Spontaneous mammalian models for Alzheimer's disease and dementia
Madeleine Ford, Frank J Gunn-Moore, Mark P Dagleish

TL;DR
This review explores non-laboratory animal models that spontaneously develop Alzheimer's-like features, suggesting they could improve understanding of dementia.
Contribution
The paper identifies non-laboratory animals as potential models for Alzheimer's disease due to their spontaneous amyloid plaque formation.
Findings
Companion, farm, and marine mammals develop amyloid plaques and some hyperphosphorylated tau.
Few non-human animals develop neurofibrillary tangles or neuronal loss like in human Alzheimer's.
These models may better represent specific aspects of Alzheimer's if further developed.
Abstract
Globally, the human population is ageing, and, consequently, the prevalence of major neurocognitive disorders is increasing, resulting in a greater need for novel dementia therapeutic interventions. Animal models are invaluable in studying underlying pathological processes in human diseases and with evidence for rising life expectancy in many domesticated animals studies have investigated neurocognitive disorders in several non-human species. Rodents have been used extensively as animal models, but this review will examine published literature suggesting candidate non-laboratory animal models for studying dementia, especially human Alzheimer's disease. Comparison of the physiological, pathological and clinical features of companion animals, farm animals and marine mammals shows that although many animals develop amyloid plaques and, to lesser degree, hyperphosphorylated tau protein,…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
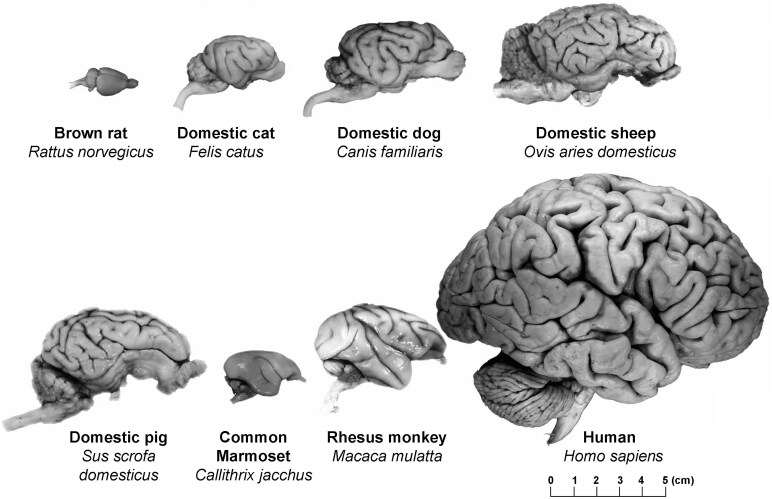 Figure 1
Figure 1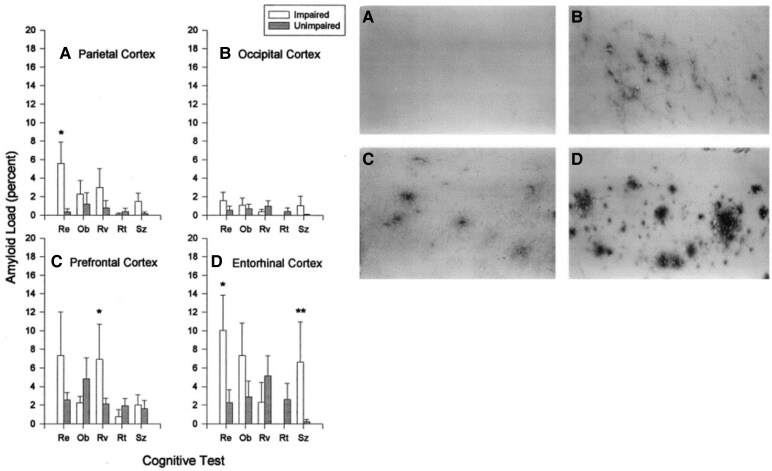 Figure 2
Figure 2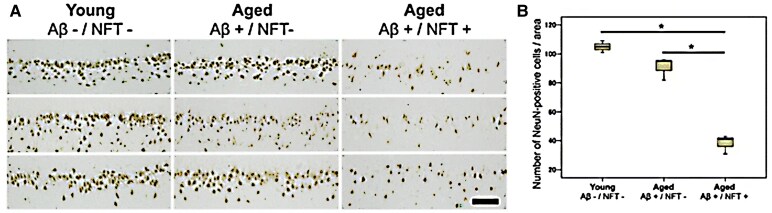 Figure 3
Figure 3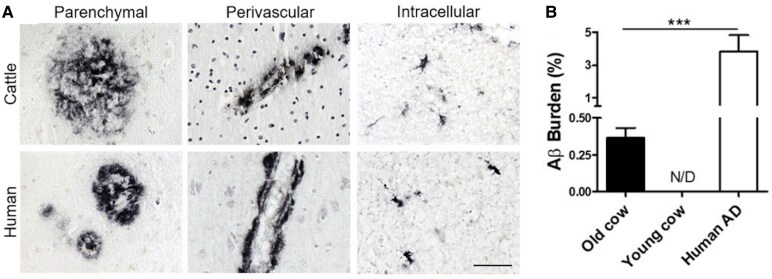 Figure 4
Figure 4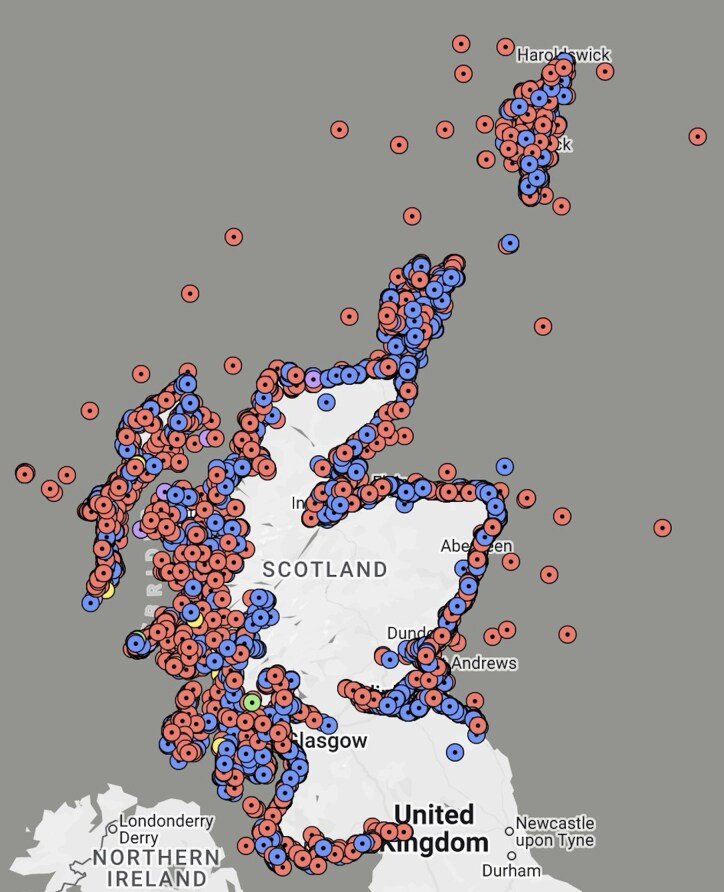 Figure 5
Figure 5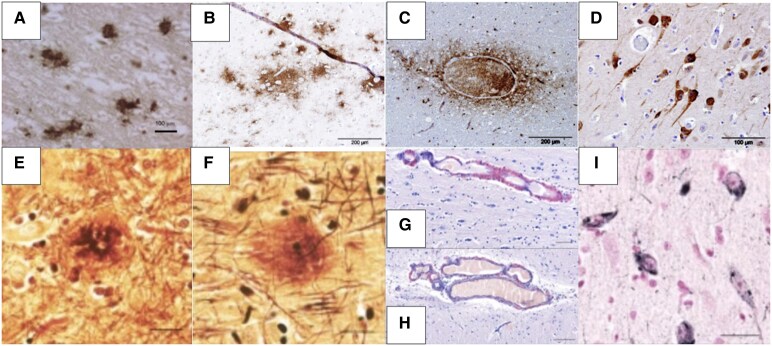 Figure 6
Figure 6| Human | 5′-DAEFRHDSGYEVHHQKLVFFAEDVGSNKGAIIGLMVGGVVIA-3′ |
| Cat | 5′-DAEFRH |
| Dog | 5′-DAEFRHDSGYEVHHQKLVFFAEDVGSNKGAIIGLMVGGVVIA-3′ |
| Chimpanzee | 5′-DAEFRHDSGYEVHHQKLVFFAEDVGSNKGAIIGLMVGGVVIA-3′ |
| Monkey | 5′-DAEFRHDSGYEVHHQKLVFFAEDVGSNKGAIIGLMVGGVVIA-3′ |
| Delphinid | 5′-DAEFRHDSGYEVHHQKLVFFAEDVGSNKGAIIGLMVGGVVIA-3′ |
| Pinniped | 5′-DAEFRHDSGYEVHHQKLVFFAEDVGSNKGAIIGLMVGGVVIA-3′ |
| Mouse | 5′-DAEF |
| Species | ||||||||
|---|---|---|---|---|---|---|---|---|
| Isoform | Human | Mouse | Cat | Dog | Sheep | Pig | Pinniped | Bovine |
| 0N3R | + | − | + | − | − | − | − | − |
| 1N3R | + | − | − | − | + | + | + | + |
| 2N3R | + | − | + | + | + | + | + | + |
| 0N4R | + | + | + | + | − | − | + | − |
| 1N4R | + | + | + | + | + | + | + | + |
