Does Short-Distance Migration Facilitate the Recovery of Black-Necked Crane Populations?
Le Yang, Lei Xu, Waner Liang, Jia Guo, Yongbing Yang, Cai Lyu, Shengling Zhou, Qing Zeng, Yifei Jia, Guangchun Lei

TL;DR
Short-distance migration in Black-Necked Cranes may help their population grow by saving energy and reducing risks during travel.
Contribution
The study identifies a low-energy, high-efficiency migration strategy in a growing subpopulation of Black-Necked Cranes.
Findings
Black-Necked Cranes in northern Tibet migrate short distances with minimal stopover reliance.
Autumn migration is shorter, higher, and slower compared to spring migration.
Younger cranes use smaller, fragmented habitats that expand as they mature.
Abstract
Black-necked Cranes are high-altitude birds that only grow and breed on plateaus. Understanding how they migrate is important for their conservation. In this study, we tracked 16 cranes from northern Tibet using satellite tags. We found that these cranes presented shorter migration distances and were less reliant on stopover sites compared to other subpopulations. The autumn migration was shorter, higher in altitude, and slower in speed, while spring migration was longer and more complex. Younger cranes used smaller and more scattered habitats, which gradually expanded as they grew older. We deduce that short-distance migration may help cranes conserve energy, decrease mortality rate during migration, and allow more flexibility in migration routes. This could be a driver for the continued growth of this subpopulation, which could support population growth in the greater region by…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
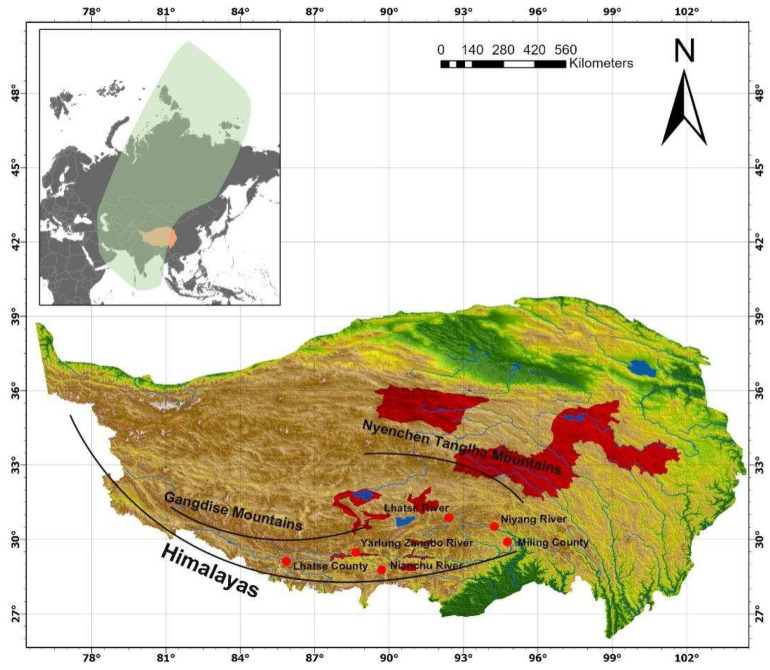 Figure 1
Figure 1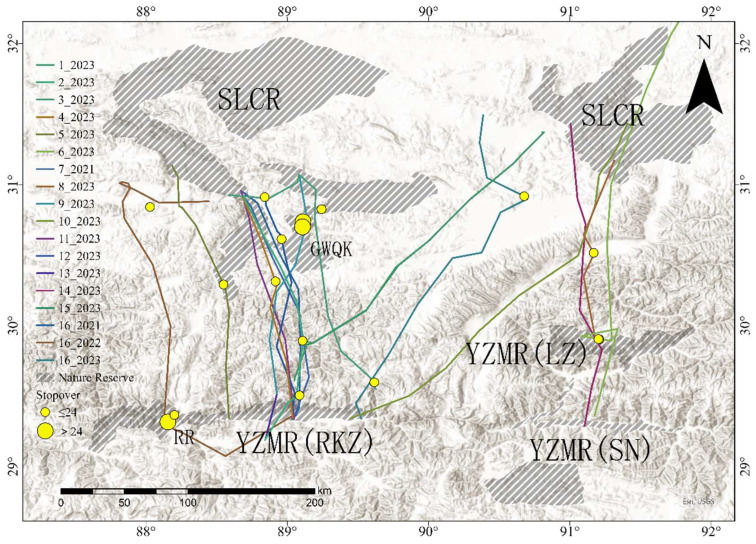 Figure 2
Figure 2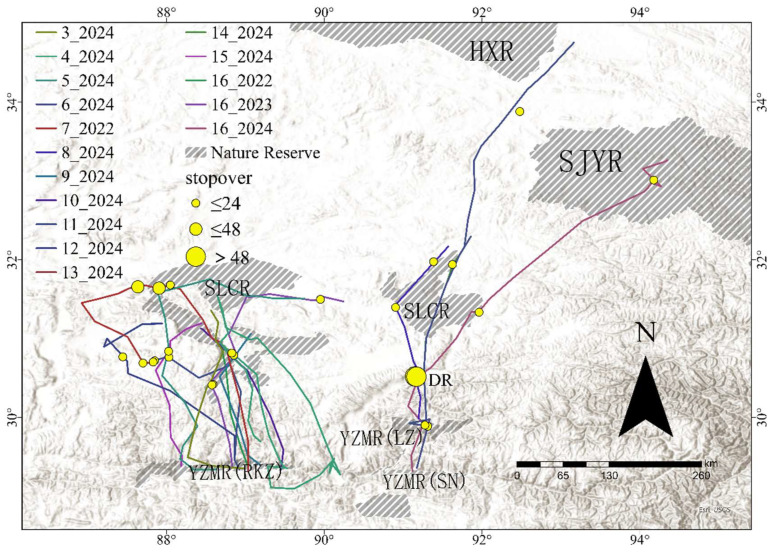 Figure 3
Figure 3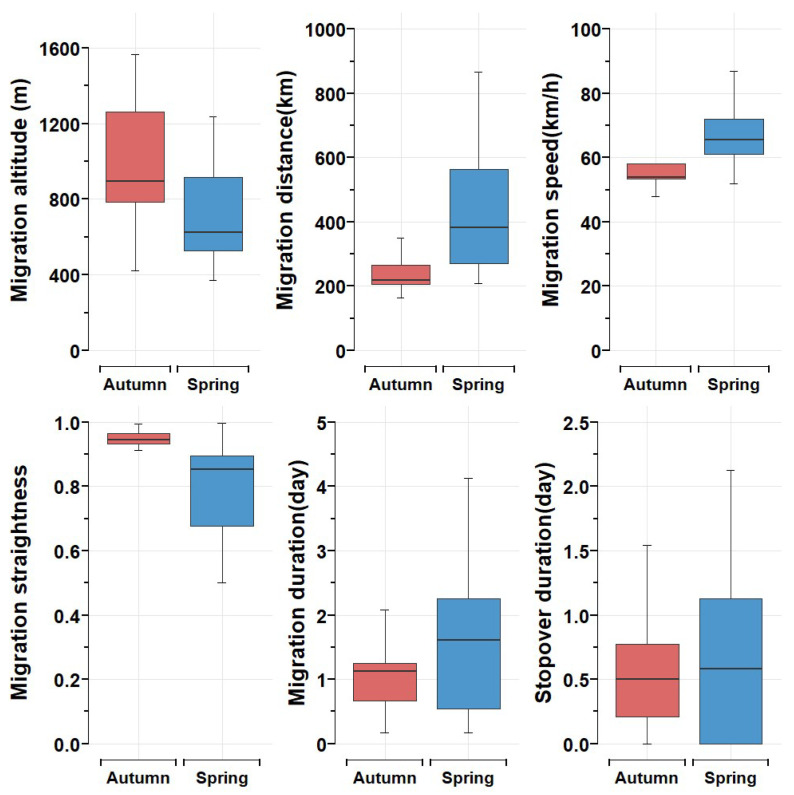 Figure 4
Figure 4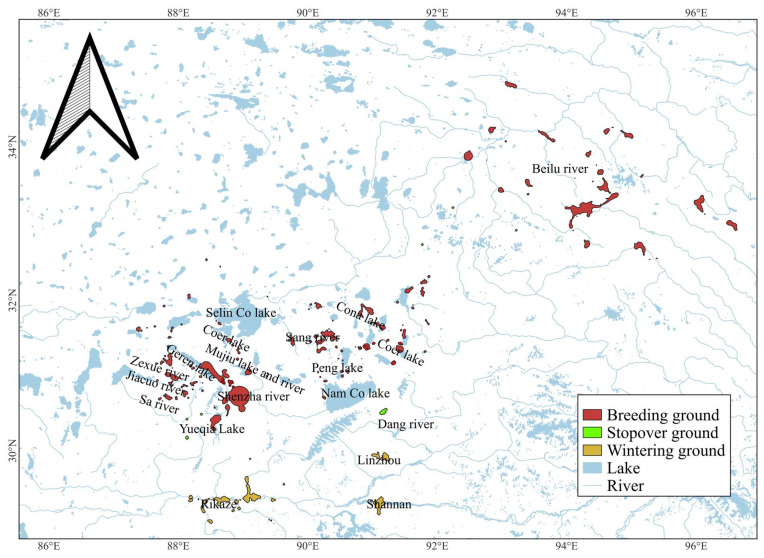 Figure 5
Figure 5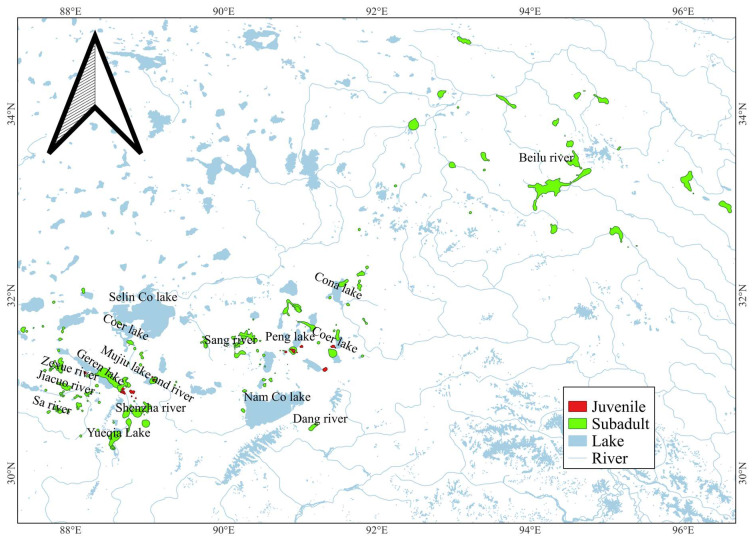 Figure 6
Figure 6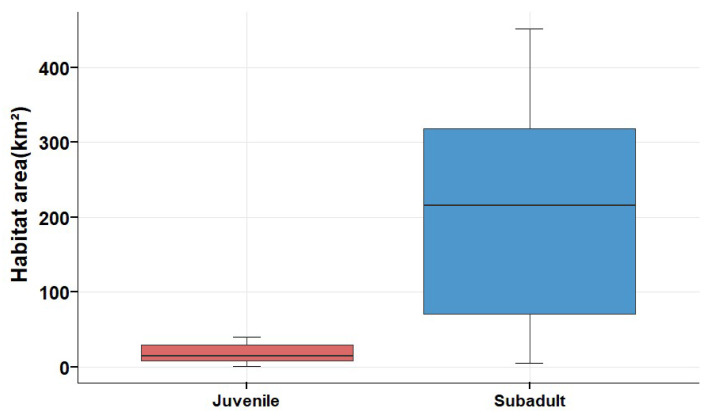 Figure 7
Figure 7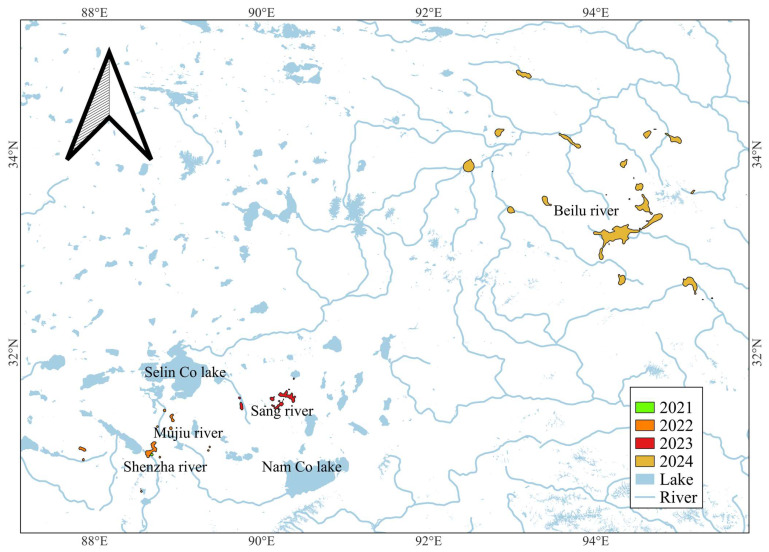 Figure 8
Figure 8- —the Second Tibetan Plateau Scientific Expedition and Research Program
- —Key scientific and technological research and development projects in Xizang Autonomous Region
- —Special Project for Central Guiding Local Science and Technology Development
- —Special Project for Central Guiding Local Science and Technology Development
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFish Ecology and Management Studies · Marine Ecology and Invasive Species · Aquatic Invertebrate Ecology and Behavior
1. Introduction
Migration dynamics, wintering and breeding ecology, and stopover site usage are important aspects of migratory waterbird studies, and are crucial for the conservation of migratory waterbird species and their habitat networks [1]. Migration in waterbirds, similar to other migratory taxa, is a strategy for tracking food resources to better conduct activities as breeding and wintering, as well as to mitigate competition and avoid predation. Therefore, it has long been a core topic for conservationists on how to manage the threats to the migratory routes of a waterbird species, and coordinate all stakeholders to conduct conservation actions [2]. Waterbirds are sensitive to environmental change, and have been considered as bio-indicators for ecosystem health and conservation effectiveness [3]. Among the nine flyways around the world, the Central Asian Flyway is the least studied flyway, even though it is one of the most threatened, with 40% of its species in decline [4]. Albeit being the shortest, it spans across a large range of countries and a variety of terrains. Long-term, large-scale studies on migratory species on this flyway are scarce on this flyway, and there’s a lack of research on the population trends and migratory strategies of its species.
