Rsp5-mediated ubiquitination of a functional analog of the Rim8 arrestin facilitates Rim pathway activation in Cryptococcus neoformans
Lukas M. du Plooy, Corinna Probst, Matthew E. Capobianco, Charles Giamberardino, Claudia A. Petrucco, J. Andrew Alspaugh

TL;DR
This study explores how the fungus Cryptococcus neoformans adapts to host conditions using a stress response pathway involving ubiquitination.
Contribution
The study identifies a new component of the Rim pathway in Cryptococcus neoformans that is targeted by Rsp5-mediated ubiquitination.
Findings
Rsp5-mediated ubiquitination is required for activation of the Rim pathway in Cryptococcus neoformans.
A new Rim pathway component analogous to ascomycete Rim8/PalF proteins was identified as a target of Rsp5 ubiquitination.
Ubiquitination facilitates interactions with Vps23, a downstream trafficking component of the Rim pathway.
Abstract
Pathogenic microorganisms use varied cellular processes to adapt to the particular stresses encountered in the infected host. These stresses include rapid alterations in ambient temperature, nutrient availability, and extracellular pH. Fungal pathogens, therefore, rely on the activation of stress response pathways such as the Pal/Rim pathway to adapt to the neutral pH encountered when infecting mammals. While this pathway is conserved among human pathogenic fungi, the proteins required for pH sensing appear to have diverged between different fungal phyla. The opportunistic fungal pathogen Cryptococcus neoformans, a basidiomycete, employs a pH-sensing protein in its Rim pathway that is distinct but functionally analogous to related proteins in well-studied ascomycete fungal systems. We recently characterized protein ubiquitination mediated through the Rsp5 ubiquitin ligase to be required…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
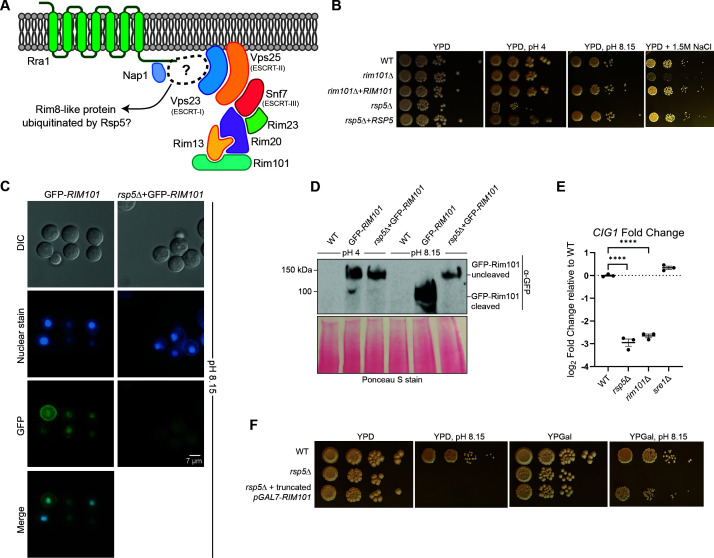 Fig 1
Fig 1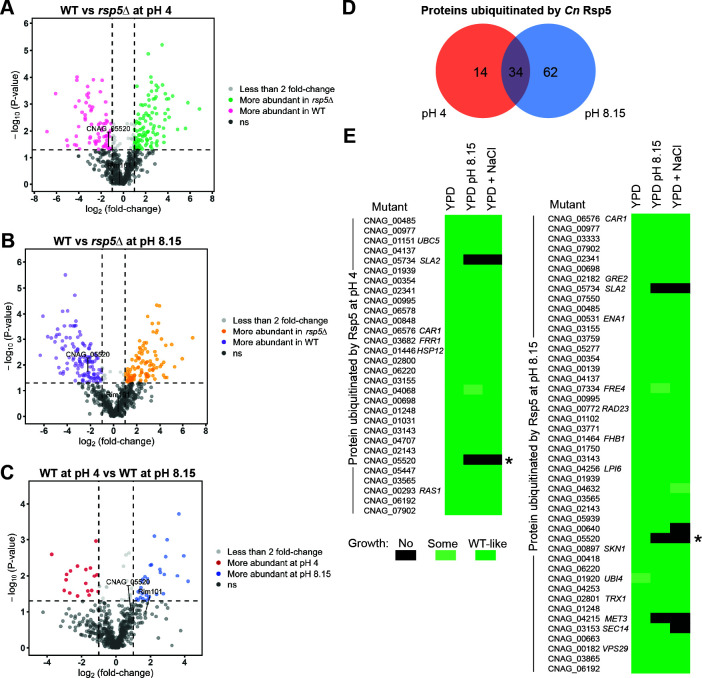 Fig 2
Fig 2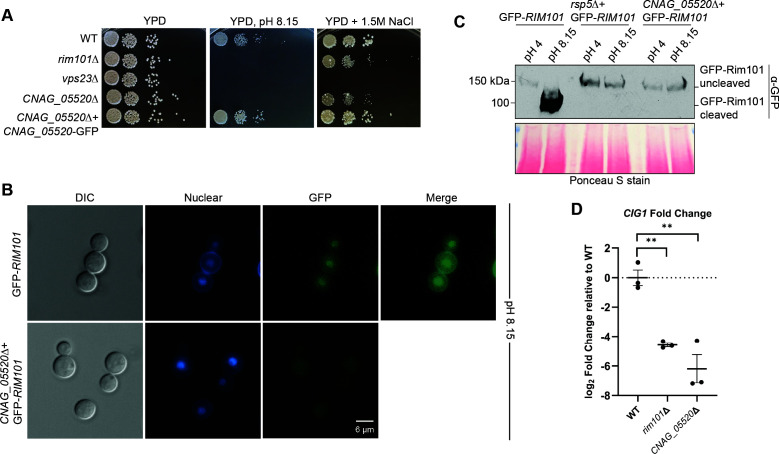 Fig 3
Fig 3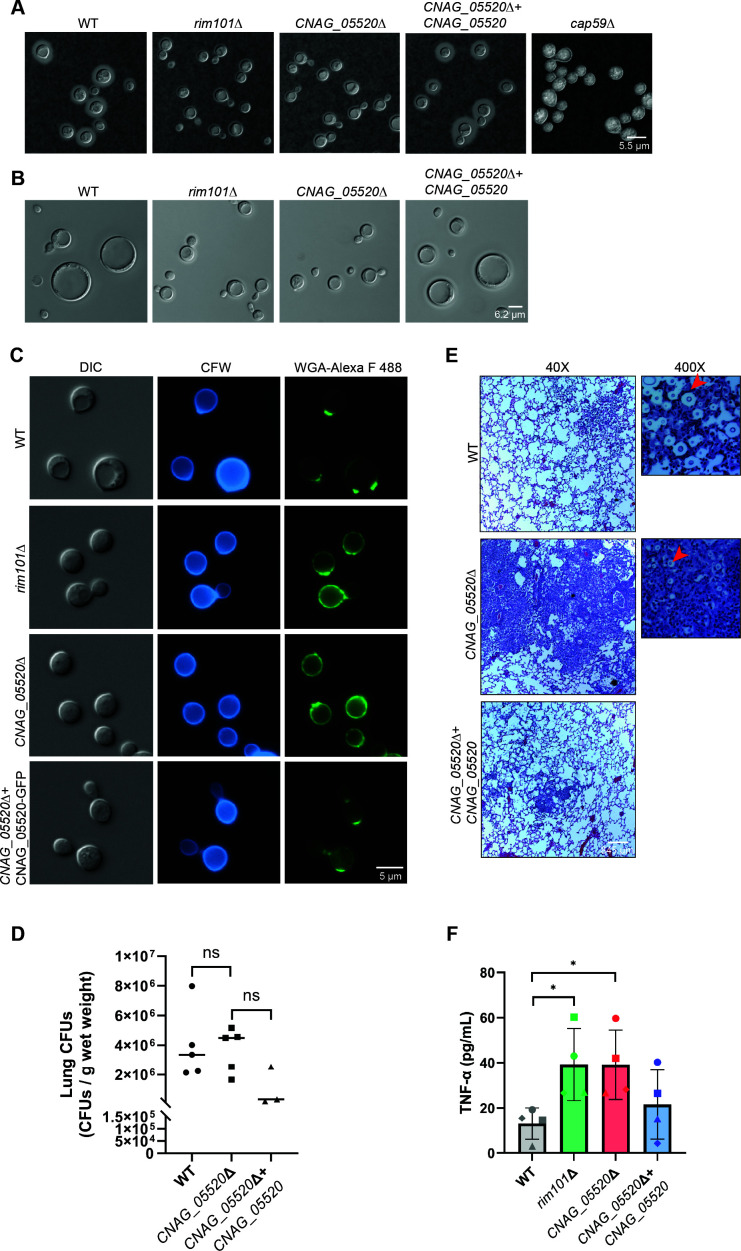 Fig 4
Fig 4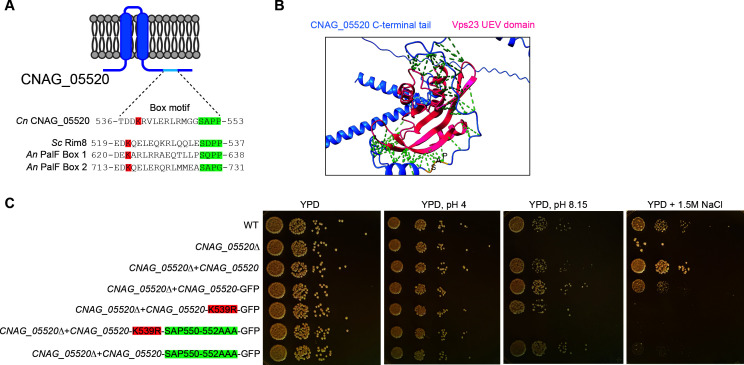 Fig 5
Fig 5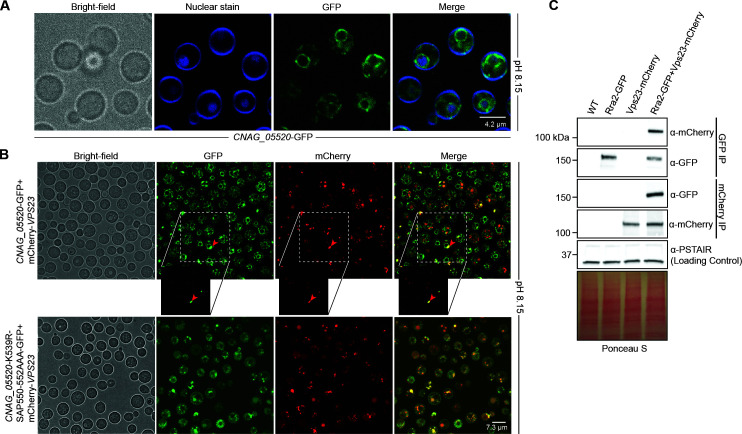 Fig 6
Fig 6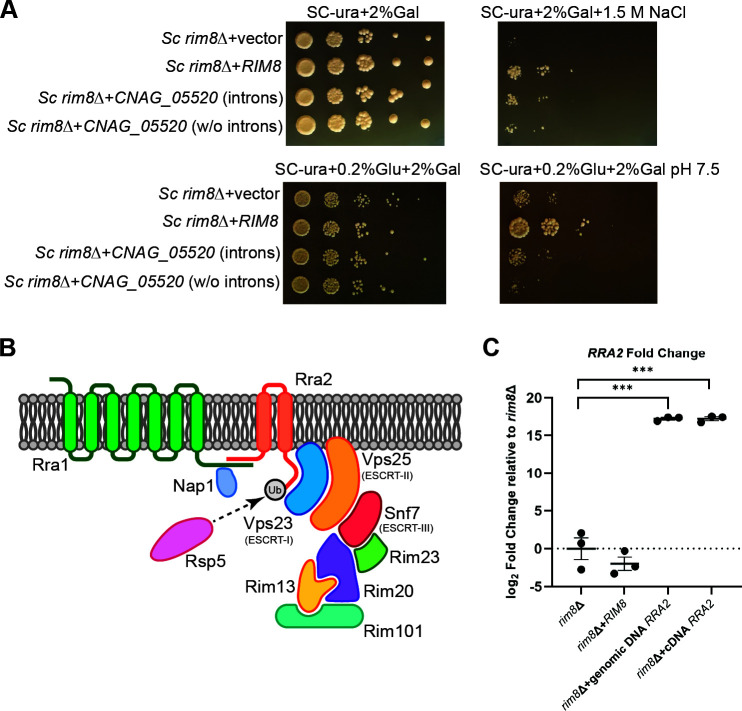 Fig 7
Fig 7- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —National Institutes of Healthhttp://dx.doi.org/10.13039/100000002
- —National Research Foundationhttp://dx.doi.org/10.13039/501100001321
- —Fulbright U.S. Student Program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFungal Infections and Studies · Antifungal resistance and susceptibility · Toxin Mechanisms and Immunotoxins
INTRODUCTION
Many of the same molecular features that favor the survival of fungi in specific environments also allow fungal pathogens to colonize and cause invasive disease in humans. For example, two well-recognized environmental stressors, rapid changes in ambient temperature and pH, are also encountered in an infected mammalian host (1–6). These ever-changing stress conditions in diverse environments, as well as local competition with other microorganisms, provide selective pressures that lead to adaptation to the environment and to conditions encountered during mammalian infection (7–11).
The Pal/Rim pathway is a fungal-specific signaling cascade that is activated in response to alkaline ambient pH (12). This pathway allows fungi to adapt to elevated pH levels as well as in the presence of high cation concentrations, such as sodium and potassium ions encountered in alkaline soil (12). Activation of this pathway during infection is also required for proper organization of cell wall components to allow for evasion of recognition by host Toll-like and C-type lectin receptors, making the Rim signaling relevant to the pathogenesis of fungal pathogens of animals (13, 14). The Pal/Rim pathway includes seven-transmembrane sensor proteins that, upon activation, initiate a cascading series of protein interactions that eventually result in the assembly of a multiprotein proteolysis complex (reviewed in reference 15). The proteolysis complex binds and cleaves the terminal transcription factor in this signaling cascade: Rim101 in yeast-like fungi and PacC in hyphal fungi. Cleavage most likely occurs at the PEST site around 100 residues from the C-terminal end (16, 17). Once cleaved, this transcriptional regulator re-localizes to the nucleus where it controls the expression of genes that favor growth at elevated pH, such as the CIG1 gene expressing a mannoprotein involved in iron chelation (18).