| 2N4R | + | + | + | + | + | + | + | + |
| Amyloid pathology | Tau pathology | Brain atrophy | Neuronal loss | Pigment accumulation | Vascular pathology | ||
|---|---|---|---|---|---|---|---|
| Humans | Aβ plaques | NFT | + | + | + | Cerebral amyloid angiopathy | |
| Companion animals | Dogs | Aβ plaques | Hyperphosphorylated tau | + | + | Lipofuscin accumulation | Cerebral amyloid angiopathy |
| Cats | Aβ plaques | Pre-tangles/NFT | D.D. | + | D.D. | Cerebral amyloid angiopathy | |
| Horses | Pre-plaques | Hyperphosphorylated tau | D.D. | D.D. | Lipofuscin accumulation | Vascular degeneration | |
| Farm animals | Pigs | Aβ42 | − | D.D. | D.D. | D.D. | D.D. |
| Donkeys | Aβ plaques | NFT-like structures | D.D. | D.D. | D.D. | D.D. | |
| Cattle | Aβ plaques | D.D. | D.D. | D.D. | D.D. | Cerebral amyloid angiopathy | |
| Sheep | Aβ plaques | Hyperphosphorylated tau/pre-tangles/NFT | D.D. | D.D. | D.D. | Vascular Aβ | |
| Marine mammals | Odontocetes | Aβ plaques | Hyperphosphorylated tau | D.D. | D.D. | D.D. | Vascular Aβ |
| Pinnipeds | Aβ plaques | NFT | D.D. | D.D. | D.D. | Vascular Aβ |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAlzheimer's disease research and treatments · Genetics, Aging, and Longevity in Model Organisms · Neurogenesis and neuroplasticity mechanisms
Introduction
Human life expectancy is increasing due to improved health care with almost 900 million individuals now over 60 years old, and ageing remains the most significant risk factor for neurodegeneration—the progressive loss of neurons in the brain.^1^ With an ageing population, the number of individuals living with a neurodegenerative condition has increased significantly and will continue to increase.^2^ In the UK, an estimated 1 in 14 individuals over 65 years old, and a total of 55 million worldwide, live with dementia, and this is expected to nearly triple by 2050^2,3^ with ∼9.9 million new dementia cases expected every year.^1^
The brain is specifically vulnerable to ageing with, commonly, several processes and functions, such as thinking and memory, declining in older age.^1^ However, people with dementia experience severe cognitive and behavioural changes compared to healthy, ageing adults. Dementia refers to the interference of an individual's daily life, including the ability to remember objects and events, perform simple tasks, utilize speech, remain socially and spatially attentive and regulate emotions due to severe cognitive decline.^4,5^ Many diseases can cause dementia, Alzheimer's disease being the most common and causing approximately two-thirds (>35 million) of all diagnosed dementia cases, but it also includes vascular dementia, dementia with Lewy bodies and frontotemporal dementia.^3,6^
