Evaluation of Alterations in Nutrient Utilization and Intestinal Health in Response to Heat Stress in Pekin Ducks Based on a Pair-Feeding Experimental Design
Xiangyi Zeng, Arshad Javid, Gregory S. Fraley, Gang Tian, Keying Zhang, Shiping Bai, Xuemei Ding, Jianping Wang, Yan Liu, Yue Xuan, Shanshan Li, Qiufeng Zeng

TL;DR
Heat stress in Pekin ducks reduces nutrient use and gut health, with reduced feed intake being a key factor in these effects.
Contribution
This study characterizes the impact of heat stress on nutrient utilization and gut health in Pekin ducks using a pair-feeding design.
Findings
Heat stress reduces dietary energy, protein, and fat utilization in Pekin ducks.
Heat stress causes gut barrier dysfunction and microbiota changes, worsening intestinal health.
Reduced feed intake is a primary driver of gut health deterioration under heat stress.
Abstract
As global temperatures rise, heat stress (HS) is becoming a prevalent stressor with deleterious consequences for duck growth and health. Under HS conditions, birds reduce feed intake (FI) and undergo physiological adaptations to mitigate heat production or enhance heat dissipation, yet HS effects on nutrient utilization and gut health in meat ducks—particularly under pair-fed experimental designs—remain poorly characterized. This study investigated alterations in nutrient utilization, standardized ileal amino acid digestibility, and intestinal health parameters in Pekin ducks exposed to HS. The results demonstrated that HS induces intestinal barrier dysfunction and microbiota dysbiosis, thereby impairing gut health and consequently reducing dietary nutrient utilization. Critically, although reduced FI constitutes an adaptive mechanism to limit metabolic heat load, data indicate this FI…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
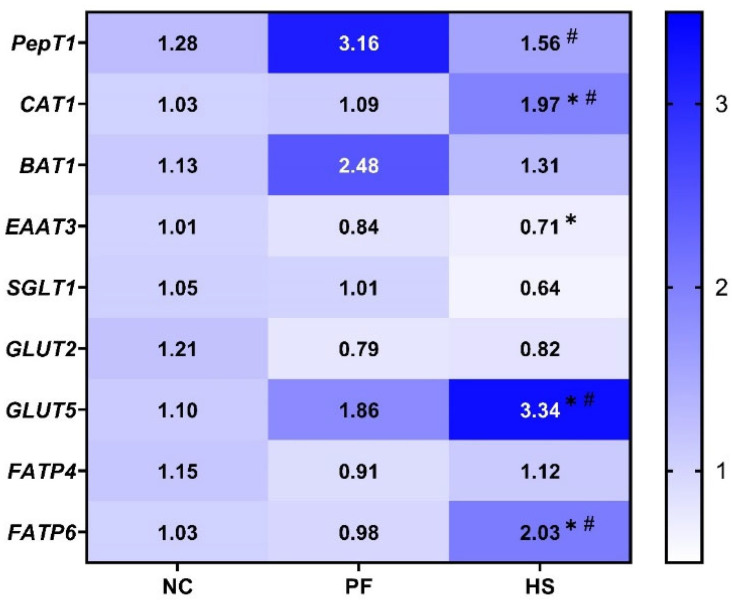 Figure 1