Many of the components required for the Pal/Rim pathway signal propagation are highly conserved among fungi. These include the Rim101/PacC transcription factor, the members of the proteolysis complex, and trafficking proteins in the endosomal sorting complexes required for transport (ESCRT). However, the proteins involved in pH sensing at the cell surface have diverged between fungal phyla. The pH-sensing Rim21/PalH proteins are highly conserved among ascomycetes, including model yeasts such as Saccharomyces cerevisiae (Sc) and model molds such as Aspergillus nidulans (An) (19, 20). These pH sensors contain seven predicted transmembrane domains anchoring them within membranes, as well as extended C-terminal cytoplasmic tails that likely serve to interact with downstream effectors. Interestingly, genes encoding sequence homologs of Rim21/PalH are not present in the genomes of fungi in the phylum Basidiomycota, a large and heterogeneous group of fungi that includes mushrooms as well as plant and animal pathogens. In contrast, basidiomycetes use a structurally similar but sequence-divergent functional ortholog as a surface pH sensor, represented by the Rra1 proteins in Cryptococcus neoformans (Cn) (21) and Malassezia sympodialis (22).
In ascomycetes, proteins linking the surface pH sensors with downstream Rim/Pal pathway components include the Rim8/PalF α-arrestin protein (17, 23). The α-arrestin proteins are often ubiquitinated by the Rsp5 ubiquitin ligase and serve as adaptors to assist in interactions between Rsp5 and other ubiquitination targets (24). The Rim8/PalF α-arrestin is mono-ubiquitinated by the Rsp5 E3 ubiquitin ligase, a modification required for interaction with downstream components of the Rim pathway (25). Ubiquitination of the Sc Rim8 and An PalF arrestin proteins facilitates interaction with the Vps23/ESCRT-I protein, eventually resulting in the assembly of the Rim20/PalA and Rim13/PalB protein complex required for cleavage and activation of the Rim101/PacC transcription factor (26, 27).
We recently characterized the role of Rsp5-mediated ubiquitination in the metabolism and pathogenesis of the opportunistic fungal pathogen C. neoformans (28). This fungus is responsible for around 120,000 deaths per year, predominantly in immunocompromised individuals, such as those living with HIV/AIDS or receiving chronic immunosuppressive therapy (29). The C. neoformans rsp5Δ loss-of-function mutant strain has pleiomorphic stress-sensitive phenotypes, including defects in thermotolerance as well as increased susceptibility to high salt and high pH (28). These latter two phenotypes are shared with strains with mutations in the C. neoformans Rim alkaline response pathway. We therefore hypothesized that Rsp5 might assist in Rim pathway activation in this human fungal pathogen, as does its ortholog in the model yeast S. cerevisiae, despite the apparent basidiomycete-wide loss of the Rim8/PalF arrestin, a target of the Rsp5 ubiquitin ligase in this pathway. We therefore explore the role of C. neoformans Rsp5 in known fungal alkaline response pathways, and we characterize a new Rsp5 target protein that mediates Rim pathway activation, growth at elevated pH, and microbe-host interactions.
RESULTS
The Cn Rsp5 E3 Ub ligase is required for activation of the Rim alkaline pH response pathway
In the model yeast, S. cerevisiae, the Rsp5 E3 ubiquitin ligase is required for activation of the Rim alkaline response pathway. One target of this ubiquitin ligase is the Rim8 α-arrestin, for which ubiquitin assists in binding to the ESCRT-I protein Vps23 (25). However, a close homolog of the Sc Rim8 protein is absent in C. neoformans (Fig. 1A) (21). To determine if the Rsp5 ubiquitin ligase enables activation of this pathway in C. neoformans, we incubated the Cn rsp5Δ mutant strain in multiple stress conditions to determine if this strain mimics the growth phenotypes of Cn Rim pathway mutant strains. Similar to the Cn rim101Δ mutant strain, the rsp5Δ mutant displayed reduced growth on alkaline media (pH 8.15) and on medium containing 1.5 M NaCl (Fig. 1B). Although activation of the Cn Rim pathway starts at pH 6, utilizing medium buffered to a pH of 8.15 allows for optimal distinction of strains with growth defects in alkaline conditions in laboratory settings (21). The rsp5Δ mutant strain also demonstrated defects in other stress-responsive phenotypes that are not shared by Rim pathway mutants, including reduced growth at 37°C and in the presence of 0.03% SDS (28). We hypothesized that the phenotypes of the rsp5Δ mutant represent a composite effect of defective ubiquitination of multiple target proteins, including those involved in Rim pathway activation. We therefore tested for specific evidence of ineffective Rim pathway activation in the rsp5Δ mutant strain.