These distinct diseases are, typically, associated with lesions in the brain due to accumulation of pathological proteins (proteinopathy), such as an accumulation of amyloid-beta (Aβ) into plaques or tau proteins into neurofibrillary tangles (NFTs) in Alzheimer's disease, Aβ in vascular dementia, alpha-synuclein in dementia with Lewy bodies or TDP-43 protein in frontotemporal dementia, in the parenchyma and/or vasculature, or secondary to vascular disease and/or a stroke.^3,6,7^ Additionally, dementia sufferers often show a significant loss of brain neurons and brain atrophy. As age-related neuropathology varies between individuals,^6^ distinguishing between normal brain ageing and pathology is crucial. Understanding these pathological processes, the brain regions affected and how these result in clinical disease is critical to finding and developing therapeutics to reduce the impact of neurodegenerative diseases on the ageing population, for which presently there are no cures.^1^
Animal models are invaluable to human research, specifically in studying biological, physiological and pathological processes implicated in many human disorders.^8^ Many of these are based on models in rodents but these species are very short-lived compared to humans. Furthermore, human life expectancy has increased, reducing further the relevance of rodent models. However, the life expectancy of many domesticated animals has increased,^6^ and even though the presence of spontaneous neurodegeneration and subsequent cognitive decline in animals is unclear, a number of studies have identified both similarities and differences compared to humans. For example, relatively recent studies have shown comparable human-like neurodegenerative pathology akin to Alzheimer's disease changes in domesticated companion animals,^6,9-12^ livestock^13-15^ and marine mammals.^16-18^ In addition to pathology, some animals developed clinical signs similar to the social, physical and psychological symptoms experienced by human patients with dementia. As well as furthering the understanding of human cognition mechanisms underlying age-related pathology, studying animals’ behaviours, cognitive abilities and neuropathologies will expand knowledge of their individual cognitive and evolutionary processes.^13,19^
A review of human and animal cognition and associated dysfunction presents a significant research opportunity. Cognitive dysfunction syndrome defines cognitive deterioration in animals not caused by other diseases.