Figure 1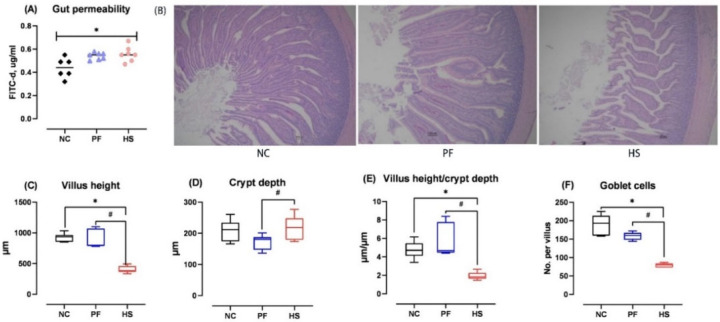 Figure 2
Figure 2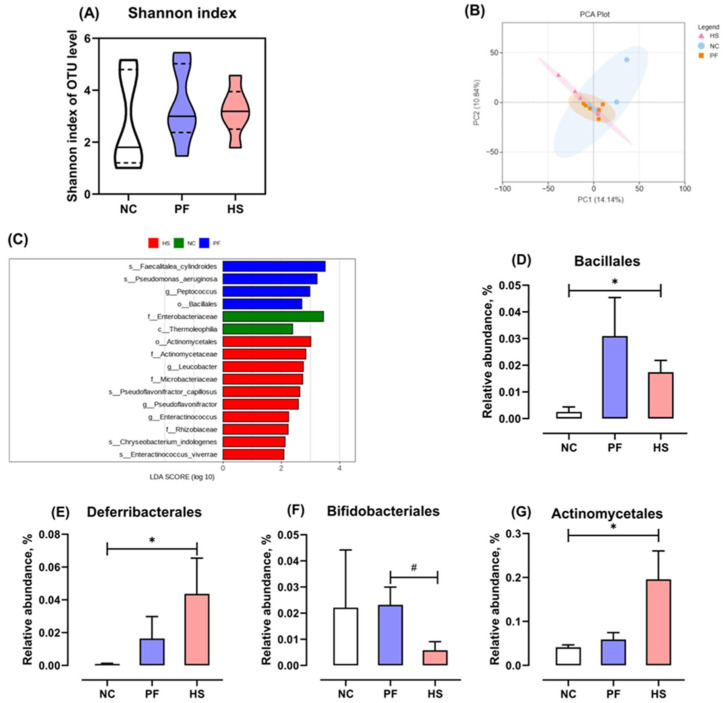 Figure 3
Figure 3- —Sichuan International Science and Technology Innovation Cooperation Program
- —Foreign Experts Affairs Bureau of China
- —National Key R&D Program of China
- —Program for Waterfowl Industry Technology System Innovation Team of Sichuan Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal Nutrition and Physiology · Effects of Environmental Stressors on Livestock · Meat and Animal Product Quality
1. Introduction
As global temperatures rise, heat stress (HS) has become a prevalent stressor with deleterious consequences for duck growth and health. Fast-growing ducks are particularly sensitive to HS due to their high stocking density, high metabolic rate, down feather coverage, and the absence of sweat glands [1]. Under HS conditions, birds typically decrease their feed intake (FI) and undergo a series of physiological alterations that reduce performance to decline heat production [2]. The use of pair-feeding techniques, where birds under thermoneutral conditions are pair-fed the same amount of feed as birds subjected to constant HS, has demonstrated that approximately 81% of the body weight gain (BWG) reduction was attributed to a reduction in FI, with the remaining 19% being the consequence of physiological changes induced by HS per se [3]. From a nutritional perspective, reduced nutrient digestibility and altered postabsorptive metabolism can impair the performance of heat-stressed birds. In our previous study, we reported the decrease in FI and BWG and the metabolic mechanisms underlying the suboptimal performance of Pekin ducks induced by HS [4]. Although FI influences nutrient digestion and absorption in animals [5,6], no studies have investigated the effects of HS on nutrient utilization in meat ducks using a pair-feeding experimental design.