*The Rsp5 E3 ubiquitin ligase is required for activation of the Cryptococcus neoformans Rim pathway. (A) Model of known Cn Rim pathway components, including the Rim pathway activation complex (Rra1 surface pH sensor, Nap1 scaffold), the ESCRT-mediated vesicular trafficking complex, and the Rim101 proteolysis complex (Rim23, Rim20, and the Rim13 protease). Unlike the Rim8 arrestin-like protein present in ascomycetes, no known protein has been identified to link the upstream pH sensing components with the downstream pathway effectors in C. neoformans. (B) Growth phenotypes on acidic, alkaline, and high salt stress plates. Indicated strains were incubated as serial spot dilutions for 72 hours at 30°C on yeast extract-peptone-dextrose (YPD) medium or on YPD medium buffered to pH 4, pH 8.15, or supplemented with 1.5 M NaCl. (C) Localization of a green fluorescent protein (GFP)-tagged Rim101 protein was assessed by epifluorescent microscopy in the WT strain or the rsp5Δ mutant strain expressing GFP-Rim101 after 1 hour of incubation in YPD medium at pH 8.15. The location of the nucleus was assessed by co-staining with Hoechst nucleic acid stain. Protein localization was analyzed via microscopy (GFP channel), and images were analyzed via ImageJ/Fiji. Representative differential interference microscopy (DIC), GFP (GFP-Rim101), and DAPI (Hoechst, nuclear) images are shown. (D) Alkaline pH-mediated proteolytic processing of the GFP-tagged Rim101 transcription factor. The WT and rsp5Δ mutant strains expressing GFP-RIM101 were incubated in YPD medium buffered to either pH 4 (Rim pathway non-activating conditions) or pH 8.15 (Rim pathway activating conditions) for 1 hour at 30°C prior to trichloroacetic acid (TCA)-based protein extraction. Proteolytic cleavage of GFP-Rim101 was analyzed via western blotting using an anti-GFP antibody (α-GFP), and even protein loading and transfer were assessed via Ponceau S staining. (E) Transcript abundance of the Rim101 target gene CIG1. Strains were incubated in YPD medium buffered to pH 4 and pH 8.15 for 1 hour at 30°C, and total RNA was harvested. Transcript abundance was assessed by quantitative real-time PCR (qRT-PCR), and the log2 fold change relative to WT was calculated using the ΔΔ CT method. Transcript levels were normalized to GPD1 transcript levels. The mean with error bars indicating the standard error of the mean of three biological replicates is plotted. Statistical analysis was performed using one-way analysis of variance (ANOVA) and an appropriate ad hoc test (***P < 0.00001). (F) Growth phenotypes of a rsp5Δ mutant strain expressing a constitutively active form of Rim101 compared to the rsp5Δ mutant strain alone. The indicated strains were incubated as serial spot dilutions for 72 hours at 30°C on YPD medium or on YPD medium buffered to pH 8.15, as well as on medium containing galactose to induce expression of the constitutively active Rim101 protein.
As previously demonstrated, a green fluorescent protein (GFP)-tagged Rim101 transcription factor re-localizes from the cytoplasm to the nucleus when activated by alkaline pH (21). We therefore used fluorescent microscopy to determine whether GFP-tagged Rim101 demonstrated pH-dependent changes in cellular localization in the rsp5Δ mutant. In contrast to wild type (WT), GFP-Rim101 did not demonstrate nuclear enrichment in the rsp5Δ strain background when the fungal cells were transferred to yeast extract-peptone-dextrose (YPD) medium, pH 8.15, for 1 hour (Fig. 1C). Since Rim101 nuclear localization is preceded by proteolytic activation, we assessed alterations in GFP-Rim101 by western blot in the WT and rsp5Δ mutant at different pH conditions. This study demonstrated alkaline pH-dependent proteolytic processing of GFP-Rim101 in the WT strain and failed proteolysis in the rsp5Δ mutant, similar to that observed in other Cn Rim pathway mutant strains (Fig. 1D) (21).
We also tested the transcriptional effects of defective Rim101 processing in the rsp5Δ mutant by measuring relative transcript levels of the CIG1 gene, a direct transcriptional target of Cn Rim101, and encoding an extracellular mannoprotein involved in iron uptake (30, 31). Transcription of CIG1 is regulated by the Rim101 transcription factor, and its expression is highly induced at alkaline pH in a Rim101-dependent manner (31). Similar to the expression measured in a rim101Δ mutant, CIG1 transcript levels were not induced under alkaline conditions in the rsp5Δ mutant strain (Fig. 1E). In contrast to the case for Rim pathway mutants, CIG1 expression was induced by alkaline pH in the sre1Δ sterol response pathway mutant, a strain with a distinct mechanism of defective alkaline pH growth (Fig. 1E) (32). Finally, we expressed a truncated form of the Rim101 transcription factor in rsp5Δ mutant cells lacking 603 amino acid residues at the C-terminal end (21). This truncated form is constitutively active and is able to rescue the growth defect of Rim pathway mutants on alkaline medium, which was observed in the rsp5Δ mutant strain expressing truncated Rim101 as well (Fig. 1F). Together, these data demonstrate that Cn Rsp5 is required for Rim pathway activation, as assessed by pH-mediated Rim101 proteolytic cleavage and nuclear localization, as well as phenotypic rescue of growth defects on alkaline medium with the expression of constitutively active Rim101 and through assessing the transcriptional regulation of a Rim pathway target gene.
Cn arrestin domain-containing proteins and growth in alkaline pH
In S. cerevisiae, Rsp5 mediates Rim pathway activation through its ubiquitinating activity of the Rim8 arrestin-like protein (25). We previously reported that two Cn proteins containing both N-terminal and C-terminal arrestin domains (arrestin-like proteins Ali1 and Ali2) are not individually required for growth at pH 8.15, and therefore are not likely required for Rim pathway activation (21). We subsequently identified two additional proteins with a single arrestin domain, Ali3 and Ali4 (28, 33). Ali1, Ali2, Ali3, and Ali4 are the only proteins in the Cn genome with classical arrestin domains predicted by simple sequence analysis (33). None of the individual aliΔ mutant strains is required for growth at alkaline pH. To rule out functional redundancy among the arrestins, we tested whether a strain lacking all four of the known Cn arrestin proteins is able to grow in alkaline pH conditions. The ali1Δ ali2Δ ali3Δ ali4Δ quadruple arrestin mutant also grew similarly to WT at pH 8.15 (Fig. S1), indicating that these four arrestin-like proteins are not essential, either individually or in combination, for Cn Rim pathway activation and alkaline pH growth.
Rsp5 does not ubiquitinate known members of the Cn Rim pathway
Since no Cn proteins with clearly identifiable arrestin domains were required for Rim pathway-related phenotypes, we hypothesized that Rsp5 might mediate Rim pathway activation through a non-canonical adaptor protein. Rsp5 adaptors in other eukaryotes are often ubiquitinated by this E3 Ub ligase as a means for their activation (34, 35). We therefore used a mass spectroscopy-based proteomic method to identify C. neoformans proteins that are likely ubiquitinated by Cn Rsp5 in a pH-dependent manner. We previously used this experimental protocol to identify Cn Rsp5 targets ubiquitinated under high salt conditions (28). We incubated the WT and rsp5Δ mutant strains in YPD medium at pH 4 and pH 8.15. Total cell lysates from each condition were digested with trypsin and enriched for ubiquitinated peptides using an antibody recognizing the characteristic “ubiquitin stump” remaining after trypsin treatment (36). Mass spectroscopy of the resulting samples identified 96 proteins that were preferentially ubiquitinated in the WT strain compared to the rsp5Δ mutant strain at pH 8.15 and 48 preferentially ubiquitinated in the WT strain compared to the rsp5Δ mutant strain at pH 4, while 34 proteins were preferentially ubiquitinated in the WT strain at both pH 4 and pH 8.15 (Fig. 2A through D; Tables S1 and S2). In WT cells, 16 proteins were more abundant in ubiquitinated form at pH 4 than at pH 8.15, and 25 were more abundant at pH 8.15 than at pH 4, although Ub ligases other than Rsp5 might be responsible for their ubiquitination (Fig. 2C; Tables S3 and S4). No known Rim pathway protein was significantly enriched in this group of proteins, although the Rim101 transcription factor did appear in this set as a ubiquitinated protein, consistent with previous findings in S. cerevisiae (Fig. 2A through C) (37).