^6^ However, evidence suggests that ‘dementia’, currently used only to describe human neurodegenerative diseases, should also include neurodegenerative diseases in other animals. This review will explore the evidence of spontaneous human dementia-like pathology and cognitive dysfunction in companion animals (cats, dogs and horses), livestock (donkeys, pigs, cattle and sheep) and marine mammals (dolphins, seals, sea lions and walruses). It is noteworthy that most of the studies have investigated the pathogenic changes associated with Alzheimer's disease, specifically changes in Aβ amyloid and tau. By defining animal cognitive dysfunction syndrome as dementia in a variety of species, we highlight priorities for future research to aid identifying therapies for human dementia.
The human brain
The human brain is a complex organization of several distinct regions composed of nerve cells and blood vessels responsible for the chemical and electrical signals controlling various processes including thought, memory, emotion and motor function, as well as breathing, heart rate and pain perception.^20^
The longitudinal cerebral fissure divides the human cerebrum into the left and right hemispheres, each of which is further divided into four lobes: frontal, parietal, temporal and occipital.^21^ The hemispheres encompass the cerebral cortex, which receives, processes and interprets information from the five senses: vision, hearing, taste, smell and touch and, arguably, proprioception,^22^ and integrates these separate inputs in the cortex using neuronal networks between these lobes.^20^ Below the cerebral cortex lies the sub-cortex consisting of the basal ganglia, the limbic system (including the thalamus, hippocampus, amygdala and hypothalamus), the cerebellum and the brain stem, the latter continuous with the spinal cord.^20,21^ These brain regions are responsible for movement and posture, emotional processing and vision, amongst other physiological functions necessary for survival including controlling heart rate, blood pressure, sleep cycle and swallowing.^20^
Animals—brains, pathology and behaviour
Studies on ageing processes and neuropathology in animals with cognitive dysfunction syndrome have primarily focused on rodents and invertebrates in laboratory settings due to accessibility and controlled environments.^6,8^ However, larger animals with longer lifespans, such as cats, dogs, equids, pigs and marine mammals, appear to be more promising candidates.^9-18,23^ Additionally, larger mammal brains are gyrencephalic (Fig. 1)^24^ like human brains and have the same highly conserved Aβ peptide amino acid sequence as humans^13,25^ that rodents lack (Table 1).^6^ With respect to tau, many yet not all of the six recognized isoforms present in humans have also been reported in other mammals (Table 2),^16,26^ although similarities and differences have not yet been fully elucidated. However, much of the functional topography in animal brains is unclear, making healthy, and subsequently declining, cognition in live animal subjects challenging to understand.
