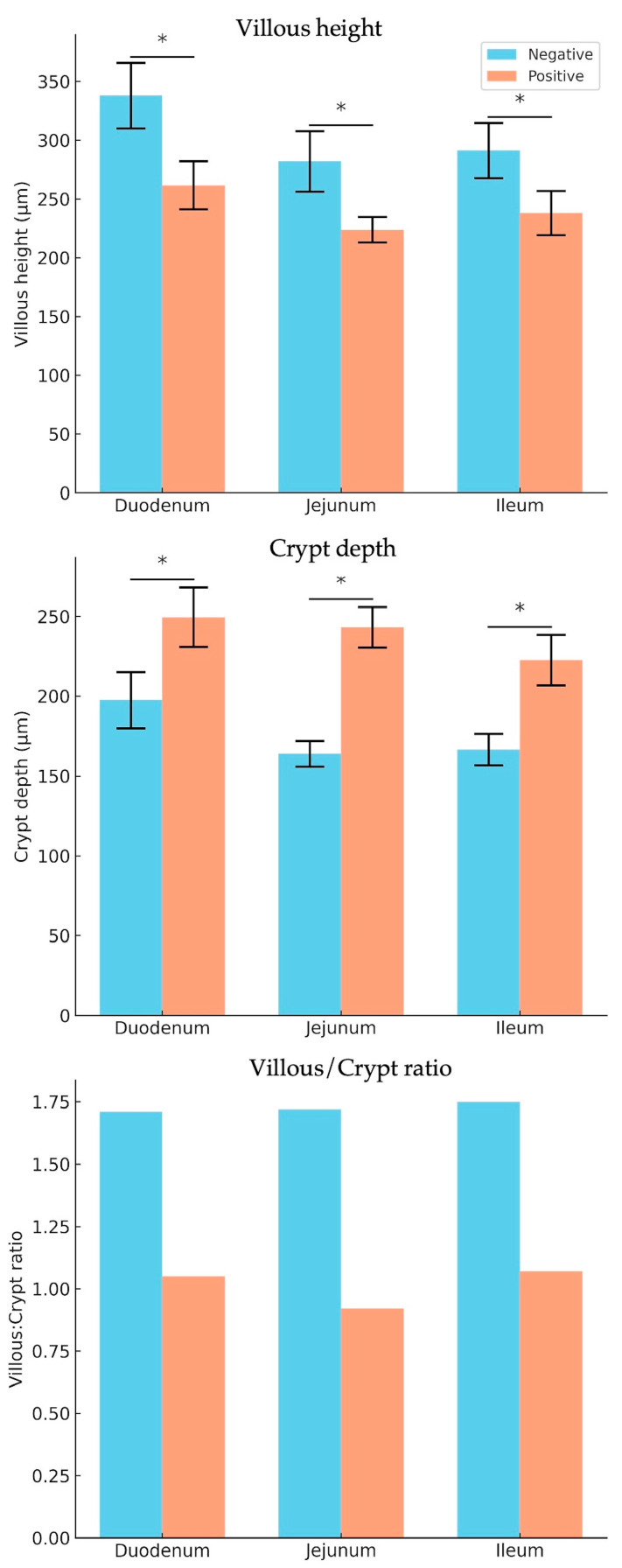 Figure 2
Figure 2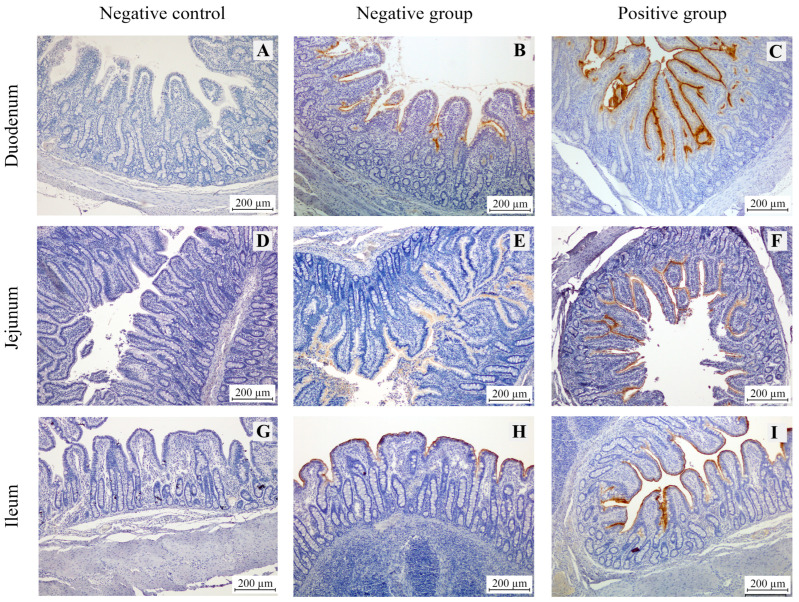 Figure 3
Figure 3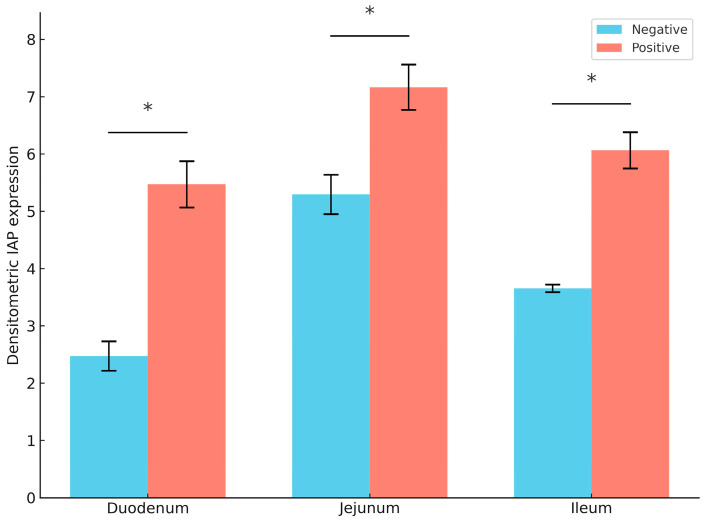 Figure 4
Figure 4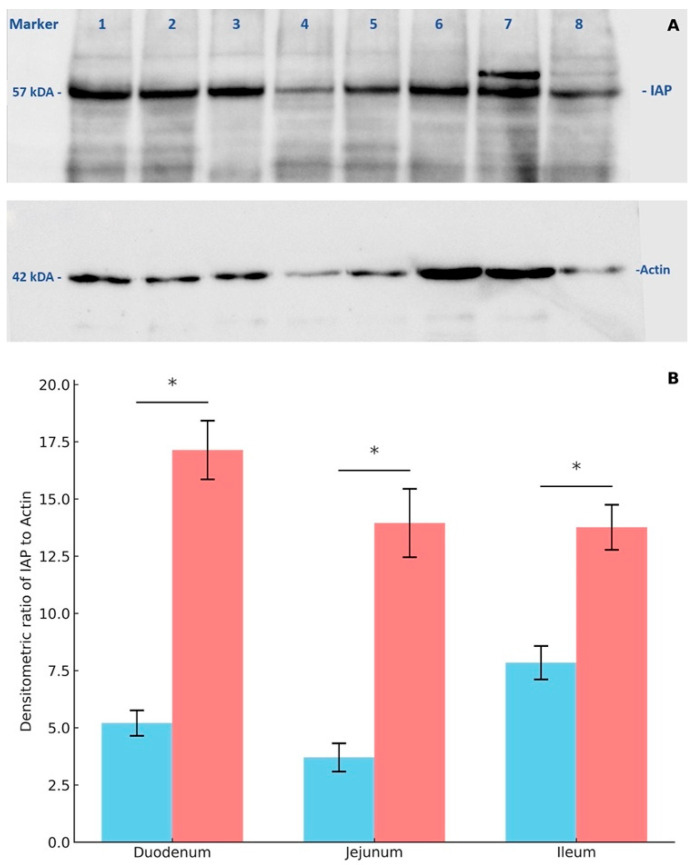 Figure 5
Figure 5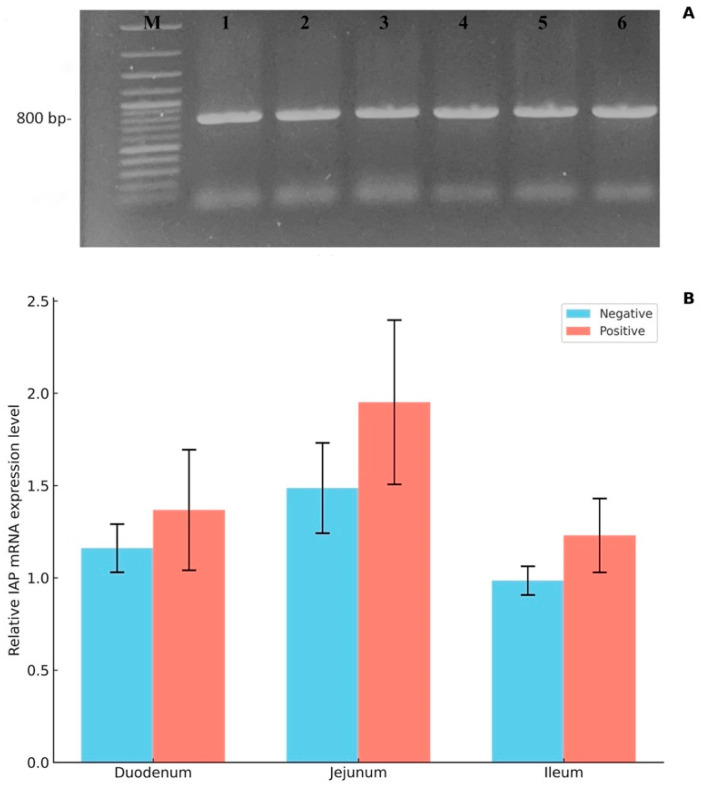 Figure 6
Figure 6- —Faculty of Veterinary Medicine, Kasetsart University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsViral gastroenteritis research and epidemiology · Alkaline Phosphatase Research Studies · Clostridium difficile and Clostridium perfringens research
1. Introduction
Piglet weaning is a critical stage of postnatal growth in a commercial pig production system and is often associated with gastrointestinal infections [1]. In particular, early weaning can have negative consequences, including increased susceptibility to stress (weaning stress), which can affect the growth performance and intestinal health of piglets [2,3]. Weaning stress can lead to diarrhea in piglets, increasing their susceptibility