The scattered wetlands on the Tibetan Plateau give it the title of Asian Water Tower. It supports migratory waterbirds with its chains of wetlands as roosting and foraging sites [5] and is a key component of the Central Asian Flyway. However, due to its high altitude of 4000 m, it is one of the most impacted regions under global warming, challenging bird survival through its extreme cold, low humidity and reduced air density [6,7,8]. Waterbird species have adapted unique migratory strategies tailored to this environment [9]. Studies in this field have been concentrated on the more famous migrations, including the flight over the Himalaya mountains [10]. In contrast, studies conducted on the species endemic to the Tibetan Plateau were scarce.
Black-necked Cranes (Grus nigricollis) are the only crane species that spends its entire life history on plateaus out of the total of 15 crane species in the world. It is also the last crane species to be discovered [11]. It is a first-class protected animal in China, and is classified as Near Threatened by the International Union for Conservation of Nature (IUCN) Redlist [12]. The Black-necked Crane consists of three populations, namely the western population, the central population and the eastern population. The 10,000-individual Western population in Tibet is the largest. Black-necked Crane is considered an important eco-indicator and a flagship species for the alpine wetland ecosystem [13]. The species is ideal for studying the migratory strategy of birds at high altitudes. So far, past research has discovered 14 unique migratory routes for the Black-neck Crane among its three populations [14,15,16,17,18,19,20]. Those studies provide insights into the long-distance migration on the Qinghai-Tibetan Plateau. However, the migration of the Black-necked Cranes of the western population is still limited, especially the breeding subpopulation of the northern Tibetan lake basin (NT), which has been increasing rapidly (Yang, unpublished data). To resolve this problem, this research used satellite tracking to explore the migration route of Black-necked Cranes in the NT subpopulation. We aim to: (1) Identify the migration route of the subpopulation, (2) analyse the patterns of this migration route between spring and autumn migration, and between juveniles and subadults.
2. Materials and Methods
2.1. The NT Subpopulation
In this study, we focused on the NT breeding subpopulation of the Black-necked Crane. The NT subpopulation is a part of the Western population. Previous studies [21,22] have shown that the NT subpopulation breeds in Selin Co National Reserve (SLCR), which includes Selin Co Lake, Shenzha River, Pengco Lake, among others, with an average altitude of the 4500 m; it migrates the Black-necked Crane National Nature Reserve on the Middle Reaches of the Yarlung Tsangpo River in Tibet Autonomous Region (YZMR) in Southern Tibet, with an average altitude of 3500 m. The subpopulation mostly spends all its life stages inside Tibet (Figure 1); however, migrants from and to the breeding communities of Hoh Xil National Nature Reserve (HXR) (Western Qinghai, with an average altitude of 4600 m); Sanjiangyuan National Nature Reserve (SJYR) (Southern Qinghai, with an average altitude of 4000–4800 m) has been recorded.
Breeding surveys during 2021–2024 showed an increase in the subpopulation. Breeding surveys covered known breeding locations in the NT from past records or reported by local communities. Some known locations were not included in the survey due to logistical issues. In 2021, 59 breeding pairs were found; in 2022, 62 breeding pairs, and in 2023, 75 breeding pairs.
2.2. Bird Capturing and Satellite Tracking
We captured and satellite tracked a total of 16 Black-necked Cranes in SLCR during the breeding season of 2021 and 2023. Fifteen of them were flightless juveniles, and one was a moulting adult. Satellite trackers (HQLG4037S, Hunan Global Messenger Technology Co., Ltd. (Changsha, China), weighing 37~44 g) were attached to the right upper leg of the individuals. Birds were weighted and made sure the satellite trackers weighed less than 3% of each bird’s body mass to minimise the impact on its behaviour [23,24]. Location, speed, direction, altitude, and temperature data were transmitted through the GSM system every hour or every three hours, depending on battery efficiency. Bird capture, tracking, and tagging were conducted under the permits of the National Forestry and Grassland Administration, China (Permit No. Lin Hu Xu Zhun (2021) 223; issued on 31 December 2021).
The satellite data accuracy was classified into 5 levels: levels A (error within 5 m), B (5–10 m), C (10–20 m), D (20–100 m), E (>100 m). We only used data with an accuracy level of C and above to ensure credibility. From 2021 to 2024, we collected 147,782 location data points from the 16 tracked individuals (Table 1).
2.3. Routes and Seasonal Differences in Migration
Two stages were identified from the tracking data: staging or flying. Staging data was characterised with a speed of 0, and the flying data was characterised with a speed faster than 10 km/h. Using the classified data, through the Geometric function in ArcGIS Pro 3.5.1, we constructed the migration route of the tracked Black-necked Crane individuals. Among the individuals, only one died before completing a migration cycle; all the others migrated at least one cycle. This resulted in 16 complete spring migrations and 18 complete autumn migrations.
We also calculated the migration distance (accumulated distance), migration straightness (shortest distance between start and end divided by accumulated distance), migration speed (accumulated distance/total migration time), flight height (flight altitude subtracted by corresponding terrain elevation) for each migration event [16,25]. The start and end of a migration event were defined as the time when an individual arrived at/departed from a breeding/summering/wintering ground.