Comparison of different species’ brains. A visual representation of gyrencephalic brains and mean brain weight (g) in humans (1320), domestic cats (25.6), dogs (50–130), sheep (175), pigs (180) and the rhesus monkey (179) in comparison to the much smaller lissencephalic brain of a brown rat (1.9).24,25
Companion animals
Companion animals are increasingly studied due to their coexistence with humans.^6,30^ As with humans, the lifespans of cats and dogs are increasing, leading to an increase in animals over 10 years old classed as ‘senior’.^31,32^ Age-related cognitive dysfunction syndrome in cats and dogs is widely acknowledged, affecting an estimated 28% of cats aged 11–14 years and dogs aged 11–12 years, increasing to 50% for cats and 68% for dogs aged over 15 years.^6,33-35^ Companion animals unaffected by cognitive dysfunction syndrome remain cognitively intact, ageing normally, a feature found in humans.^6,33^ Comorbidities in ageing animals, including degenerative joint disease, hyperthyroidism and diabetes, can cause altered behaviours confounding cognitive dysfunction syndrome diagnosis. Therefore, diagnosing cognitive dysfunction syndrome requires observing animals’ behaviours during daily activities rather than the interviews and cognitive assessments used in humans.^34,36,37^
In humans, behavioural and psychological symptoms of dementia (BPSD) define the cognitive, psychological and clinical signs that are severe enough to negatively impact or prevent entirely the daily routine of dementia sufferers.^38^ BPSD commonly include memory loss, dysregulation of emotion, thought and personality and motor function deterioration,^4,39^ with specific symptoms of delusion, hallucination, aggression, depression or anxiety, extreme elation and abnormal sleep and wake cycles, amongst others.^38^
Dogs
Ageing dogs with cognitive dysfunction syndrome show disorientation, impaired social interaction, altered house training and changes in sleep–wake cycles when cognitive function is assessed in a wide range of breeds of varying ages.^35,40^ Dogs aged 11–16 years showed progressive cognitive impairment over a 6–18-month period, with greater progression in dogs with at least one impairment at the onset of the study than those without any.^40^ This is similar to humans in that BPSD occur in ∼90% of dementia patients, with ∼50% presenting with at least four BPSD simultaneously.^38,39,41^ The mean age of onset of impairment in dogs is 11 years, ∼75% of the mean 15-year lifespan, and aligns with the mean age of human dementia diagnosis between 70 and 80 years.^28,38^
In dogs, a variety of cognitive tests investigate spatial, discrimination and reversal learning to assess spatial awareness, object and social recognition and memory through task learning and ‘unlearning’.^6,10^ Ageing dogs display human-like age-related social changes, including social withdrawal (e.g. lack of interest in others or available toys) or lack of self-recognition (e.g. increased interaction with own reflection).^10,42^ Studying social interaction in companion animals is invaluable as this cannot be assessed to the same extent in laboratory rodents due to their simplified brain anatomy and lack of training and communication.^6^
Dogs show a stronger association between pathological lesions and behavioural changes than cats, especially cerebral Aβ deposition, hyperphosphorylated tau and neuronal loss.^10,33-35,43,44^ Similar to humans with Alzheimer's disease, ageing impaired dogs often have more Aβ in the prefrontal and the entorhinal cortices (Fig. 2), and dogs aged 2–13 years have a significant correlation between Aβ load in the prefrontal lobe with decreased abilities in object and visual discrimination learning, whereas Aβ load in the entorhinal cortex correlates more with size discrimination and object-approach learning.^10^
*The association between cerebral Aβ plaques and cognitive dysfunction in aged dogs. Left panel: distribution of amyloid in the brains of impaired and unimpaired dogs and the relationship with cognitive tasks; note significant differences in the prefrontal and entorhinal cortices (*P < 0.05; *P < 0.075). Re, reward-approach learning; Ob, object-approach learning; Rv, reversal learning; Rt, retention testing; Sz, size-discrimination learning. Right panel: photomicrographs of Aβ deposition, using immunoreactivity, in the prefrontal cortex of four beagles: (A) young, (B) unimpaired middle-aged, (C) unimpaired aged, (D) impaired aged.10
Additionally, humans develop cerebral amyloid angiopathy, defined as an impaired or inability to clear Aβ causing an accumulation of Aβ in the cerebral arteries, which causes vascular cognitive impairment most commonly in the form of vascular dementia.^45-47^ Similarly, dogs frequently develop Aβ deposition in the cerebral blood vessels causing cerebral amyloid angiopathy^6^ supporting the significance of Aβ pathology causing the clinical signs seen in ageing dogs.