to pathogenic bacterial infections [4]. Post-weaning diarrhea (PWD) in piglets is a multifactorial disease associated with enterotoxigenic Escherichia coli (ETEC) strains [5]. It typically appears during the first two weeks after weaning. ETEC strains expressing F4 and F18 fimbriae are the primary pathogens responsible for PWD in piglets [6]. They impair gut health by adhering to intestinal enterocytes via fimbriae. The presence of fimbrial adhesins mediates attachment to porcine enterocytes, whereas enterotoxins disrupt fluid homeostasis in the small intestine, leading to watery diarrhea [7]. Moreover, ETEC infection promotes inflammation, as indicated by the expression of pro-inflammatory cytokines (IL-1β, IL-6, IL-17, IL-18, and TNF-α) in the jejunum [7]. PWD contributes to significant economic losses globally in swine production due to increased morbidity and mortality and decreased growth rates [5,8,9].
Intestinal alkaline phosphatase (IAP) is a small intestinal brush border enzyme secreted by enterocytes, playing a vital role as a gut mucosal defense factor [10,11]. IAP is expressed and secreted by intestinal epithelial cells, remaining active within both the mucosal membrane and the intestinal lumen [12]. IAP is essential for inactivating lipopolysaccharide (LPS) through dephosphorylation of its lipid A moiety [13]. In wild-type (WT) mice under normal conditions, LPS that crosses the intestinal brush border is typically in a dephosphorylated (inactive) form due to exposure to IAP. In contrast, IAP-knockout (KO) mice lack the ability to detoxify LPS in this manner [14]. In addition to investigating the effect of IAP in reducing the toxification of LPS, rats were orally exposed to LPS and L-phenylalanine, an IAP inhibitor, administered simultaneously with LPS. As a result, serum LPS levels increased twofold [15]. It has been reported that eight-week-old pigs were given calf intestinal alkaline phosphatase before LPS administration to neutralize and detoxify LPS [16].