As is well known, intestinal health is critically important for the digestion and absorption of nutrients in poultry [7]. Several studies have demonstrated that HS led to increased crypt depth and reduced villus height and villus height to crypt depth ratio in poultry [8,9,10], resulting in lower nutrient absorptive capacity [11]. The adverse effects of high ambient temperatures are also evident in gut leak, which allow pathogens and toxins present in the intestinal lumen to be translocated, ultimately leading to gut dysfunction [12]. Zhang et al. [13] confirmed that increased gut permeability in broilers under HS conditions was linked to reduced tight junction proteins and altered gut microbiota composition. In particular, gut microbiota could ferment indigestible fiber to produce a multitude of fermentation products, including short-chain fatty acids, that play a vital role in maintaining intestinal barrier function [14]. Nevertheless, the effects of HS on gut microbiota in meat ducks remain largely unexplored. Therefore, in our current study, changes in nutrient utilization and intestinal health in response to HS in Pekin ducks were further interpreted using a metabolic trial, morphological analyses, qRT-PCR and 16S rRNA sequencing. We postulated that HS would have adverse effects on intestinal health, as evidenced by the impairment of jejunal morphology and the alteration of ileal microbiota composition, which reduced intestinal nutrient digestible and absorptive capacity in Pekin ducks. The research results can provide a theoretical basis for mitigating HS-induced intestinal damage in meat ducks through nutritional regulation.
2. Materials and Methods
2.1. Birds, Diets, and Management
This experiment was conducted in the Academic and Research Zone of Ya’an Campus, Sichuan Agricultural University. A total of 240 healthy 28-day-old male Pekin ducks (2054 ± 28 g) were allotted randomly to the following three groups with 8 replicate cages of 10 birds each: the normal control (NC) group at constant 21 ± 1 °C, the HS group at 34 ± 1 °C for 7 h daily with relative humidity (RH) between 50% and 60% and the rest of the day at 25 ± 1 °C, and the pair-fed (PF) group at constant 21 ± 1 °C (in the same house as the ducks in the NC group). The ducks were provided with the same amount of feed as the HS group on the previous day. Temperature and humidity were monitored using hygrothermographs (Tianjin Kehui Instrument Factory, Tianjin, China) mounted above each cage. Each replicate group was housed in a single cage (1.0 × 0.8 × 0.6 m) under a 16 h light/8 h dark cycle. All birds received a starter and a finisher diet, the composition and nutrient density of which are listed in Table 1 and Table 2, respectively, in accordance with our previous research [4]. In our previous study, Pekin ducks subjected to HS conditions (34 ± 1 °C, 7 h/d, 14 d) increased their panting frequency and rectal temperature, as well as the serum concentration of heat shock protein 70 and corticosterone, suggesting that the chronic HS model was successfully established [4]. Therefore, we further evaluated the effect of HS on intestinal health and dietary nutrient utilization and standardized ileal digestibility of amino acids (SIDAA) in this study.
2.2. Intestinal Permeability Determination
On day 42, one duckling per replicate was selected to determine intestinal permeability using fluorescein isothiocyanate dextran (FITC-d, 4kDa, Sigma, Ronkonkoma, NY, USA), an indicator to examine barrier function. All birds were administered FITC-d orally (2.2 mg per duck), and then blood was collected from the jugular vein at two hours post-FITC-d administration. Serum FITC-d levels were quantified at excitation and emission wavelengths of 485 and 528 nm, respectively (BioTek Instruments, Winooski, VT, USA). Subsequently, serum FITC-d concentration was calculated based on a standard curve with known FITC-d levels.
2.3. Jejunal Morphology Examination
On day 42, one duckling per replicate was humanely euthanized through exsanguination for jejunal morphology analysis, gene expression assays, and 16S rRNA sequencing. A fixed jejunal segment was embedded in 10% paraffin, sectioned into 5 μm slices, and stained with either hematoxylin and eosin (H&E) to evaluate intestinal architecture or Alcian blue (AB) to quantify the number of goblet cells. Images were captured using a microscope (BA400 Digital, Xiamen, China) and analyzed using Image-Pro Plus 6.0 (Media Cybernetics, Rockville, MD, USA) to determine villus height (VH), crypt depth (CD), and their ratio (VH/CD). Goblet cells along the villus were counted by light microscopy.