We identified stopover sites used in each migration event. A stopover site was defined as a location during a migration event where the individual stayed for more than six hours within a 10 km^2^ area. The stopover duration was calculated for each site. A stopover does not necessarily indicate a refuelling site; it may also serve other purposes such as resting or waiting for favourable conditions.
Statistical tests were performed on the above parameters between spring and autumn migration. For data with a normal distribution (p > 0.05 in the Shapiro–Wilk test), we performed t-tests. Conversely, for data without a normal distribution (p < 0.05 in the Shapiro–Wilk test), we performed Wilcoxon rank-sum tests. All statistical analyses were conducted in R version 4.2.2 [26], using the stats package [27], which is included in the base R distribution.
2.4. Analysis of Habitat Utilization and Distribution
We used the Dynamic Brownian Bridge Movement Models (DBBMM) from the move package [28] of R to construct the utility distribution (UD) of each migration event. UD uses percentages to describe the probability distribution of an animal’s spatial use, i.e., the probability of presence of said animal in a certain location. We used 50% and 95% UD in our study to represent the habitat use for each migration event. The areas of 50% UD and 95% UD were calculated. The habitat area (95% UD) was compared between juvenile and subadult, and a Wilcoxon rank-sum test was conducted.
3. Results
3.1. Migration Route and Seasonal Variation
Most individuals (n = 12) showed a consistent migration route between years. The migration of the NT subpopulation was characterised by its short distance, short duration and low dependency on stopover sites and high inter-individual variations.
During autumn migration (Figure 2), stopover sites were used in 16 out of 18 migration events. In total, 16 stopover sites were identified for autumn migration, with an average of 0.89 ± 0.58 stopover sites used per autumn migration event. The stopover duration per site was an average of 1.69 ± 1.87 days. All autumn migrations were conducted within Tibet, starting from the wetlands in SLCR, and ending at the wintering site of YZMR. Three major routes can be concluded between the individuals: ① West of SLCR (Shenzha river, Geren lake, the river of Mujiu, etc.) -YZMR (Rikaze, RKZ) (n = 11); ② East of SLCR (Peng Lake, Coer lake, Cuona Lake, etc.) -YZMR (Linzho, LZ/Shannan, SN) (n = 3); ③ East of SLCR-YZMR (RKZ) (n = 4).
During spring migration (Figure 3), stopover sites were used in 12 out of 16 migration events. In total, 13 stopover sites were identified for spring migration, with an average of 2.16 ± 0.93 stopover sites used per spring migration event. The stopover duration per site was an average of 3.28 ± 7.04 days. 14 spring migrations were conducted within Tibet. Two migration events were conducted across both Tibet and Qinghai. The spring migration routes within Tibet were similar to the autumn migration routes described above, while the two involving Qinghai were: ① YZMR (SN)-HXR (the Beilu River) (n = 1); ② YZMR (SN)-SJYR (Zhana River) (n = 1).
The autumn migration was conducted from 18 October to 6 November. The spring migration was conducted from 22 March to 25 May. Comparison between autumn and spring migration (Figure 4) showed significant difference in migration distance (autumn: 235.58 ± 45.87 km; spring: 428.70 ± 188.42 km; p = 0.0001), migration altitude (autumn: 979.74 ± 362.03 m; spring: 696.73 ± 248.07 m; p = 0.012), migration speed (autumn: 56.64 ± 7.91 km/h; spring: 181.8 ± 93.2 km/h; p = 0.002), migration straightness (autumn: 0.89 ± 0.13; spring: 0.75 ± 0.21; p = 0.002). There was no significant distance in migration difference (autumn: 1.69 ± 1.67 days; spring: 4.15 ± 7 days; p = 0.27) and stopover duration (see above, p = 0.66) between the two seasons. Overall, the spring migration was longer in distance, faster in-flight speed, the route was less straight and direct, and flight was lower.
Among the identified stopover sites, the majority were utilized by only a single individual, typically located in small and fragmented wetlands such as tributaries or sub-lakes. In contrast, a few sites were used by multiple individuals, either during different migration seasons or across different years. For example, the GeWangQueKang was used exclusively by individuals 9 and 13 during separate autumn migrations. The Dangqu River was visited by individuals 8 and 16, but in different years. Linzhou county, located near the wintering grounds, functioned as a key corridor node and was used repeatedly: by individuals 6 and 14 during autumn migration in 2023, and again by individual 6 during spring migration in 2024. Notably, simultaneous use of Linzhou was observed, with individuals 6 and 14 occupying the site during overlapping migration periods across both spring and autumn seasons.
3.2. Habitat Use
Our results from the DBBMM models identified 252 habitats used (Figure 5). The total habitat area (95% UD) was 6773.99 square kilometres, and the core area used (50% UD) comprised 131.6 square kilometres. For wintering grounds, the total area was 2077.80 square kilometres, and the core area was 82.5 square kilometres. The wintering grounds were mainly concentrated at the middle reaches of the Yarlung Zangbo River valley (Rikaze area and Shannan City’s Jiedexiu Forest National Park) and at the Kazi Reservoir and Hutoushan Reservoir in Linzhou County, Lhasa City. For breeding grounds, the total area was 49,696.18 square kilometres, and the core area was 47.27 square kilometres. The breeding area includes sub-lakes and rivers in northern Tibet, such as Shenzha River–Geren Lake, Coer Lake, Mujiu Lake and river, Sang River, Peng Lake–Coer Lake, Yueqia Lake and Beilu River, etc. Stopover sites have a total area (95% UD) of 111.32 square kilometres, including the Sa River, Nam Co Lake–Dang River, and Zexue River–Jiacuo River.