Yu et al.^37^ evaluated the brains of 10 dogs with cognitive dysfunction after ruling out comorbidities and found a significant age-dependent increase in Aβ and hyperphosphorylated tau in impaired dogs’ cerebral cortices and hippocampi compared to healthy controls. However, expression of p-tau Ser396 and accumulation of ubiquitin were significantly increased in the parietal cortex and dorsal part of the hippocampus of the brain of aged dogs compared to humans with Alzheimer’s disease, Braak Stage V, potentially due to faster accumulation in dog brains than human brains in the earlier stages of cognitive dysfunction. Interestingly, no reports of fully formed mature NFTs in aged, cognitively impaired canine brains exist.^6,37,48^ Further investigation of more dogs is necessary, but several possible explanations exist for these discrepancies. One suggestion is that a dog's lifespan is too short to allow mature NFTs to develop; another suggests limited phosphorylation sites exist in dog tau compared to human tau, therefore preventing NFT development.^37,49^ Another is that all the studies that failed to find mature NFTs in dogs used antibody AT8, which recognizes Ser202, a Thr205 epitope of hyperphosphorylated tau,^6,37,48^ whereas one that found early NFTs used an antibody targeting the earlier forming Ser396 epitope.^50^ Furthermore, most research of dementia-like pathology in the brains of dogs has been performed in laboratory beagles, but a study examining other breeds showed breed-related differences in Aβ accumulation, particularly with respect to age of onset.^51^
Cognitively impaired dogs develop additional human-like pathology including neuronal loss and brain atrophy, ventricular enlargement, reduced neurogenesis and increased lipofuscin accumulation.^6,35,42,48^ Yu et al.^37^ reported significant neuronal loss in the cornu ammonis 1 (CA1) hippocampal region in cognitively impaired dogs, with less than half the average number of neurons present in healthy controls, and Siwak-Tapp et al.^42^ estimated up to 30% neuronal loss in the hippocampus and 90–95% neurogenesis decline in aged beagles. Studies found macroscopic brain atrophy with functional topography beginning in the prefrontal and mesial temporal cortices as early as 8 years old, 53% of mean dog lifespan, followed by hippocampal atrophy after ∼75% of the mean lifespan, at 11 years old.^29,42,52^ These regions align with the mesial temporal lobe and the entorhinal cortex of the human brain, which are susceptible to pronounced atrophy in patients with Alzheimer's disease^53^ and suggest, similar to ageing human brains, specific regions of the ageing dog brain are more vulnerable to neurodegeneration.^48,49^ However, another study found no macroscopic atrophy.^37^
It should be noted that laboratory beagles have been used in the pharmaceutical industry for testing potential therapeutic interventions for Alzheimer's disease.
Cats
Less evidence exists for direct clinicopathological correlation between age-related pathology and cognitive dysfunction in cats compared to dogs.^34,48^ However, cognitive dysfunction syndrome in cats is widely acknowledged, and the associated behavioural and psychological changes are described by V-I-S-H-D-A-A-L (vocalization increase; interaction with owners or other pets; sleep–wake cycle changes; house soiling; disorientation; activity changes; anxiety; and learning and memory),^6,54,55^ which are comparable to the BPSD in humans,^38^ although cats in the most recent study did not have a diagnosis of cognitive dysfunction.^52^
Recent discussions have suggested abnormal behaviours in ageing cats include aberrant vocalizations.^55,56^ Although these vocalizations are challenging to determine, they are a likely indicator of cognitive dysfunction syndrome in the absence of other diseases. Černá et al.^57^ investigated unusual vocalization in 37 aged cats with cognitive dysfunction syndrome, upon exclusion of other comorbidities, and owners suggested disorientation, attention-seeking, hunger or pain causing this behaviour in some cases. More extensive studies, including one investigating 883 cats older than 11 years, found excessive vocalization in ∼60%.^58^ Aberrant vocalizations in ageing cats likely align with abnormal and inappropriate outbursts comparable to agitation or aggression displayed by humans with dementia.^38^ Further investigation into the pathological features of cats with aberrant vocalization is necessary to compare this behaviour with humans.