2.4. Gene Expression Assays
Total RNA was extracted from frozen jejunal mucosa samples using a Trizol reagent (TaKaRa, Dalian, China), and first-strand cDNA synthesis was performed with the PrimeScript™ RT Reagent Kit (Takara, China) in accordance with the manufacturer’s instructions. The quantitative real-time PCR (qRT-PCR) was performed on the ABI QuantStudio™ 6 Flex system (Applied Biosystems, Waltham, MA, USA). The primer sequences for the target genes were designed using the National Centre for Biotechnology Information (NCBI) Blast tool. All primers set in the qRT-PCR reaction were run for melting curve analyses to generate a standard curve to assess PCR efficiency. A comprehensive list of all primer sequences used in this study is presented in Table 3. Relative gene expression was quantified by normalizing to the expression of β-actin according to the 2^−ΔΔCt^ method, with the quantity of the NC group scaled to approximately 1.
2.5. 16S rRNA Gene Sequencing
Total genome DNA from the samples was extracted using the cetyltrimethylammonium bromide method, and the concentration and purity of DNA were detected using a 1% agarose gel. Subsequently, PCR amplification and product purification were performed sequentially. Sequencing libraries were generated using the TruSeq^®^ DNA PCR-Free Sample Preparation Kit (Illumina, San Diego, CA, USA) in accordance with the manufacturer’s instructions, and library quality was assessed on the Qubit@ 2.0 Fluorometer (Thermo Scientific, Waltham, MA, USA) and Agilent Bioanalyzer 2100 systems. The samples were then sequenced using the Illumina NovaSeq 6000 platform, and high-quality clean tags were obtained according to the QIIME 2 quality-controlled process. The representative sequences of Operational Taxonomic Units (OTUs) were selected, and OTU clustering was performed based on the principle of 97% sequence similarity.
2.6. Assay of Nutrient Utilization and Standard Ileal Digestibility of Amino Acids
On day 43, two ducks randomly selected from each replicate were housed in metabolic cages (two ducks per cage) and assigned to the original groups. In addition, this experiment included two nitrogen-free diet groups under thermoneutral and HS conditions, with eight replicates per group. All ducks were fed the original diets or the nitrogen-free diet supplemented with 0.5% titanium dioxide (TiO_2_) as an indigestible marker. Excreta samples were collected from each cage over a period of 72 h. Following the removal of any debris, the samples were gathered in each cage and dried in an oven at 65 °C for 3 days. All samples were ground to pass through a 0.5 mm screen and then analyzed for dry matter (DM), gross energy (GE), ether extract (EE), nitrogen (N), calcium (Ca), and total phosphorus (TP) in accordance with the method (AOAC, 2005) [15]. Crude protein (CP) was calculated as N × 6.25. EE in the diets and excreta was measured with a Soxhlet apparatus for approximately 8 h. GE was analyzed by using Parr 6400 oxygen bomb calorimeter (Parr Instrument Co., Moline, IL, USA). The TiO_2_ content in feed and excreta samples was measured according to the method proposed by Short et al. [16]. Nutrient utilization for the experimental diets was calculated with the following formula: Nutrient utilization (%) = {1 − [(N_e_ × T_d_)/(N_d_ × T_e_)]} × 100, where T_e_ = TiO_2_ concentration in excreta (% dry matter, DM), T_d_ = TiO_2_ concentration in the diet (% DM), N_e_ = nutrient concentration in excreta (% DM), and N_d_ = nutrient concentration in the diet (% DM).
Following the 72 h period of excreta collection on day 46, ducks were fed for 4 h, and then euthanized with carbon dioxide (CO_2_). Ileal digesta was collected from two birds per cage, pooled, and freeze-dried at −50 °C for three days to allow for subsequent analyses of TiO_2_ and amino acids (AAs). The AA contents were analyzed with an automatic amino acid analyzer (L-8900, HITACHI, Tokyo, Japan), in accordance with the method as described by Zhang et al. [17]. These data were used to calculate SIDAA based on our previous study [18].