The habitats used by juveniles were very different from subadults (Figure 6). The area used was also significantly different according to the result of the Wilcoxon rank-sum test (p < 0.01, Figure 7). Habitats (95% UD) used each year for each juvenile were at 17.48 ± 11.84 square kilometres, 206.99 ± 156.56 square kilometres for each subadult.
4. Discussion
4.1. Autumn and Spring Migration
Black-necked Cranes are altitude-driven migratory species, instead of latitude-driven [18]. This results in a difference in fundamental energy requirements depending on whether the migration is travelling towards higher or lower altitudes. In autumn, Black-necked Cranes in Tibet migrate from high breeding grounds to lower altitude wintering grounds where resources are more available. Conversely, spring migration requires Black-necked Cranes to travel to locations with higher altitudes, resulting in more energy consumption. In many species, spring migration duration is shorter due to the fact that early arrival on the breeding ground allows for higher breeding success [29,30]. Our study did not show a faster spring migration. Spring migration consisted of a longer migration distance and a less direct route, which is compensated by higher migration speed, resulting in a similar migration duration to autumn migration. The longer migration distance and the detours are likely due to the very fact that spring migration requires more energy in order to raise altitude. In autumn migration, the migration from high altitude to low altitude might allow for more effective flights. In spring, it needs to climb, and therefore is more challenging. Altitudinal changes might not be the only reason for longer and less direct spring migration routes. In Tibet, the wind direction is mostly southward [31], which aids in the southward autumn migration, and is more likely to hinder the northward spring migration. The less straight routes in spring migration might be a result of Black-necked Cranes avoiding difficult terrain to allow for quicker flight.
4.2. Juveniles and Subadults
In our study, juveniles used a very limited habitat area initially, and the area used grew as they became subadults. The individual ID = 16 in our study were tracked for four consecutive years, which showed low breeding/summering site fidelity. Such explorative behaviour in juveniles [32,33,34] could be a key driver of population expansion [24]. In the first year of ID = 16, its habitat use was a mere 11.02 square kilometres (2021), in the second year, this expanded to 221.18 square kilometres (2022); the third year, 254.59 square kilometres (2023); and the fourth year finally expanded to 2736 square kilometres (2024). In the first three years, it only stayed in its birthplace of the typical lake basin area in northern Tibet. In the fourth-year wintering period (2023 winter), it expanded to migrate from wintering grounds west of YZMR to wintering grounds east of YZMR. And only in the following spring did it migrate to SJYR in Qinghai for summering, and thus in 2024, its habitat area expanded dramatically. SJYR is the main breeding ground for another subpopulation of the Western population of the Black-necked Crane. This result shows that juveniles might be a key factor in facilitating inter-subpopulation communication (Figure 8).
4.3. Short-Distance Migration Facilitates Fast Population Recovery
The overall population trend of the Black-necked Crane has been partially increasing recently [35]. According to previous surveys [36], in 2022, the breeding population of Black-necked Cranes in Qinghai-Tibet Plateau was over 11,800; the breeding population in Yunnan–Guizhou Plateau (east population) was over 6000 individuals. Those numbers have increased compared to the numbers in 2012/2013 (over 6000 in Qinghai–Tibet Plateau; over 3900 in Yunnan–Guizhou Plateau) [37,38]. Compared to the eastern population (annual increase of 4.1%), the western population showed a faster growth rate (annual increase of 9.7%). We believe the main driver of the recent population growth in this species is the increase in suitable habitat. Climate change induced an overall trend of an increase in wetland area in Tibet [39,40,41] due to melting glaciers [42]. The expansion of wetlands provides more suitable habitats for the foraging, breeding, wintering and other activities throughout all the life stages of the Black-necked Crane.
The growth in the NT subpopulation is even higher. According to a breeding survey conducted during our study period, the annual growth rate of the NT subpopulation was an average of 12.17% (unpublished data by Yang), higher than the average of the western population. We deduce that the reason for this exceptionally high population growth rate of the NT subpopulation is its short migratory distance. According to previous studies [14,15,16,17,18,19,20], the migration distance of the NT subpopulation is shorter than other Black-necked Crane populations, except for Pumqu and Longbaotan/Shaluli (Table 2). The shortest migration distance in our study was only 163.97 km (Appendix A).