Identification of proteins requiring Rsp5 for ubiquitination at pH 4 and pH 8.15. The WT and rsp5Δ strains were incubated in either YPD medium at pH 4 or YPD medium buffered to pH 8.15 for 1 hour at 30°C. Total cell lysates were treated with trypsin, resulting in a ubiquitin remnant stump attached to ubiquitinated protein fragments. These previously ubiquitinated proteins were enriched by immunoaffinity and identified by quantitative liquid chromatography-tandem mass spectrometry. (A and B) Volcano plots of individual peptides identified by mass spectrometry demonstrate relative peptide abundance (log2 fold change) in the rsp5Δ strain compared to WT at either pH 4 (A) or pH 8.15 (B). (C) A volcano plot of individual peptides identified by mass spectroscopy from WT cells with abundance at either pH 4 or at pH 8.15. The identified peptides belonging to the CNAG_05520 and Rim101 proteins are indicated. (D) A Venn diagram indicating protein amounts requiring Rsp5 for ubiquitination when exposed to acidic pH or alkaline pH, or that are insensitive to pH. (E) A growth phenotype screen using a Cryptococcus neoformans mutant strains collection (38) to screen for rim101Δ mutant-like stress phenotypes (decreased growth in alkaline pH and high salt media) in mutant strains corresponding to the proteins identified to be ubiquitinated at pH 4 (left) and pH 8.15 (right) in an Rsp5-dependent manner. Black boxes indicate no growth, light green boxes indicate some growth, and dark green boxes indicate WT-like growth. The CNAG_05520Δ mutant strain, indicated by an asterisk, was the only strain with decreased CIG1 expression, as is also seen in a rim101Δ mutant strain.
To determine if any of the 144 proteins that require Rsp5 for their ubiquitination at either pH 4 or pH 8.15 are required for Rim pathway activation, we made use of an established collection of strains with mutations in non-essential Cn genes to screen for Rim pathway-related phenotypes: impaired growth at pH 4 and pH 8.15 was compared for each mutant as well as impaired growth on 1.5 M NaCl (39). Of the 144 differentially ubiquitinated proteins, 76 had a corresponding mutant strain in this collection at the time of this analysis, and more mutants might become available as mutant strain collections are expanded. Three of these strains demonstrated defects in both of these Rim pathway-related stress conditions. However, only one mutant strain, lacking the uncharacterized CNAG_05520 gene, also demonstrated reduced CIG1 expression under alkaline conditions similar to Rim pathway mutants (Fig. 3D).
*The CNAG_05520 gene is required for C. neoformans Rim pathway activation. (A) Growth phenotype on alkaline and high salt stress plates. Indicated strains were incubated as serial spot dilutions for 72 hours at 30°C on YPD medium or on YPD medium either buffered to pH 8.15 or supplemented with 1.5 M NaCl. (B) Localization of a GFP-tagged Rim101 protein was assessed by epifluorescent microscopy in the WT strain or the CNAG_05520Δ mutant expressing GFP-Rim101 after 1 hour of incubation in YPD medium at pH 8.15. The location of the nucleus was assessed by co-staining with Hoechst nucleic acid stain. Protein localization was analyzed via microscopy (GFP channel), and images were analyzed via ImageJ/Fiji. Representative DIC, GFP (GFP-Rim101), and DAPI (Hoechst, nuclear) images are shown. (C) Alkaline pH-mediated proteolytic processing of the GFP-tagged Rim101 transcription factor. The WT, rsp5Δ mutant, and CNAG_05520Δ mutant strains expressing GFP-RIM101 were incubated in YPD medium buffered to either pH 4 (Rim pathway non-activating conditions) or pH 8.15 (Rim pathway activating conditions) for 1 hour at 30°C prior to TCA-based protein extraction. Proteolytic cleavage of GFP-Rim101 was analyzed via western blotting using an anti-GFP antibody (α-GFP), and even protein loading and transfer were assessed via Ponceau staining. (D) Transcript abundance of the Rim101 target gene CIG1. Indicated strains were incubated in YPD medium, pH 4 or pH 8.15, for 1 hour at 30°C, and total RNA was harvested. Transcript abundance was assessed by qRT-PCR, and the log2 fold change relative to WT was calculated using the ΔΔ CT method. Transcript levels were normalized to GPD1 transcript levels. The mean with error bars indicating the standard error of the mean of three biological replicates is plotted. Statistical analysis was performed using one-way ANOVA and an appropriate ad hoc test (*P < 0.001).
The CNAG_05520 gene product is required for activation of the Cn Rim101 transcription factor
To determine if the CNAG_05520Δ mutant strain, like the rsp5Δ mutant strain, is required for Rim101 pathway activation, we assessed alkaline pH and high salt stress growth, Rim101 cleavage and nuclear localization as well as Rim pathway target gene expression in the CNAG_05520Δ mutant strain (Fig. 3). Similar to the rsp5Δ ubiquitin ligase mutant strain, the GFP-Rim101 protein fails to re-localize in the nucleus in the absence of the CNAG_05520 gene when cells are incubated in high pH medium (Fig. 3B). Additionally, GFP-Rim101 remains in an uncleaved form in the CNAG_00520Δ mutant strain at high pH (Fig. 3C). The functional relevance of failed activation of Rim101 was confirmed by ineffective pH-mediated induction of the Rim101 target gene CIG1 as determined by quantitative real-time PCR (Fig. 3D).
Absence of the CNAG_05520 gene product displays similar pathogenesis-related phenotypes as the rim101Δ mutant strain
We previously established that the Cn Rim pathway is required for efficient capsule production as well as titan cell formation (40). To determine if the CNAG_05520 gene product might play a similar role in these pathogenesis-associated microbial phenotypes, we cultivated the WT strain, the rim101Δ and CNAG_05520Δ mutant, and a CNAG_05520Δ mutant complemented strain in capsule and titan cell-inducing conditions. We observed that cells lacking the CNAG_05520 gene displayed similar capsule and titan cell formation defects as the rim101Δ mutant (Fig. 4A and B). Furthermore, like the rim101Δ mutant strain, the CNAG_05520Δ mutant strain displayed increased chitin exposure, as assessed by staining with Alexa Fluor 488-conjugated wheat germ agglutinin (WGA-Alexa F488) (Fig. 4C) (41).
*A CNAG_05520Δ mutant mimics the rim101Δ mutant virulence traits. (A) Capsule formation. Indicated strains were grown at capsule-inducing conditions (CO2-independent medium at 37°C for 3 days). Capsule formation was analyzed via India ink contrast staining and microscopy. Images were analyzed using ImageJ/Fiji. Representative DIC images are shown. (B) Titan cell formation. Indicated strains were grown overnight in YNB medium at 30°C and then transferred to in vitro titan cell-inducing conditions (OD600 of 0.001 in PBS + 10% HI-FBS) and then incubated for 24 hours at 37°C with 5% CO2. Cells were harvested and cell size assessed via microscopy. Images were analyzed using ImageJ/Fiji. Representative DIC images are shown. (C) Total chitin staining and exposure of cells grown under host-mimicking stress. Indicated strains were grown overnight in YPD medium and then transferred to CO2-independent medium (at an OD600 of 0.2) and grown for around 18 hours at 37°C. Cells were harvested and stained with calcofluor white (CFW—binds total cell wall chitin, blue channel) and Alexa Fluor 488 conjugated wheat germ agglutinin (WGA-Alexa F488—binds exposed cell wall chitin, GFP channel). Staining pattern and intensity were assessed via microscopy, and images were analyzed via ImageJ/Fiji. Representative DIC, blue channel (CFW staining), and GFP channel (WGA-Alexa F488) images are shown. (D) Lung fungal burden 7 days post-infection. Female A/J mice (5 mice per strain) were infected intranasally with 105 cells/mouse of the indicated C. neoformans strains. Mice were sacrificed 7 days after infection, lungs harvested, and the fungal burden was assessed by quantitative culture. Data were plotted, and a Mann-Whitney U-test was done to assess statistical significance. (E) Histopathology analysis of infected murine lungs 7 days post-infection. Mice were infected as described above. Mice were sacrificed 7 days after infection, and lungs were fixed via intratracheal instillation with neutral buffered formalin under gravity flow. For visualization of lung pathology, lung sections were stained with hematoxylin and eosin. Red arrows show encapsulated yeast inside the inflamed lung tissue. (F) Tumor necrosis factor-α (TNF-α) was produced by murine bone marrow macrophages upon co-culturing with the indicated strains. Macrophages and fungal cells were co-cultured for 6 hours, and the supernatant was harvested. The secreted TNF-α was quantified with an enzyme-linked immunosorbent assay (ELISA). Student’s t-test was done to evaluate statistical significance (P < 0.05).
Due to this altered cell wall polysaccharide exposure, Rim pathway mutants induce an excessive immune response in the infected lungs, resulting in failed immune evasion and early host death due to immune-mediated organ damage (40). To further assess phenotypic similarities between the CNAG_05520Δ mutant and the rim101Δ mutant, we assessed the degree of lung inflammation by histopathology analysis after an inhalational infection with either the WT, CNAG_05520Δ mutant, or CNAG_05520 complemented strain at 7 days post-infection (Fig. 4D and E). A/J mice infected with the WT strain demonstrated an expected pattern of diffuse, minimal inflammatory infiltration of the infected lungs due to the potential for this microorganism to partially evade initial immune recognition. In contrast, infection with the CNAG_05520Δ mutant leads to a hyperinflammatory response in the lungs at 7 days post-infection, characterized by early and increased lung infiltration by neutrophils, despite similar fungal burdens (Fig. 4D and E). This observation is similar to what was previously observed in mice infected with a Cn rim101Δ mutant strain (21, 40). Additionally, we assessed production of the pro-inflammatory cytokine tumor necrosis factor-α (TNF-α) in bone marrow-derived macrophages harvested from CD1 mice in response to co-culturing with WT, CNAG_05520Δ mutant, rim101Δ mutant, or CNAG_05520 complemented cells. In agreement with our histopathology results as well as previously published observations, more TNF-α was produced in response to co-culturing with the CNAG_05520Δ mutant and rim101Δ mutant cells compared to WT and CNAG_05520 complemented cells (Fig. 4F) (40). These results indicate that defective capsule formation and altered cell wall organization due to defective Rim signaling result in increased exposure of epitopes recognized by host immune cells.