2.7. Statistical Analyses
Data were subjected to Shapiro–Wilk and Levene’s tests to assess for normal distribution and variance homogeneity, respectively, with the SAS 9.4 software [19]. Two-tailed unpaired t-test or Mann–Whitney U test for normally or non-normally distributed datasets, respectively, was employed to ascertain statistical differences in parameters between the HS and control groups (NC and PF). The probability of p < 0.05 was considered to be statistically significant, and 0.05 < p < 0.1 was considered a trend. Data are shown as the mean ± standard error.
3. Results
3.1. Nutrient Utilization
As shown in Table 4, HS notably reduced the apparent utilization of dietary energy, EE, and CP, as well as apparent metabolizable energy (AME) when compared to the NC and PF groups (p < 0.05). Furthermore, the apparent utilization of dietary DM, TP, and Ca in the HS group was significantly lower than that in the PF group (p < 0.05).
3.2. Standardized Ileal Digestibility of Amino Acids
The Standardized Ileal Digestibility values of eight non-essential amino acids (Asp, Ser, Glu, Gly, Ala, Cys, Tyr, and Pro), seven essential amino acids (Thr, Val, Ile, Leu, Phe, His, and Arg), total non-essential amino acids (Total NEAAs), total essential amino acids (Total EAAs), and total amino acids were significantly higher in the HS group compared with those in the NC group (p < 0.05, Table 5). It was observed that ducks in the HS group had a tendency to increase the SID of Tyr in comparison to the PF group (p = 0.053).
3.3. Nutrient Transporter Gene Expression
A reduction in the mRNA levels of excitatory amino acid transporters 3 (EAAT3) and oligopeptide transporter 1 (PepT1) was observed in the HS group in comparison to the NC and PF groups, respectively (p < 0.05, Figure 1). Nevertheless, HS notably up-regulated the mRNA expression of cationic amino acid transporter 1 (CAT1), glucose transporter 5 (GLUT5), and fatty acid transporter protein 6 (FATP6) compared to the NC and PF groups (p < 0.05).
3.4. Intestinal Permeability and Morphology
The serum FITC-d concentration of ducks in the HS group was significantly higher than that in the NC group (p < 0.05, Figure 2A), but was similar to that in the PF group. The jejunal morphology of ducks under HS conditions was impaired, as evidenced by lower VH, VH/CD, and goblet cell count, in addition to higher CD when compared to the NC and PF groups (p < 0.05, Figure 2B–F).
3.5. Changes in Ileal Mucosal Microbiota
The effects of HS on the microbiota in the ileal mucosa are illustrated in Figure 3. The Shannon index in the HS group was not significantly different compared to that in the NC and PF groups (p > 0.05, Figure 3A). Principal component analysis (PCA) demonstrated that the samples in the HS group did not form a distinct cluster that was clearly separated from those in the NC and PF groups (Figure 3B). The specific taxa in the microbiota of ileal mucosa that were significantly associated with HS were identified by Linear discriminant analysis Effect Size (LEfSe ≥ 2, family level). Actinomycetaceae, Microbacteriaceae, and Rhizobiaceae were notably enriched in the HS group, while Enterobacteriaceae was significantly enriched in the NC group (Figure 3C). More specifically, HS increased the relative abundance of Bacillales, Deferribacterales, and Actinomycetales at the order level compared to the NC group, whereas it notably reduced the relative abundance of Bifidobacteriales in comparison to the PF group (p < 0.05, Figure 3D–G).