Migration is one of the life stages with the highest mortality in migratory species [43,44,45]. This is largely due to the high energetic demands of long-distance travel, which can exhaust individuals, especially juveniles or those in poor condition. In addition, migrants face numerous external threats during transit [46], such as extreme weather events, habitat loss at stopover sites, and anthropogenic pressures like collisions with power lines or hunting. Shorter migration distance means less mortality, fewer ecological barriers to cross and less dependence on stopover sites for survival. Because the migration distance is short, the refuel requirement during migration is less, and thus less dependency on high-quality stopover sites, and more resilience to adverse changes. With fewer requirements on stopover sites, there is a wider range of potential stopover sites to choose from, allowing more flexibility in migration routes, which explains the high inter-individual variability in our study. With shorter migration distances, the required fuel load for migration decreases and the cost for juveniles to learn the migration route is less. Those factors combined result in the fast growth of the population in the NT subpopulation. The link between migration distance and population growth can also be backed by the higher growth rate of the western population than the eastern, as the former has a shorter migration distance. The fast-growing subpopulations or populations like the NT subpopulation have the potential to be a source population for the entire species and aid in species recovery. Our study shows that the NT subpopulation has communication with other subpopulations, and a previous study has shown that there is communication between the Western population and other populations [18]. Therefore, the possibility of the high-growing population spilling over to other populations is high. It is of high conservation value to conserve those populations as well as those in fast decline.
4.4. Habitat Protection in the Heart of the Tibetan Plateau Will Be Key to the Conservation of Waterbirds Along the Central Asian Flyway
The Black-necked Crane, a first-class nationally protected species in China, serves as an indicator species for the Tibetan Plateau and the Central Asian flyway. Thanks to a series of government protection measures, including the establishment of national-level nature reserves, ecological restoration projects, and the enactment of laws like the Law of the People’s Republic of China on Ecological Protection of the Tibetan Plateau, significant strides have been made in the restoration and protection of Black-necked Crane habitats. As a result, the crane’s IUCN Red List status was downgraded from “Vulnerable” to “Near Threatened” in 2020 [12,47]. However, despite these conservation efforts, Black-necked Cranes continue to face significant threats from climate change and human activities. Climate change has led to dramatic shifts in the wetland ecosystems on the Tibetan Plateau, with rising temperatures, altered precipitation patterns, and increased frequency of extreme weather events potentially changing the hydrological conditions of wetlands [48]. For example, while glacial melting and increased rainfall have expanded some lake areas, permafrost degradation has led to the loss of shallow wetlands crucial for crane survival [49,50]. Additionally, human activities such as infrastructure development have further exacerbated habitat degradation. In the Lhasa River Basin, hydropower projects have altered river flow, reducing shallow water areas and severely limiting suitable habitats for the Black-necked Crane [51]. These environmental changes threaten not only Black-necked Cranes but also the broader wetland ecosystem that supports various migratory species.
As a critical node along the Central Asian flyway, the quality and extent of wetlands on the Tibetan Plateau directly influence the survival and migration success of a wide array of waterbird species [4]. However, the ongoing shrinkage of wetlands, degradation of vegetation, and increasing human activities have led to a significant decline in habitat quality and a reduction in the connectivity essential for waterbird migration [52]. This fragmentation of habitats has disrupted migration routes, making it more difficult for migratory species to find suitable breeding, foraging, and stopover sites. Moreover, climate-induced changes to the timing and availability of resources, such as food and water, have further complicated the migratory behaviour of Black-necked Cranes. High-quality stopover sites, which provide rest and energy replenishment during migration, are particularly critical to reducing survival risks related to energy expenditure and habitat competition [53]. The loss of these sites exacerbates the challenges cranes face during migration, leading to increased energy expenditure, higher risks of predation, and heightened competition for limited resources. In this context, Black-necked Cranes’ adoption of a short-distance migration strategy may reduce the risks and energetic costs associated with migration, while also supporting the stability and growth of their wintering population in Tibet.
Beyond these direct threats, broader environmental changes such as the shifting of species ranges due to climate change are also of concern [54,55]. As temperatures rise, many species, including those that share habitats with Black-necked Cranes, are moving to higher altitudes or latitudes, leading to increased competition for space and resources. The northward movement of species may alter the dynamics of migration corridors, creating new challenges for Black-necked Cranes as they navigate these shifting ecological landscapes. These environmental changes are forcing Black-necked Cranes to adapt to new conditions, potentially altering their migration, timing, and behaviour [56,57,58,59]. Therefore, in the context of future climate change, short-distance migration strategies may become even more crucial for the adaptive evolution of Black-necked Cranes. To support the long-term survival and resilience of this species, it is imperative to strengthen habitat connectivity along key short-distance migration corridors, such as the typical lake basin area in northern Tibet–Yarlung Tsangpo River corridor, and to consolidate a robust network of protected areas to maintain this “low-energy, high-efficiency” migration model. Effective conservation strategies must consider the broader impacts of environmental change, and we must act quickly to protect and restore habitats, ensuring the future sustainability of Black-necked Cranes and other migratory species that rely on the Tibetan Plateau’s wetlands.