Due to the properties of IAP in detoxifying LPS, this study aimed to reveal the relationship between IAP expression in the small intestine of pigs challenged with E. coli K88 compared to normal pigs, as this has become a key diagnostic or therapeutic marker for gut health in pigs.
2. Materials and Methods
2.1. Animals and Experimental Design
Sixteen growing pigs (5 weeks old) were obtained from Charoen Pokphand Foods Public Company Limited (CPF, Thailand) and housed at the farm animal demonstration and research facility of the Faculty of Veterinary Medicine, Kasetsart University. The animals were randomly assigned to two groups (n = 8 per group): a negative (control) group and a positive (treatment) group. The positive group was orally challenged with Escherichia coli (E. coli) strain K88 at a concentration of 2 × 10^8^ CFU/mL, administered at a dosage of 2 mL per pig at 0 and 24 h. The pigs were monitored daily, and fecal consistency was scored using a five-point scale based on the physical characteristics of the feces: 1 = firm and shaped, 2 = moist and shaped, 3 = loose, 4 = homogeneous watery, and 5 = watery with separated components. Scores ≥ 3 were operationally defined as indicative of diarrhea in this study.
At the end of the experiment (5 days post-challenge), all pigs were humanely euthanized via intravenous injection of sodium thiopental (100 mg/kg body weight). Intestinal tissue samples were collected from three segments—the duodenum, jejunum, and ileum—for further histological and molecular analyses. All animal procedures were reviewed and approved by the Animal Care and Use Committee of the Faculty of Veterinary Medicine, Kasetsart University, Thailand (ACKU64-VET-064).
2.2. Morphometric Analysis of the Small Intestinal Mucosa
Segments of the small intestine were fixed in 10% neutral-buffered formalin and embedded in paraffin. The samples were sectioned at a thickness of 5 µm, deparaffinized with xylene, and rehydrated through a graded ethanol series (100%, 95%, and 75%). The sections were subsequently stained with hematoxylin and eosin (H&E), followed by dehydration in 70%, 95%, and 100% ethanol, and cleared with xylene. All slides were examined and imaged using a light microscope equipped with a digital camera system (Olympus, Tokyo, Japan).
For morphometric analysis, transverse H&E-stained sections of the small intestinal mucosa were evaluated under the microscope. Only images captured at 100x magnification were used for measurements of villous height (VH) and crypt depth (CD). Ten villi and crypts were randomly selected per section, and the mean VH and CD values were calculated for each slide. These values were used for statistical comparison between the groups and for determining villous height-to-crypt depth (VH/CD) ratios.
2.3. Immunohistochemistry
Paraffin-embedded tissue sections were deparaffinized in xylene and rehydrated through a graded ethanol series. Antigen retrieval was performed by immersing the slides in Tris–EDTA buffer (pH 9.0) at 95 °C for 30 min, followed by three washes with 0.1% Tween 20 in Tris-buffered saline (TBST). To block nonspecific protein binding, the slides were incubated with 5% normal goat serum in TBST at room temperature (25 °C) for 30 min. The sections were then incubated overnight at 4 °C with a 1:200 (v/v) dilution of anti-intestinal alkaline phosphatase antibody (ab186422, Abcam, UK). Endogenous peroxidase activity was quenched using 3% hydrogen peroxide in TBS. After three washes with 0.1% TBST, the slides were incubated with a 1:1000 (v/v) dilution of horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG (KPL, Gaithersburg, MD, USA) for 30 min at room temperature (25 °C). Immunoreactivity was visualized using a DAB Detection Kit (Vector Laboratories, Newark, CA, USA), and the nuclei were counterstained with hematoxylin. Negative controls were performed by omitting the primary antibody, and healthy pig tissues were used as positive controls to confirm antibody specificity. The slides were dehydrated in 70%, 95%, and 100% ethanol, cleared in xylene, and mounted. All samples were observed and imaged using a light microscope equipped with a digital camera system (Olympus, Tokyo, Japan). The intensity of IAP expression was analyzed using Fiji (ImageJ version 1.53t; NIH, Bethesda, MD, USA).