The uncharacterized CNAG_05520 gene product has similar functional motifs as the ascomycete PalF/Rim8 protein
The CNAG_05520 gene encodes an uncharacterized protein with no clear homologs in ascomycete fungi. However, similar to the Cn Rra1 pH sensing protein, it shares a high degree of sequence similarity with genes present in the genomes of more closely related basidiomycete fungi when aligned with fungal genomes (Fig. S2). Unlike Rim8/PalF, the CNAG_05520 predicted protein does not have identifiable arrestin domains but two predicted membrane-spanning regions and several short motifs, which are homologous to functional motifs observed in Rim8/PalF proteins (25, 42). The CNAG_05520 protein contains four PxY motifs that typically bind the Rsp5 E3 ubiquitin ligase, similar to the three PxY motifs present in Sc Rim8 (25). Additionally, the CNAG_05520 protein contains a single lysine residue adjacent to two negatively charged amino acids in a C-terminal tail region, similar to the site of Rsp5-mediated ubiquitination in Sc Rim8. Notably, this lysine residue is predicted to be ubiquitinated by Cn Rsp5 by our above-described differential ubiquitination screen. Furthermore, the CNAG_05520 gene product shares with Sc Rim8 a SxP motif 10-12 amino acids downstream from the predicted ubiquitinated lysine residue (42). This SxP motif, together with ubiquitin, is known as the box motif and is predicted to interact with the ubiquitin E2 variant (UEV) domain of an ESCRT-I protein (Fig. 5A). Computational modeling using the protein structure prediction artificial intelligence program, AlphaFold v. 3, predicts that the CNAG_05520 protein SxP motif is indeed in close proximity with the UEV domain of the Cn Rim pathway ESCRT-I scaffold protein Vps23 when interaction between these two proteins is modeled through the multimer mode of AlphaFold V. 3 (Fig. 5B). Furthermore, a CNAG_05520 mutant protein, with mutations of both the ubiquitinated lysine residue and the SxP motif, failed to functionally complement the CNAG_05520Δ mutant growth defects in the presence of high pH or 1.5 M NaCl (Fig. 5C). In contrast, in mutant strains expressing CNAG_05520 protein in which the ubiquitinated lysine residue or the SxP motif alone was mutated to arginine residues, the mutant strain growth defect is partially rescued in the presence of high pH or 1.5 M NaCl. These observations suggest functional relevance in CNAG_05520 of the predicted Vps23-interacting regions.
CNAG_05520 has similar protein features to the ascomycete Rim8 arrestin-like protein. (A) Model of predicted functional domains of the Cn CNAG_05520 protein, including two hydrophobic membrane-spanning domains and residues in the C-terminus similar to the Vps23-binding “box domain” of the Rim8/PalF proteins in S. cerevisiae (Sc) or A. nidulans (An). (B) AlphaFold model of potential interaction between Cn Vps23 and Cn CNAG_05520. The SxP motif is predicted to reside at the site of interaction. Distances of 5 Å or less are indicated with green dashed lines. (C) Growth phenotypes of strains with mutated CNAG_05520 lacking the SxP motif and ubiquitination site on alkaline and high salt stress-inducing plates. Indicated strains were incubated as serial spot dilutions for 72 hours at 30°C on YPD medium or on YPD medium either buffered to pH 8.15 or supplemented with 1.5 M NaCl.
The CNAG_05520 gene product interacts with Cn Vps23
The Sc Rim8 arrestin protein interacts with Vps23, an early component of the ESCRT complex that facilitates recruitment and assembly of downstream Rim pathway proteins (25, 42). We, therefore, tested whether the CNAG_05520 protein similarly interacts with the Cn Vps23 protein. This interaction is predicted based on shared UEV-interaction motifs, present in both the CNAG_05520 protein and PalF/Rim8 proteins, that have been previously demonstrated to promote this protein-protein interaction (25, 42).
These potential intermolecular interactions were further explored by assessing patterns of cellular localization of a CNAG_05520-GFP fluorescent fusion protein and a Vps23-mCherry fluorescent fusion protein using confocal microscopy. The CNAG_05520-GFP protein demonstrates prominent perinuclear localization, with focal regions of punctate enrichment both near the nucleus and at the cell surface (Fig. 6A). In the absence of the ubiquitinated lysine residue as well as the SxP motif, the perinuclear localization is lost (Fig. 6B). These surface puncta are similar to those observed in strains expressing the Rra1-GFP protein (21, 43). The Vps23-mCherry protein is enriched in punctate regions near the nucleus, and these structures largely co-localize with CNAG_05520-GFP (Fig. 6B). To confirm binding of Cn Vps23 to the CNAG_05520 gene product, we performed a co-immunoprecipitation assay with cells expressing both a CNAG_05520-GFP fluorescent fusion protein and a Vps23-mCherry fluorescent fusion protein. We were able to detect the GFP fluorescent protein fused to the CNAG_05520 gene product by probing with an anti-GFP antibody when Vps23-mCherry was immunoprecipitated, as well as the mCherry fluorescent protein fused to Vps23 with an anti-mCherry antibody when the CNAG_05520-GFP gene product was immunoprecipitated (Fig. 6C). This co-immunoprecipitation results complement our co-localization data and, together with our protein prediction modeling with AlphaFold and mutagenesis study, supports a model of conservation of function for CNAG_05520 as a Vps23-interactor.
CNAG_05520 interacts with the Vsp23 ESCRT protein. (A) Localization of CNAG_05520-GFP at alkaline pH. The CNAG_05520Δ mutant strain expressing a GFP-tagged CNAG_05520 (CNAG_05520-GFP) was incubated for 1 hour in YPD medium buffered to pH 8.15. The location of the nucleus was assessed by staining with Hoechst nucleic acid stain. Protein localization was analyzed via confocal microscopy. Images were analyzed with ImageJ/Fiji. Representative Bright field and GFP channel images are shown. (B) Co-localization of CNAG_05520-GFP with mCherry-Vsp23 at alkaline pH. The CNAG_05520Δ and vsp23Δ mutant strains expressing a GFP-tagged CNAG_05520 and an mCherry-tagged Vsp23 (CNAG_05520-GFP + mCherry-VSP23) were incubated for 1 hour in YPD medium buffered to pH 8.15. Protein co-localization was analyzed via confocal microscopy. Images were analyzed via ImageJ/Fiji. Representative Bright field, GFP, and RFP channel images are shown with co-localization indicated with red arrows. (C) Co-immunoprecipitation of the GFP-tagged Rra2 protein and mCherry-tagged Vps23. The WT strain and the strains expressing RRA2-GFP, VPS23-mCherry, or both were incubated in YPD medium buffered to pH 8.15 for 1 hour at 30°C prior to fixing with 0.8% formaldehyde and NP-40-based protein extraction. Interaction of Rra2-GFP and Vps23-mCherry was analyzed via western blotting using an anti-GFP antibody (α-GFP) and anti-mCherry antibody (α-mCherry). Normalized cell lysates used as input for the immunoprecipitation assay were blotted in parallel and probed with anti-PSTAIR (α-PSTAIR) to assess even protein loading in addition to Ponceau staining.
The CNAG_05520 gene partially complements Sc rim8Δ mutant phenotypes
Although the Sc Rim8 protein and the Cn CNAG_05520 protein are divergent in primary sequence, they appear to serve orthologous roles in Rim pathway activation through interactions with ESCRT-I proteins. We therefore tested whether expression of Cn CNAG_05520 in S. cerevisiae could partially or completely complement Sc rim8Δ mutant phenotypes. The Cn CNAG_05520 gene was cloned into the 2µ pYES galactose-regulatable yeast expression vector and transformed into a Sc rim8Δ mutant. An intron-less complementary DNA (cDNA) version of this gene was also cloned into the same vector and transformed into the Sc rim8Δ mutant. Expression of the Cn CNAG_05520 gene in the Sc rim8Δ mutant cells was confirmed by real-time quantitative PCR (Fig. 7B). In contrast to a rim8Δ strain containing the empty pYES plasmid, the rim8Δ strain containing both the complete and the intron-less pYES-CNAG_05520 plasmid strain displayed partial growth restoration on media containing 1.5 M NaCl (stress condition) and 2% galactose (drives gene expression in the pYES vector) (Fig. 7A). No phenotypic complementation was observed on galactose-containing media at pH 7.5 (Fig. 7A), even when 0.2% glucose was added to support growth. This partial functional complementation of a Sc Rim pathway mutant-associated phenotype by Cn CNAG_05520 suggests functional convergence of CNAG_05520 and Rim8 despite sequence and evolutionary divergence.