4.5. Limitations and Future Directions
Our investigation into the NT subpopulation of Black-necked Cranes has revealed a distinctive “low-energy, high-efficiency” migratory strategy within a single, stable growth group. However, by concentrating on this one subpopulation and a four-year tracking window (2021–2024), variation may exist in other regional groups or across longer timeframes. We also focused on age-related shifts in habitat use, but did not explore how sex differences or individual migration history might influence movement. Moreover, although we framed our findings against the backdrop of a warming plateau and changing wetlands, we did not explicitly integrate environmental drivers, such as climate shifts or habitat degradation, into our movement analyses.
We believe future research can continue to build on these insights by: (1) expanding to additional subpopulations—especially those undertaking longer migrations—to test the generality of the NT pattern; (2) extending tracking efforts over a decade or more to capture interannual responses to climate and land-use shifts; and (3) examining individual-level factors such as sex, age, and prior migratory experience through larger samples and complementary methods, including drone surveys and citizen science data. Such integrative approaches will deepen our understanding of the mechanisms that shape crane migration and support more targeted, climate-adaptive conservation actions.
5. Conclusions
In this study, we used satellite tracking data from 16 Black-necked Cranes over 2021–2024 to reveal the migration strategies and habitat use patterns of the NT subpopulation—a little-known but rapidly growing group within the western population. We found that the NT subpopulation undertakes short-distance migrations within Tibet, with a mere mean distance of 284.21 km. These migrations were characterized by minimal reliance on stopover sites, particularly in autumn, and high inter-individual consistency in route selection. Statistically significant seasonal differences were detected: autumn migrations were shorter, more direct, slower, and at higher altitudes, while spring migrations were longer and more fragmented, likely due to the combined effects of topographical change and differences in individual life stages and reproductive status. We also found clear ontogenetic patterns in habitat use: juveniles initially occupied small and fragmented habitats, but their spatial range expanded significantly as they matured, with one individual eventually migrating to a different subpopulation’s summering/breeding area, highlighting the potential for dispersal and gene flow between subpopulations.
These findings collectively indicate that the NT subpopulation has adopted a unique “low-energy, high-efficiency” migration strategy tailored to the harsh conditions of the Tibetan Plateau. Such a strategy may reduce energetic costs and mortality risks, increase route flexibility, and allow earlier or more predictable arrival at breeding sites—advantages that likely contribute to the observed population increase in this region. Importantly, our results underscore the conservation value of short-distance migration corridors like the typical lake basin area in northern Tibet–Yarlung Tsangpo River system, which supports these movements. As environmental pressures mount due to climate change, habitat fragmentation, and infrastructure development, safeguarding the core breeding and wintering wetlands and maintaining connectivity along these short routes will be crucial. Our study not only provides new insights into black-necked crane migratory strategies but also provides a scientific basis for the protection of black-necked cranes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Xu Y. Si Y. Wang Y. Zhang Y. Prins H.H.T. Cao L. de Boer W.F. Loss of functional connectivity in migration networks induces population decline in migratory birds Ecol. Appl.201929 e 0196010.1002/eap.196031237968 PMC 6852588 · doi ↗ · pubmed ↗
- 2Flack A. Aikens E.O. Kolzsch A. Nourani E. Snell K.R.S. Fiedler W. Linek N. Bauer H.G. Thorup K. Partecke J. New frontiers in bird migration research Curr. Biol.202232 R 1187 R 119910.1016/j.cub.2022.08.02836283388 · doi ↗ · pubmed ↗
- 3Clausen K.K. Madsen J. Cottaar F. Kuijken E. Verscheure C. Highly dynamic wintering strategies in migratory geese: Coping with environmental change Glob. Change Biol.2018243214322510.1111/gcb.1406129350875 PMC 6032841 · doi ↗ · pubmed ↗
- 4Mundkur T. Ananzeh A. Chaudhary A. Evans M. Jia Y. Koshkina A. Kumar R. Nergui J. Niven R. Rao M. Central Asian Flyway—Situation Analysis: The Status of Migratory Birds and Their Habitats and Recommendations for Their Conservation Bird Life International Cambridge, UK 2023
- 5Li D. Davison G. Lisovski S. Battley P.F. Ma Z.J. Yang S.F. How C.B. Watkins D. Round P. Yee A. Shorebirds wintering in Southeast Asia demonstrate trans-Himalayan flights Sci. Rep.2020102123210.1038/s 41598-020-77897-z 33311583 PMC 7732824 · doi ↗ · pubmed ↗
- 6Altshuler D.L. Dudley R. The physiology and biomechanics of avian flight at high altitude Integr. Comp. Biol.200646627110.1093/icb/icj 00821672723 · doi ↗ · pubmed ↗
- 7Butler P.J. High fliers: The physiology of bar-headed geese Comp. Biochem. Physiol. A-Mol. Integr. Physiol.201015632532910.1016/j.cbpa.2010.01.01620116442 · doi ↗ · pubmed ↗
- 8Scott G.R. Elevated performance: The unique physiology of birds that fly at high altitudes J. Exp. Biol.20112142455246210.1242/jeb.05254821753038 · doi ↗ · pubmed ↗