2.4. Western Blotting
Intestinal tissues were lysed in RIPA buffer (50 mM Tris-HCl, pH 7.4; 150 mM NaCl; 0.25% sodium deoxycholate; 1% Triton X-100; and 1 mM EDTA), supplemented with 1% protease inhibitor cocktail (Sigma-Aldrich, St. Louis, MO, USA). The lysates were centrifuged at 15,000× g for 20 min at 4 °C, and the supernatant was collected. Total protein concentrations were determined using the Bradford assay (Bio-Rad, Hercules, CA, USA) with bovine serum albumin as the standard. Equal amounts of protein (10 µg per sample) were mixed with 2× loading buffer and denatured at 95 °C for 5 min.
Proteins were separated using 10% SDS–PAGE and transferred to polyvinylidene fluoride (PVDF) membranes (Bio-Rad, Hercules, CA, USA) under electrophoresis conditions (200 V or 20 mA). The membranes were blocked in TBS (50 mM Tris-HCl, pH 8.0; 150 mM NaCl; 0.1%) containing 5% bovine serum albumin (BSA) for 1 h at room temperature (25 °C). The blots were incubated overnight at 4 °C with a 1:2000 dilution of anti-intestinal alkaline phosphatase (IAP) antibody (Abcam, Cambridge, UK). The following day, the membranes were washed five times (5 min each) in TBST (TBS with 0.1% Tween-20) and incubated for 1 h at 25 °C with horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG secondary antibody (Sigma, USA) at a 1:10,000 dilution. After additional washing in TBST and a final rinse in TBS, the signal was developed using Clarity™ Western ECL Substrate (Bio-Rad, Hercules, CA, USA) and visualized using a ChemiDoc Imaging System (Bio-Rad, USA). Band intensities were quantified using Image Lab software (Bio-Rad, Hercules, CA, USA) and were expressed as relative optical density.
To detect the internal control protein, the membranes were stripped with stripping buffer (10% SDS, 0.5 M Tris-HCl, pH 6.8, and 2-mercaptoethanol) at 50 °C for 45 min. The membranes were then washed with TBST, blocked again with 5% BSA in TBS, and reprobed with anti-β-actin antibody (Abcam, Cambridge, UK) at a 1:5000 dilution. HRP-conjugated goat anti-mouse IgG (Sigma, St. Louis, MO, USA) was used as the secondary antibody. The remaining steps were performed as described for IAP detection.
2.5. Real-Time Polymerase Chain Reaction (qPCR)
Total RNA was extracted from the intestinal samples (duodenum, jejunum, and ileum) using an RNA extraction kit (Geneaid^®^, New Taipei City, Taiwan), according to the manufacturer’s instructions. The concentration and purity of the extracted RNA were determined by measuring the absorbance ratio at 260/280 using a NanoDrop ND1000 spectrophotometer (Thermo Scientific, Wilmington, DE, USA). Complementary DNA (cDNA) was synthesized from total RNA using the Omniscript RT Kit (Qiagen, Hilden, Germany), following the manufacturer’s protocol.
To validate the target gene, selected RNA samples were reverse transcribed into cDNA, followed by gel electrophoresis and gel purification. The purified product was subjected to Sanger sequencing to confirm the identity of the intestinal alkaline phosphatase (IAP) gene. The validated sequence was used to design specific primers for quantitative PCR. The primer sequences (5′–3′) were as follows:
IAP forward: CTAAAGGGGCAGATGAATGG
IAP reverse: CACCTGTCTGTCCACGTTGT
RPL4 forward: CAAGAGTAACTACAACCTTC
RPL4 reverse: GAACTCTACGATGAATCTTC
The qPCR was performed using the following thermal cycling conditions: initial denaturation at 95 °C for 2 min, followed by 40 cycles of denaturation at 95 °C for 30 s and annealing/extension at 72 °C for 1 min.
2.6. Statistical Analysis
Statistical analyses were performed using R software version 4.0.2 (R Foundation for Statistical Computing, Vienna, Austria). Student’s t-test was used to compare villous height (VH), crypt depth (CD), the villous height-to-crypt depth (VH/CD) ratio, fecal scores, intestinal alkaline phosphatase (IAP) protein expression (assessed by Western blot and DAB staining), and IAP mRNA expression levels between normal piglets and those challenged with Escherichia coli K88. A p-value of <0.05 was considered statistically significant.