*CNAG_05520 partially complements rim8Δ mutant growth phenotypes. (A) Cross-species complementation of the S. cerevisiae (Sc) rim8Δ high salt growth defect with C. neoformans (Cn) CNAG_05520. Indicated strains were incubated as serial spot dilutions for 72 hours at 30°C on synthetic complete medium (SC) without uracil and galactose as carbon source (SC-ura+Gal) or SC-ura+Gal medium supplemented with 1.5 M NaCl. (B) Transcript abundance of the C. neoformans (Cn) gene RRA2 expressed in Sc. Indicated strains were incubated in SC medium at pH 7.5 for 1 hour at 30°C, and total RNA was harvested. Transcript abundance was assessed by qRT-PCR, and the log2 fold change relative to Sc rim8∆ was calculated using the ΔΔ CT method. Transcript levels were normalized to GPD1 transcript levels. The mean with error bars indicating the standard error of the mean of three biological replicates is plotted. Statistical analysis was performed using one-way ANOVA and an appropriate ad hoc test (**P < 0.0001). (C) Updated model of the Cn Rim pathway. The newly discovered CNAG_05520 protein functions as a linker between the Rim pathway activation complex and the downstream ESCRT complex. Rsp5 assists in Rim pathway activation via ubiquitination of CNAG_05520.
Consistent with nomenclature for the previously identified RRA1 gene (required for Rim pathway activation-1) (21), and to acknowledge the recent and independent identification of this gene through a large screen of Cn mutants for genes required for Rim pathway activity (38), the CNAG_00520 gene was therefore named RRA2. The data presented here suggest that it serves to link Rra1 pH-sensing with downstream Rim pathway activation. Therefore, despite no identifiable sequence or structural similarities with the Sc Rim8 protein, it does share Rsp5-mediated ubiquitination and ESCRT-protein interactions that are common among Rim8/PalF proteins (Fig. 7C) (25–27).
DISCUSSION
The fungal kingdom includes a vast array of ecologically diverse species that employ several unique molecular strategies to thrive in equally diverse habitats. Studying these stress-response pathways provides insight into evolutionary biology, but it also reveals novel cell biology that can be exploited in biotechnology or targeted by pharmaceuticals for the treatment of fungal infections. In this study, we explored the fungal-specific Rim alkaline pH response pathway, allowing acidophilic fungi to adapt to a pH above acidic levels. Although several downstream Rim pathway proteins are homologous among fungi across the ascomycota and basidiomycota phyla, differences occur in the upstream sensing complex (44).
External alkalinization is sensed by the seven transmembrane protein Rim21 in Saccharomyces cerevisiae and the homologous PalH in Aspergillus nidulans (19, 45). We have previously identified a sequence divergent, but functionally analogous, sensing protein in Cryptococcus neoformans, Rra1 (21). Although very different in primary amino acid sequence, there is evidence that the Sc Rim21 and the Cn Rra1 sensing proteins are both activated by alkaline pH-associated changes in the plasma membrane lipid bilayer asymmetry (19, 46). This model of pH sensing was supported by studies of the Cn Cdc50 lipid flippase regulatory subunit that was identified as a temporal effector of Rim pathway activation (47).
Containing seven predicted membrane-spanning domains, the Sc Rim21, An PalH, and Cn Rra1 pH sensor proteins resemble G-protein coupled receptors (GPCRs), although no interaction with G-proteins is known (21, 23). These GPCR-like sensor proteins might represent a growing group of seven-transmembrane domain-containing proteins with structural homology to GPCRs, but no known G protein binding partners (reviewed by reference 48). However, like GPCRs, Sc Rim21 and An PalH interact with arrestin-domain containing proteins that facilitate binding to ubiquitinating enzymes (23). While GPCRs typically associate with visual and β-arrestins, the Rim pathway sensor proteins associate with α-arrestin proteins, a distinct class of arrestins with PxY motifs to facilitate interaction with the WW domains of HECT E3 Ub ligases (49). Ubiquitination is best known for its function in the degradation of spent or misfolded protein, but it can also help direct changes in protein localization and function (50, 51). The specific ubiquitination events important in the Sc and An Rim pathway signal propagation involve the HECT E3 Ub ligase Rsp5 and its interaction with the arrestin domain-containing protein Rim8/PalF (23, 52). These events involve the ubiquitination of the Sc Rim8 arrestin upon binding to the Rim21 sensor protein, irrespective of the external pH (25). In contrast, ubiquitination of An PalF depends on sensor binding as well as an alkaline environment (23).
There are no proteins encoded in the Cn genome with a high degree of sequence similarity to Sc Rim8 or An PalF. We have previously identified four arrestin domain-containing proteins in Cn (Ali1, Ali2, Ali3, and Ali4), and subsequent phenotypic analysis of single and quadruple mutant strains suggests that none of these arrestin-like proteins are required for Cn Rim pathway activation.
Arrestin-mediated ubiquitination of the Rim8/PalF proteins by Rsp5 does not result in its accelerated protein degradation, but rather in the addition of a ubiquitin moiety, which likely serves as an interface for binding of the UEV domain of the ESCRT-I protein Vps23 (42). Proteins of the ESCRT complex are typically involved in the sorting of proteins to lysosomes or the vacuole for degradation (reviewed in reference 53). However, in this instance, the ESCRT complex has an established role in the fungal Rim/Pal pathway to recruit the Rim13/PalB protease to activate the Rim101/PacC transcription factor and to potentially internalize the sensor proteins for their regulated degradation. In addition to protein sorting to the vacuole/lysosomes, the only other known function of Vps23 is to serve as a linker between the Rim8/PalF proteins and the ESCRT-II protein, a role for which it was likely co-opted (52).
We have previously demonstrated that ESCRT proteins are also required for Cn Rim pathway activation (21). However, the steps linking surface pH recognition and ESCRT assembly have not been previously identified in this pathogenic fungus. Our data have identified Rra2 as a target of Rsp5 and as a protein required for Cn Rim pathway activation. Although Rra2 lacks distinct sequence similarity to Rim8, it shares similar protein features, including a ubiquitinated lysine residue 11 to 13 amino acid residues upstream of an SxP motif in the C-terminal region of these proteins. We also demonstrate its co-localization and co-immunoprecipitation with Vps23, strongly supporting that it serves as an Rsp5 adapter protein for Rim pathway activation. Molecular modeling of potential structural interactions between Cn Vps23 suggests similar sites of interaction, as has been experimentally verified between Sc Rim8 and Sc Vps23 (25). A recent study conducted by Madhani et al. suggests functional interaction between the membrane pH sensor Rra1 and the newly identified Rim1-pathway component Rra2 (38). Based on their findings as well as our findings here, we therefore suggest that Rra2 acts as a linking component between the Rra1 pH membrane sensor and the Rim101-activating ESCRT complex. Our data further suggest that, similar to Sc Rsp5, Cn Rsp5 regulates Rim pathway activation by ubiquitination of the linker protein Rra2 (Fig. 7A).
Our data presented here demonstrate that Rra2, like other Rim pathway proteins, is essential for Rim pathway-dependent adaptation of this fungal pathogen to the alkaline pH environment within the infected host. These adaptations include host-mediated cell surface changes that provide efficient capsule attachment and immune avoidance. In the absence of Rra2, Cn fails to maintain its surface capsule and does not make titan cells in host-like in vitro growth conditions. Additionally, the rra2Δ mutant induces an altered pattern of early inflammatory responses in the infected lung. Similar patterns of excessive early inflammatory responses are observed in other Cn Rim pathway mutants, resulting in immune-mediated pathology and accelerated mortality (40). Together, these data demonstrate a central role for this non-traditional Ub ligase adaptor in Cn Rim signaling.
The Cn Rsp5 E3 Ub ligase regulates several stress response processes in addition to adaptation to alkaline pH (28). Moreover, this ubiquitinating enzyme uses both arrestin-like proteins as well as proteins with no discernible arrestin domains as adaptors to bind its target proteins (28, 33). There is considerable interest in ubiquitination as a therapeutic target for many conditions. These include many cancers and infections by eukaryotic pathogens (reviewed in references 54, 55). Importantly, many of the proteins involved in the Rim pathway interactions are either only found in fungi or contain fungal-specific interacting domains (56). Our results here, therefore, suggest that fungal-specific inhibitors of Rsp5-mediated ubiquitination might be harnessed to modulate microbial survival in vivo.