4. Discussion
Several studies have reported that the apparent utilization of DM, CP, and energy in the diet were significantly reduced in heat-stressed broilers compared to those under thermoneutral conditions [20,21]. In a study by Bonnet et al. [22], which excluded the impact of FI by using pair-feeding techniques, it was observed that HS also decreased the apparent utilization of dietary DM, CP, and EE in broilers. In accordance with these previous studies, our findings suggest that HS led to a reduction in the apparent utilization of dietary DM, energy, EE, CP, Ca, and TP. Wallis and Balnave [23] observed a slight decrease in the apparent ileal digestibility (AID) for Thr, Ala, Met, Ile, and Leu under HS conditions (31 °C, 24 h/d, 29 d). Teyssier et al. [2] revealed the detrimental effect of HS (35 °C, 24 h/d, 21 d) on AA digestibility, with an average reduction of 5% in AID of total AAs. However, there was a tendency for the SID of Tyr to increase in the HS group compared to the PF group in this study. This difference may be due to the fact that ducks have a more developed digestive tract compared to chickens [24,25], and thus HS exerts less impact on the SIDAA of meat ducks. In our experimental setting (34 ± 1 °C, 7 h/d, 14 d), heat-stressed ducks exhibited slightly higher SIDAA in accordance with an elevated CAT1 mRNA level in the jejunal mucosa, potentially compensating for enhanced AA requirements. Numerous studies have demonstrated significant upregulation of amino acid and small peptide transporters in the small intestine of heat-stressed broilers and swine [11,26]. In the present study, we observed marked upregulation of the CAT1 compared to both the PF and NC groups. This phenomenon exhibited a consistent trend with GLUT5 upregulation, potentially serving as a compensatory mechanism to address glucose supply deficits in heat-stressed meat ducks [4]. The maintenance of SIDAA under HS is attributed to the combined effects of reduced FI and intestinal compensatory mechanisms, with adaptive changes in the gut playing the predominant role. However, we found that HS resulted in reduced CP utilization in the diet. This paradoxical phenomenon may be partially attributed to hindgut damage causing increased endogenous nitrogen loss or microbiota dysbiosis reducing protein fermentation during the HS condition, thereby reducing the apparent utilization of CP.
In a study by He et al. [27], it was demonstrated that HS negatively affected intestinal morphology, resulting in a reduction in VH and VH/CD in the jejunum and ileum of ducks. Similarly, the results of our study showed that HS impaired jejunal morphology of ducks, as illustrated by reduced VH and VH/CD, as well as increased CD. The intestinal epithelia are covered with mucus secreted by goblet cells that can prevent intestinal mucosa from pathogen attacks and environmental toxins [8]. In the present study, HS significantly reduced jejunal goblet cell count, in agreement with the findings of Liu et al. [8] and Zhang et al. [28]. Further, FITC-d has been used as an indicator for the assessment of intestinal paracellular permeability, and the elevated concentration of FITC-d in serum implies impaired intestinal barrier function [29]. In the present study, the serum FITC-d levels of the HS group were found to be notably higher than those of the NC group, but similar to those of the PF group. Gilani et al. [30] showed that both a 4.5 h and a 9 h feed restriction period led to increased FITC-d concentrations in the blood of broilers. These findings indicate that HS may disrupt the intestinal barrier function in ducks by reducing FI. As demonstrated by Koch’s research, HS (Temperature–Humidity Index = 76) directly induced jejunal barrier damage in bovines compared to the PF control [31]. However, this effect was not observed in the current experiment, suggesting that triggering such responses may require either prolonged exposure duration or elevated temperature conditions.
Gut microbiota plays an indispensable role in intestinal mucosal homeostasis and gut health [32]. Several studies observed higher alpha diversity indices of microbiota in ileal contents and mucosal scrapings in chickens raised under high ambient temperature (HT) compared to thermoneutral conditions. However, our results showed that HS did not influence the alpha diversity of the microbiota in the ileal mucosa of ducks. Patra and Kar [33] highlighted that the effect of HT on the alpha diversity of gastrointestinal tract microbiota is contingent upon the duration and intensity of heat exposure. Previously, it was observed that the microflora structure of ileal content in chickens was altered by HT, based on unweighted UniFrac distance metric matrices [34]. Nevertheless, the results of our experiment in ducks demonstrated that HS had no effect on the microbiota structure in the ileal mucosa, in accordance with the findings of [35]. Xing et al. [36] posited that HT had a significant impact on the microbial structure of laying hens, which was primarily associated with a reduction in FI. The discrepancies observed in the aforementioned studies may be partly explained by the fact that intestinal mucosal flora is not easily affected by FI. The relative abundance of Deferribacterales was found to be significantly elevated in both mouse [37] and porcine [38] models suffering from inflammatory bowel disease and may release lipopolysaccharides to trigger inflammatory injuries and aggravate energy metabolism abnormalities [39]. In this study, we observed that HS markedly increased the relative abundance of Deferribacterales, whereas it notably reduced the relative abundance of Bifidobacteriales. Similarly, previous studies reported that HS decreased the populations of Bifidobacterium in the jejunal digesta of broilers [28,40]. These results suggest that HS increased harmful bacteria but decreased beneficial bacteria in the ileal mucosa at the order level.