Western blot data were quantified by measuring the area under the curve of immunoreactive bands using Image Lab software version 6.0.1 (Bio-Rad, USA). DAB staining intensity was analyzed using ImageJ software (NIH, Bethesda, MD, USA). IAP mRNA expression levels were determined using quantitative PCR and were normalized to the reference gene RPL4.
3. Results
3.1. Assessment of Diarrheal Response Induced by E. coli Infection
Fecal consistency was evaluated daily from Day 0 (D0) to Day 5 (D5) post-challenge in both the negative and positive piglet groups. On D0, all piglets exhibited normal fecal scores (score = 1), indicating no pre-existing symptoms. However, from D2 onwards, the positive group showed a marked increase in fecal scores, with values reaching 3–4 by D4–D5, indicative of moderate to severe diarrhea. In contrast, the negative group maintained consistently low scores throughout the study. A detailed summary of individual fecal scores by treatment group and post-challenge day is provided in Table 1. On Day 0, both groups showed uniformly low scores (1.00 ± 0.00). From Day 1 onward, the E. coli-infected group displayed significantly higher scores than the control group (p < 0.0001). The mean fecal scores in the infected group were above 3.0 from Day 2 to Day 5, with a peak of 3.75 ± 0.16 on Days 3 and 4, clearly indicating diarrhea. In contrast, the control group maintained consistently low scores throughout the trial. These results confirm the successful induction of diarrhea following E. coli challenge and support the validity of the infection model.
3.2. Mucosal Morphometry
In pigs infected with E. coli, the villi appeared shortened, indicating atrophy due to epithelial damage, and exhibited increased degradation at the tips, accompanied by deeper crypts extending into the lamina propria (Figure 1). Morphometric analysis of the villi in the three sections of the small intestine (duodenum, jejunum, and ileum) revealed a significant reduction in villous height in E. coli-infected piglets across all three sections compared to the uninfected control group. This reduction in villous height (VH) was statistically significant, as shown in Table 2 and Figure 2. Additionally, this finding was consistent with a significant increase in the depth of the crypts of Lieberkühn in the E. coli-infected group. Consequently, the villous height-to-crypt depth ratio (VH/CD ratio) was higher in the control group than in E. coli-infected piglets.
3.3. Immunohistochemical Analysis of Intestinal Alkaline Phosphatase Expression
The immunohistochemical staining results for intestinal alkaline phosphatase (IAP) in the duodenum, jejunum, and ileum of piglets revealed that IAP expression was localized at the tips of the villi in all sections, as indicated by brown staining (Figure 3). Image analysis using ImageJ software showed that the E. coli-infected group exhibited significantly higher IAP density in all sections of the small intestine compared to the non-infected group (Figure 4). Among the analyzed intestinal sections, the jejunum exhibited the highest IAP density in both E. coli-infected and control piglets (Figure 4).
3.4. Western Blot Analysis of Intestinal Alkaline Phosphatase Expression
The expression of intestinal alkaline phosphatase (IAP) in small intestinal tissues was analyzed through protein extraction and Western blotting, revealing an IAP molecular weight of 57 kDa and a β-actin molecular weight of 42 kDa (Figure 5A). Image analysis using Chemidoc software (version 6.1; Bio-Rad, Hercules, CA, USA) was performed to determine the relative density of the IAP-to-β-actin ratio. The results demonstrate that IAP expression was significantly higher in all sections of the small intestine in E. coli-infected piglets compared to the non-infected group (Figure 5B). Notably, IAP expression was significantly elevated in the jejunum and ileum of the E. coli-infected group compared to the non-infected group.
3.5. Quantitative Real-Time RT-PCR
The mRNA expression of intestinal alkaline phosphatase (IAP) in the small intestine (mid-part of the jejunum) of E. coli-infected piglets was analyzed using qPCR with Sybr green and was compared to the non-infected group. Bands corresponding to IAP mRNA expression were detected in all samples, with a band density observed at 800 bp (Figure 6A). The relative expression of IAP was conducted and normalized to RPL4 mRNA expression as an internal control. The results reveal that all segments of the small intestine exhibited higher IAP mRNA expression in E. coli-infected piglets compared to the corresponding sections in the non-infected group; however, this difference was not statistically significant (Figure 6B).