MATERIALS AND METHODS
Strains, media, and growth conditions
A list of the strains used in this study is given in Table S5, including details of strains obtained from Dr. Hiten Madhani (39). The C. neoformans var. grubii H99 (MATα) strain background was used as wild type (WT) for this study unless otherwise specified (57). All strains were stored in glycerol stocks at −80°C and were recovered on YPD medium (1% yeast extract, 2% peptone, 2% dextrose, and 2% agar for solid medium) before each experiment. Unless otherwise indicated, strains were incubated in YPD medium at 30°C with shaking at 150 rpm. Complemented mutant strains and fluorescent-tagged strains were constructed using CRISPR-Cas9 gene editing techniques as previously described (28). These constructs were integrated into the Cn genome at the Safe Haven site following targeting with a single guide RNA oligonucleotide with a GGTCTGATCAGTTATAGACG binding site (58). To obtain strains of the opposite mating type, the MATα rra2Δ mutant strain (MDP80) and the MATα *VPS23-*mCherry expressing strain (KS272) were crossed with the KN99a strain to obtain MATa strains (MDP87 and MDP93, respectively), following the approach described earlier (28). The MATa rra2Δ (MDP87) and MATa rsp5Δ (MDP11) mutant strains were crossed with the KS208 strain expressing GFP-tagged RIM101, yielding MDP81 (rsp5Δ expressing GFP-RIM101) and MDP82 (rra2Δ expressing GFP-RIM101). Similarly, the MATa rsp5Δ (MDP11) mutant strain was crossed with the KS161 strain expressing the constitutively active truncated RIM101 gene under transcriptional control of the GAL7 promoter to obtain strain MDP47, an rsp5∆ strain expressing truncated RIM101. Primer pair AA5778 and AA5779 were used to amplify the CNAG_05520 open reading frame (ORF) as well as 600 bp upstream and downstream for integration with the pSDMA57 plasmid to create the rra2Δ+RRA2 complemented strain (58). The plasmid was linearized with primer pair AA5780 and AA5781 with overlaps to the CNAG_05520 fragment for assembly with the NEBuilder HiFi DNA Assembly kit (NEB), as previously described (28). In a similar manner, the primer pair AA5754 and AA5755 were used to amplify the CNAG_05520 ORF, and primer pair AA5752 and AA5753 the pGWKS10 plasmid containing the clover (GFP) fluorescent gene for assembly with the NEBuilder HiFi DNA Assembly kit to create the GFP-tagged RRA2 expressing strain (59).
Overlapping primer pair AA5782 and AA5783 was used to introduce the K539R mutation into the CNAG_05520 ORF in the pGWKS10 plasmid. The SAP550-552AAA mutation was introduced using the primer pair AA5817 and AA5755 to create a megaprimer that was used in a subsequent PCR to amplify the whole CNAG_05520 ORF for assembly into the pGWKS10 plasmid, an approach described earlier (60). The GFP-tagged RRA2-expressing strains were crossed with the MDP93 VPS23-mCherry-expressing strain to obtain strains with both tagged genes.
The pYES2.1 plasmid containing the WT Sc RIM8 gene was constructed using the primer pair AA5862 and AA5863 to amplify the RIM8 gene from Sc BY4741 genomic DNA. The primer pair AA5864 and AA5865 was used to amplify the pYES2.1 plasmid with overlapping regions to the RIM8 gene for assembly with the NEBuilder HiFi DNA Assembly kit. The primer pair AA5858 and AA5859 was used to amplify the CNAG_05520 ORF from Cn WT genomic DNA (with introns) or cDNA (without introns), and primer pair AA5860 and AA5861 was used to amplify the pYES2.1 plasmid with overlapping regions to the CNAG_05520 ORF. These two fragments were assembled as before using the NEBuilder HiFi DNA Assembly kit. The Sc pYES2.1 plasmids were transformed into Sc rim8Δ mutant cells with the lithium acetate approach described in the pYES2.1 TOPO TA Yeast Expression Kit protocol (Thermo Fisher Scientific). Strains were verified with diagnostic PCR and Sanger sequencing. Oligonucleotide primers used for strain validation and construction are listed in Table S6.
Capsule induction was performed by incubating WT, rra2Δ mutant, rim101Δ mutant, and cap59Δ mutant (acapsular control strain) cells in CO_2_-independent medium (CIM; Gibco) at 37°C while shaking for 72 hours and then staining with India ink before microscopy imaging (Zeiss Axio Imager A1) (28).
To induce titanization, cells were precultured for 18 hours in YNB at 30°C. Cells were then washed 1× with phosphate-buffered saline (PBS) and transferred to 10 mL PBS + 10% heat-inactivated fetal bovine serum (HI-FBS) in 6-well tissue culture plates. For low-density growth (titan induction), cells were inoculated to an OD_600_ of 0.001 and incubated for 72 hours in a tissue culture incubator at 37°C with 5% CO_2_. Cells were then imaged with microscopy as before (61).
To assess cell wall integrity through the extent of chitin exposure, WT cryptococcal cells, rra2Δ mutant, rim101Δ mutant, as well as rra2Δ+RRA2 complemented cells, were grown for 18 hours at 30°C in YPD medium before being shifted to fresh YPD medium for 7 hours at 30°C. Cells were harvested, standardized to an OD_600_ of 0.25, resuspended in CO_2_-independent medium, and incubated at 37°C for 18 hours. Cells were harvested and stained with 100 µg/mL Alexa F488-conjugated wheat germ agglutinin (WGA; Molecular Probes) for 35 minutes, followed by 25 µg/mL calcofluor white (CFW; Fluka Analytical) for 10 minutes, and finally imaged with microscopy, as described before (62).
Survival in the presence of stress requiring intact Rim pathway activation was assessed through a spot-dilution assay on supplemented YPD medium as previously described (21). Fungal cells were serially diluted and spotted onto solid media plates and exposed to alkaline pH (disrupts electrochemical gradient) and sodium chloride (NaCl; provides osmotic/ionic stress). To assess S. cerevisiae rim8Δ mutant cell growth—as well as the growth of rim8Δ mutant cells complemented with intact RIM8, RRA2 including introns, or RRA2 without introns—on stress-inducing plates, synthetic complete (SC) medium was used, lacking uracil and with 2% galactose as carbon source. In alkaline solid medium plates with 2% galactose, 0.2% glucose was added to support the initial growth of complemented mutant strains.
Real-time quantitative PCR
To quantify the expression levels of the CIG1 gene, under the transcriptional regulation of the Rim101 transcription factor, WT, rsp5Δ mutant, sre1Δ mutant, rim101Δ mutant, and rra2Δ mutant cells were grown for 18 hours at 30°C in YPD medium. Cells were harvested and resuspended in YPD medium buffered to pH 8.15 and incubated for 1 hour at 30°C. Cells were collected and RNA was extracted following the protocol of the Qiagen RNeasy Plant Minikit (Qiagen). cDNA was prepared by reverse-transcriptase PCR using the AffinityScript cDNA qPCR Synthesis kit (Agilent Technologies), following the included protocol. Real-time quantitative PCR was performed as described before, using the described primers for CIG1 cDNA amplification (47). Similarly, Sc rim8Δ mutant cells containing an empty pYES 2.1 vector, a pYES 2.1 vector containing WT RIM8, or Cn RRA2 with or without introns were grown for 18 hours at 30°C in SC medium without uracil and with 2% galactose. Cells were harvested and resuspended in SC medium buffered to pH 7.5 and incubated for 1 hour at 30°C. Cells were collected and RNA was extracted as before. Complementary DNA was prepared as before, and real-time quantitative PCR was performed as described before, using primer pair AA5924 and AA5925 for RRA2 cDNA amplification.
Histology and persistence in a murine model
We used the murine inhalation model of cryptococcosis (28) to assess fungal survival and host response after lung infection with WT cryptococcal cells, as well as with rra2Δ mutant and rra2Δ+RRA2 complemented cells. For each strain, five female A/J mice were used (Charles River Laboratories). Mice were anesthetized by isoflurane inhalation and intranasally inoculated with 1 × 10^5^ fungal cells of PBS-washed overnight cultures (18 hours) of the indicated strains. Mice were sacrificed at 7 days post-infection, and lungs were harvested, weighed, and homogenized in cold PBS. Colony-forming units (CFU) were calculated through quantitative culturing and were represented as CFU/gram lung wet weight. For histopathology at 7 days post-infection, mice were sacrificed using a lethal dose of pentobarbital sodium, and lungs were fixed via intratracheal instillation of neutral buffered formalin under gravity flow (63). Fixed lungs were submitted to the Immunohistopathology Core at Duke University for paraffin embedding and staining with hematoxylin and eosin. Brightfield microscopy images were visualized with a Zeiss Axio Imager A1 microscope (using 4× or 40× objectives) to assess the distribution of the yeast cells and the immune response. Images were taken with an AxioCam MRm digital camera with ZEN Pro software (Zeiss).
Preparation of bone marrow-derived macrophages and in vitro co-culturing with cryptococcal cells
Bone marrow cells were derived from male CD1 mice purchased from Charles River Laboratories. Murine bone marrow cells were prepared and co-cultured with cryptococcal cells as described previously (14, 64). This included the isolation of femurs from mice and flushing bone marrow with 10 mL cold PBS using a 27½ gauge needle. Red blood cells were lysed in 1× red blood cell lysis buffer (0.15 M NH_4_Cl, 1 mM NaHCO_3_, pH 7.4) for 5 minutes and cells were resuspended in 1× Dulbecco’s modified Eagle’s medium (DMEM; +4.5 g/L D-glucose, +L-glutamine, +110 mg/L sodium pyruvate) with 1 U/mL penicillin/streptomycin with 10% (vol/vol) HI-FBS and 3 ng/mL recombinant mouse GM-CSF (BioLegend) for differentiation at a cell concentration of 2.5 × 10^5^ cells/mL in 150 × 15 mm petri plates at 37°C with 5% CO_2_. The media was refreshed after 4 days, and the cells were harvested on day 7.
For co-culturing, C. neoformans cells were washed 2× with PBS, counted, and added to bone marrow macrophages in wells of a 96-well plate at a concentration of 5 × 10^5^ fungal cells per well (10:1 C. neoformans cells:bone marrow macrophages). Co-cultures were incubated for 6 hours at 37°C with 5% CO_2_. Supernatants were collected and stored at −80°C until analysis. Secreted TNF-α was quantified in supernatants by enzyme-linked immunosorbent assay (ELISA; BioLegend). Three replicates were analyzed for each Cn strain.