5. Conclusions
Collectively, chronic HS-induced intestinal morphological injury, heightened intestinal permeability, and ileal mucosal microbiota dysbiosis contributed to reduced dietary nutrient utilization in Pekin ducks. The detrimental impact of FI reduction on intestinal health underscores FI regulation as a promising nutritional strategy to alleviate HS challenges in meat duck production. These insights into HS-driven interplay between gut health and nutrient utilization hold relevance for broader poultry species amid climate change, though strategy adaptation will be necessary to account for species-specific thermotolerance and intestinal physiology.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Deeb N. Shlosberg A. Cahaner A. Genotype-by-environment interaction with broiler genotypes differing in growth rate. 4. Association between responses to heat stress and to cold-induced ascites Poult. Sci.2002811454146210.1093/ps/81.10.145412412909 · doi ↗ · pubmed ↗
- 2Teyssier J.R. Cozannet P. Greene E. Dridi S. Rochell S.J. Influence of different heat stress models on nutrient digestibility and markers of stress, inflammation, lipid, and protein metabolism in broilers Poult. Sci.202310210304810.1016/j.psj.2023.10304837797358 PMC 10613759 · doi ↗ · pubmed ↗
- 3Teyssier J.R. Preynat A. Cozannet P. Briens M. Mauromoustakos A. Greene E.S. Owens C.M. Dridi S. Rochell S.J. Constant and cyclic chronic heat stress models differentially influence growth performance, carcass traits and meat quality of broilers Poult. Sci.202210110196310.1016/j.psj.2022.10196335709683 PMC 9201008 · doi ↗ · pubmed ↗
- 4Zeng X. Javid A. Tian G. Zhang K. Bai S. Ding X. Wang J. Lv L. Xuan Y. Li S. Metabolomics analysis to interpret changes in physiological and metabolic responses to chronic heat stress in Pekin ducks Sci. Total Environ.202491216938210.1016/j.scitotenv.2023.16938238110095 · doi ↗ · pubmed ↗
- 5Shi F.Y. Guo N. Degen A.A. Niu J.H. Wei H.Y. Jing X.P. Ding L.M. Shang Z.H. Long R.J. Effects of level of feed intake and season on digestibility of dietary components, efficiency of microbial protein synthesis, rumen fermentation and ruminal microbiota in yaks Anim. Feed Sci. Technol.202025911435910.1016/j.anifeedsci.2019.114359 · doi ↗
- 6Findeisen E. Südekum K.H. Fritz J. Hummel J. Clauss M. Increasing food intake affects digesta retention, digestibility and gut fill but not chewing efficiency in domestic rabbits (Oryctolagus cuniculus)J. Exp. Zool. Part A 202133561462210.1002/jez.2505 PMC 836211234254468 · doi ↗ · pubmed ↗
- 7Ducatelle R. Goossens E. Eeckhaut V. Van Immerseel F. Poultry gut health and beyond Anim. Nutr.20231324024810.1016/j.aninu.2023.03.00537168453 PMC 10164775 · doi ↗ · pubmed ↗
- 8Liu L. Fu C. Yan M. Xie H. Li S. Yu Q. He S. He J. Resveratrol modulates intestinal morphology and HSP 70/90, NF-κB and EGF expression in the jejunal mucosa of black-boned chickens on exposure to circular heat stress Food Funct.201671329133810.1039/C 5FO 01338 K 26843443 · doi ↗ · pubmed ↗