4. Discussion
In this study, we analyzed the expression of intestinal alkaline phosphatase (IAP) in pigs following an enterotoxigenic Escherichia coli (ETEC) K88 challenge and compared this to healthy controls. Post-weaning piglets are highly susceptible to E. coli infections, which often result in post-weaning diarrhea (PWD). A survey by Luppi et al. across multiple farms in various European countries identified ETEC as the primary causative agent of PWD, with F4 and F8 ETEC being the most prevalent strains [8].
IAP expression was predominantly observed on the apical surface of villi, consistent with the localization of zebrafish IAP on the intestinal lumen brush border [17]. Notably, IAP expression levels were highest in the jejunum compared to the duodenum and ileum in both healthy and E. coli-challenged pigs. This regional variation may be attributed to the higher affinity of membrane-bound IAP in the jejunum, underscoring site-specific differences in IAP distribution along the intestine [18]. Moreover, pigs challenged with E. coli exhibited a significant increase in IAP expression across all segments of the small intestine compared to their healthy counterparts, suggesting the potential role of IAP in the intestinal response to E. coli infection. Moreover, pigs challenged with E. coli exhibited a significant increase in IAP expression across all segments of the small intestine compared to their healthy counterparts, suggesting the potential role of IAP in the intestinal response to E. coli infection.
Although IAP protein expression was markedly increased in the intestinal tissues of E. coli-infected pigs, the corresponding mRNA levels did not differ significantly from those in the healthy group. This discrepancy suggests that IAP expression may be regulated at the post-transcriptional or post-translational level. Supporting this notion, Bates et al. [17] demonstrated that IAP activity was induced in zebrafish during gut microbiota colonization without a corresponding increase in mRNA expression, implying transcription-independent activation of the enzyme. Possible mechanisms may include increased protein stability, enzyme phosphorylation, or enhanced catalytic efficiency in response to microbial or inflammatory stimuli [17].
ETEC-induced diarrhea in weaned pigs leads to mucosal inflammation and villous atrophy, as evidenced by a reduction in villous height observed in the duodenum, jejunum, and ileum. These findings align with previous studies [2,4,19]. A decreased villous height-to-crypt depth (VH/CD) ratio was also observed in the small intestine, consistent with previous studies reporting similar findings in various intestinal regions [4,20,21,22]. Given the critical role of mucosal morphometry in the small intestine, particularly the VH/CD ratio, this parameter is fundamental to nutrient absorption [23]. Therefore, it serves as a key indicator in studies investigating intestinal morphology and function.
Lipopolysaccharide (LPS) is a major component of the outer membrane of Gram-negative bacteria, including E. coli, and functions as a potent endotoxin by excessively stimulating innate immune signaling through Toll-like receptor 4 (TLR-4) [24]. Upregulation of TLR-4 expression triggers the activation of NF-κB and MAPK pathways, leading to the production of proinflammatory cytokines [7]. Pigs challenged with ETEC or LPS exhibit a pronounced pro-inflammatory response, characterized by increased expression of IL-1β, IL-6, IL-8, IL-17, IL-18, and TNF-α [25,26], while simultaneously suppressing anti-inflammatory cytokines such as IL-4, IL-10 and TGF-β [20].
Intestinal alkaline phosphatase (IAP) plays a crucial role in dephosphorylating LPS, a key component of Gram-negative bacteria [11]. By detoxifying LPS, IAP helps mitigate inflammation, as demonstrated in previous studies [11,15,16,27]. Chen et al. reported that IAP-knockout (IAP-KO) mice challenged with LPS exhibited significantly higher IL-6 levels compared to wild-type (WT) mice [14]. Additionally, dietary supplementation with exogenous IAP has been shown to reduce the incidence of diarrhea, thereby improving growth performance [28,29]. Furthermore, exogenous IAP has been found to support gut microbial homeostasis by promoting the growth of intestinal commensal bacteria while simultaneously inhibiting the Enterobacteriaceae population [29,30,31].