Differential ubiquitination experiment and analysis
The WT strain (n = 3) and the rsp5Δ mutant strain (n = 3) were incubated in liquid YPD medium for 18 hours. Cultures were normalized to an OD_600_ of 3 in fresh YPD medium and fresh YPD medium buffered to pH 4 and pH 8.15 and conditioned for 1 hour. Samples were washed twice with PBS and flash frozen on dry ice. Sample preparation and mass spectrometry analysis were performed by the Duke Proteomics Core Facility as previously described (28). Differentially ubiquitinated peptides were plotted using the ggplot package in the R coding language using RStudio. We calculated fold changes between the peptides identified from the different strains based on the intensity values and calculated a two-tailed heteroscedastic t-test on log_2_-transformed data for each of these comparisons. We then indicated if a particular peptide was increasing or decreasing with a P-value of 0.05 and fold change of 1.5 between the different strains. Mutants available in a mutant collection lacking the equivalent proteins ubiquitinated by Cn Rsp5, as identified through the differential ubiquitination assay, were then assessed for growth defects in the presence of alkaline pH and a high salt concentration, as well as the inability to induce WT levels of CIG1 expression.
Protein purification and immunoblotting
The WT strain, the rsp5Δ mutant strain, the rim101Δ mutant strain, and the rra2Δ mutant strain were incubated in liquid YPD medium for 18 hours. Cultures were washed twice in PBS and subsequently normalized by OD_600_. Cells were conditioned in fresh YPD medium and fresh YPD medium buffered at pH 4 or pH 8 for 1 hour. To collect total cell lysates, cultures were pelleted, flash frozen on dry ice, and lysed by bead beating. Lysates were collected in 20% trichloroacetic acid (TCA) buffer and processed for western blotting as described before (61). Western blot membranes were stained with Ponceau S stain and then probed with an anti-GFP monoclonal primary antibody (1/5,000 dilution; Roche Applied Science, 11814460001; lot 14717400) and an anti-mouse peroxidase-conjugated secondary antibody (1/25,000 dilution; Jackson ImmunoResearch Laboratories, Inc., 111-035-008; lot 128022), before detection by enhanced chemiluminescence using SuperSignal West Pico (Thermofisher).
The WT strain (H99) and all strains expressing GFP-tagged RRA2, mCherry-tagged VPS23, or both tagged genes were incubated in liquid YPD medium for 18 hours. The strains were then normalized by OD_600_ and conditioned in YPD medium buffered to pH 8.15 for 1 hour. The cells were then fixed with 0.8% (vol/vol) formaldehyde to crosslink protein interactions and quenched with 125 mM glycine. Total cell lysates were prepared as before, and normalized cell lysates were immunoprecipitated through the addition of 25 µL GFP-TRAP or RFP-TRAP agarose bead suspension (ChromoTek) equilibrated in NP-40 lysis buffer, as described previously (21). Lysates were pre-cleared with control agarose beads for 1 hour, and GFP-TRAP and RFP-TRAP agarose beads were blocked with 30 mg/mL bovine serum albumin in NP-40 lysis buffer for 1 hour. Protein eluted from the agarose beads was added to LDS loading buffer and incubated at 95°C for 5 minutes. Western blotting was performed as earlier. To detect precipitation of fluorescent protein, blots were incubated with an anti-GFP monoclonal primary antibody (1/4,000 dilution; Roche Applied Science, 11814460001; lot 14717400) or an anti-red fluorescent protein (anti-RFP) polyclonal primary antibody (1:4,000 dilution, Abcam, ab124754) overnight. An anti-mouse peroxidase-conjugated secondary antibody (1/25,000 dilution; Jackson ImmunoResearch Laboratories, Inc., 111-035-008; lot 128022) was used to allow detection by enhanced chemiluminescence when probing with anti-GFP and an anti-rabbit peroxidase-conjugated secondary antibody (1/25,000 dilution; Jackson ImmunoResearch Laboratories, Inc., 115-035-174; lot 127837 when probing with anti-RFP. A control western blot was performed in parallel with aliquots of normalized cell lysates used as input for the immunoprecipitation experiments. This blot was probed with an anti-PSTAIR monoclonal primary antibody (1/4,000 dilution; Sigma, P7962; lot 015M4840V) and an anti-mouse peroxidase-conjugated secondary antibody as before.
Microscopy
Strains expressing GFP-tagged RIM101 were incubated in liquid YPD medium for 18 hours at 30°C while shaking. Cells were harvested and resuspended in YPD medium adjusted to pH 8.15 and incubated for a further 1 hour at 30°C while shaking. Cells were collected and stained with Hoechst 33342 live nuclear stain for 10 minutes, washed with PBS, and imaged immediately. Differential interference microscopy (DIC) and fluorescent images were visualized with a Zeiss Axio Imager A1 fluorescence microscope (60× or 100× objectives) using a GFP filter for GFP fluorescence. Images were taken with an AxioCam MRm digital camera with ZEN Pro software (Zeiss). The same exposure time was standardized across all strains, and images were analyzed using ImageJ/Fiji software (65).
To obtain high-resolution images of GFP-tagged RRA2 and mCherry-tagged VPS23-expressing cells, we employed confocal microscopy. Cells were prepared as for fluorescence microscopy, and 4 µL of cell suspension was placed in an ibidi 8-well glass bottom µ-slide and covered with a 200 µL slab of solidified 2% agarose (Thermo Fisher Scientific). Specific cell density was adjusted to allow for single-cell monolayers for optimal imaging. Imaging was conducted at room temperature (21°C to 23°C). Images were acquired with a Nikon Ti2-E stand equipped with a Yokogawa CSU-W1 spinning disk confocal unit and a 100×/1.35 silicone oil objective. Images were captured using a Hamamatsu ORCA-Fusion BT sCMOS camera controlled by NIS-Elements software (Nikon Instruments). To capture the entire volume of the cell, 17 Z-slices were acquired at 0.48 µm steps. Cells expressing GFP-tagged RRA2 were imaged at 25% laser power, and mCherry-tagged VPS23-expressing cells at 75% laser power. The exposure time for all confocal images was 400 milliseconds. Images were processed with ImageJ/Fiji software (65).
Phylogenetic tree construction and in silico modeling
Cryptococcus neoformans Rra1 and Rra2 protein sequences were searched against the NCBI GenBank database using a BLASTP search with default settings. Multiple sequence alignments were generated from the top 100 significant BLASTP search hits using Clustal Omega hosted by EMBL-EBI with default settings (66). Phylogenetic trees were constructed using aligned protein sequences as well as the S. cerevisiae Rim8 and A. nidulans PalF protein sequences using IQ-TREE v. 2 and were visualized with iTOL v. 7 (67, 68).
To model potential interactions between the Cn Rra2 protein and Cn Vps23, Google DeepMind AlphaFold v. 3 was used (69). Full protein sequences were uploaded to the AlphaFold server and processed using the default settings. The results, including the predicted aligned error plot, were visualized with ChimeraX, and distances between the two proteins of 5 Å or less were determined (70).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Fernandes TR, Segorbe D, Prusky D, Di Pietro A. 2017. How alkalinization drives fungal pathogenicity. P Lo S Pathog 13:e 1006621. doi:10.1371/journal.ppat.100662129121119 PMC 5679519 · doi ↗ · pubmed ↗
- 2Davis DA. 2009. How human pathogenic fungi sense and adapt to p H: the link to virulence. Curr Opin Microbiol 12:365–370. doi:10.1016/j.mib.2009.05.00619632143 · doi ↗ · pubmed ↗
- 3Prusky D, Yakoby N. 2003. Pathogenic fungi: leading or led by ambient p H? Mol Plant Pathol 4:509–516. doi:10.1046/j.1364-3703.2003.00196.x 20569410 · doi ↗ · pubmed ↗
- 4Robert V, Cardinali G, Casadevall A. 2015. Distribution and impact of yeast thermal tolerance permissive for mammalian infection. BMC Biol 13:18. doi:10.1186/s 12915-015-0127-325762372 PMC 4381509 · doi ↗ · pubmed ↗
- 5Gostinčar C, Zajc J, Lenassi M, Plemenitaš A, de Hoog S, Al-Hatmi AMS, Gunde-Cimerman N. 2018. Fungi between extremotolerance and opportunistic pathogenicity on humans. Fungal Divers 93:195–213. doi:10.1007/s 13225-018-0414-8 · doi ↗
- 6Money NP. 2024. Fungal thermotolerance revisited and why climate change is unlikely to be supercharging pathogenic fungi (yet). Fungal Biol 128:1638–1641. doi:10.1016/j.funbio.2024.01.00538341269 · doi ↗ · pubmed ↗
- 7Weyman-Kaczmarkowa W, Pedziwilk Z. 2000. The development of fungi as affected by p H and type of soil, in relation to the occurrence of bacteria and soil fungistatic activity. Microbiol Res 155:107–112. doi:10.1016/S 0944-5013(00)80045-210950193 · doi ↗ · pubmed ↗
- 8Chakraborty S, Old KM. 1982. Mycophagous soil amoeba: Interactions with three plant pathogenic fungi. Soil Biol Biochem 14:247–255. doi:10.1016/0038-0717(82)90034-7 · doi ↗