Although our study demonstrates increased IAP expression following E. coli infection, we did not evaluate TLR4 expression, proinflammatory cytokines (e.g., IL-6 and TNF-α), or NF-κB activation. Therefore, it remains uncertain whether the elevated IAP levels directly suppress TLR4 signaling in piglets. Nonetheless, based on previous studies in mice demonstrating IAP-mediated inhibition of TLR4 signaling [14,27], it is plausible that IAP may modulate inflammatory responses through a similar pathway in pigs. Further studies are needed to confirm whether this pathway operates similarly in pigs.
5. Conclusions
Gram-negative bacteria, such as Escherichia coli, can induce intestinal alkaline phosphatase (IAP) expression in response to lipopolysaccharide (LPS), which serves as a substrate and may enhance enzymatic activity. This observation is consistent with the findings of Melo et al., who reported IAP upregulation in the jejunum of pigs challenged with LPS [27]. The upregulation of IAP likely represents an adaptive protective mechanism to detoxify LPS and maintain intestinal homeostasis. These findings highlight the critical role of IAP in promoting gut health, modulating immune responses, and supporting the performance of nursery pigs.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Sargeant H.R. Mc Dowall K.J. Miller H.M. Shaw M.A. Dietary zinc oxide affects the expression of genes associated with inflammation: Transcriptome analysis in piglets challenged with ETEC K 88Vet. Immunol. Immunopathol.201013712012910.1016/j.vetimm.2010.05.00120605641 · doi ↗ · pubmed ↗
- 2Mc Lamb B.L. Gibson A.J. Overman E.L. Stahl C. Moeser A.J. Early Weaning Stress in Pigs Impairs Innate Mucosal Immune Responses to Enterotoxigenic E. coli Challenge and Exacerbates Intestinal Injury and Clinical Disease P Lo S ONE 20138 e 5983810.1371/journal.pone.005983823637741 PMC 3634819 · doi ↗ · pubmed ↗
- 3Lu X. Zhang M. Zhao L. Ge K. Wang Z. Jun L. Ren F. Growth performance and post-weaning diarrhea in piglets fed a diet supplemented with probiotic complexes J. Microbiol. Biotechnol.2018281791179910.4014/jmb.1807.0702630199923 · doi ↗ · pubmed ↗
- 4Zou C. Zhao W. Yin S. Xiang X. Tang J. Jia G. Che L. Liu G. Chen X. Tian G. Artificial parasin I protein (API) supplementation improves growth performance and intestinal health in weaned piglets challenged with enterotoxigenic Escherichia coli Anim. Nutr.20241815416510.1016/j.aninu.2024.04.01539263444 PMC 11388718 · doi ↗ · pubmed ↗
- 5Jenkins T.P. Ács N. Arendrup E.W. Swift A. Duzs A. Chatzigiannidou I. Pichler M. KittiläT. Peachey L. Gram L. Protecting the piglet gut microbiota against ETEC-mediated post-weaning diarrhoea using specific binding proteinsnpj Biofilms Microbiomes 2024104210.1038/s 41522-024-00514-838697985 PMC 11066037 · doi ↗ · pubmed ↗
- 6Luise D. Lauridsen C. Bosi P. Trevisi P. Methodology and application of Escherichia coli F 4 and F 18 encoding infection models in post-weaning pigs J. Anim. Sci. Biotechnol.2019105310.1186/s 40104-019-0352-731210932 PMC 6567477 · doi ↗ · pubmed ↗
- 7Ren W. Yin J. Duan J. Liu G. Zhu X. Chen S. Li T. Wang S. Tang Y. Hardwidge P.R. Mouse intestinal innate immune responses altered by enterotoxigenic Escherichia coli (ETEC) infection Microbes Infect.20141695496110.1016/j.micinf.2014.09.00525267358 · doi ↗ · pubmed ↗
- 8Luppi A. Gibellini M. Gin T. Vangroenweghe F. Vandenbroucke V. Bauerfeind R. Bonilauri P. Labarque G. Hidalgo A. Prevalence of virulence factors in enterotoxigenic Escherichia coli isolated from pigs with post-weaning diarrhoea in Europe Porc. Health Manag.201622010.1186/s 40813-016-0039-928405446 PMC 5382528 · doi ↗ · pubmed ↗