Cognitive tests, similar to those used for dogs,^6,10^ advance the interpretations of owner observations. These tests show an age-related decline in cognitive abilities, the progression of which is estimated to begin around 10 years of age, or 60–70% of average life expectancy of the companion cat, which is considered geriatric by 15 years,^55,56^ aligning with the average age at which dementia symptoms occur in humans.^38^
Examination of ageing cat brains shows Aβ plaques in the neuropil and the cerebral blood vessels,^34,48^ although some suggest cats only develop diffuse deposits of Aβ representative of early or pre-plaque formation.^30,33,59,60^ In humans, dementia-associated plaques consist predominantly of Aβ composed of peptides of 40 or 42 amino acid residues (Aβ40 and Aβ42, respectively).^6,61^ Aβ42, due to its larger size, lower solubility and increased fibrillization, is more abundant than Aβ40, but both are present.^62^ The plaques found in ageing cat brains are composed of Aβ42, not Aβ40, whereas the vascular amyloid angiopathy is composed of both Aβ40 and Aβ42,^33,59^ identical to the vascular amyloid composition in humans with dementia.^62^ The relationship between amyloid pathology and cognitive dysfunction in ageing cats is debated as there is no clear evidence of correlation between the amount and distribution of Aβ in the brain with the severity of cognitive dysfunction.^30,33,48^
Ageing cats with cognitive dysfunction syndrome rarely display fully formed NFTs,^30,33,34^ but they do have five of the six normal hyperphosphorylated tau isoforms identical to humans.^26,34^ Hyperphosphorylated tau is neurotoxic in humans, causing a loss of the protein's normal function and promoting aggregation.^63^ As such, the hyperphosphorylated tau in ageing cat brains suggests the possibility of pre-tangles that could develop later into pathological NFTs, but the shorter lifespans of cats compared with humans may reduce this phenomenon.^34,49,55^ However, one study reported an association between the presence of pre-tangles and cognitive dysfunction syndrome,^60^ whilst another study reported fully formed NFTs present in the hippocampus of a few aged cats.^64^ Comparisons of neuronal loss in cats with and without Aβ or NFTs found a significant correlation between neuronal loss and NFT formation as aged groups without NFT pathology showed similar neuron load to young cats with no Aβ accumulation or NFT (Fig. 3).^64^ This suggests NFT pathology is more closely associated with neuronal loss than Aβ accumulation in cats and possibly a more accurate biomarker for cognitive decline in ageing cats.^55^ Chambers et al.^64^ measured neuronal loss in cats post-mortem using immunohistochemistry and found that Aβ accumulation precedes tau accumulation and neuronal loss in ageing cats.
*Neuronal load in the hippocampal regions of young and aged cats. (A) Immunohistochemistry detecting NeuN (a protein used to detect mature neurons) load in one young (Aβ−/NFT−; n = 3; mean age = 3.7 years) and two aged groups (Aβ+/NFT−; n = 3; mean age = 18 years and Aβ+/NFT+; n = 3; mean age = 17.6 years) with or without Aβ and NFT pathology (scale bar = 100 µm). (B) Quantification of the number of NeuN-positive cells in the hippocampal regions of the cats.64 P < 0.001. Aβ, amyloid-beta; NFTs, neurofibrillary tangles.
Although domestic cats and dogs display features of human dementia, further investigation should focus on the differences such as the absence of NFT in aged, cognitively impaired dogs yet its presence in some aged cognitively impaired cats. As hyperphosphorylated tau exists in these animals and is a hallmark pathological feature of Alzheimer's disease, understanding the progression from hyperphosphorylated tau to NFT formation and its implications on cognitive decline is critical to determine the full potential of these animals as research models for this disease.^48^ Chambers et al.^64^ suggest that cats develop NFT, but aged dogs do not, possibly due to the involvement of Aβ in furthering tau pathology, and vice versa. As such, one hypothesis highlights the disparity in amyloid peptide amino acid sequences between cats and dogs (Table 1).^64^ Human and dog amyloid peptide amino acid sequences are 100% homologous.^29^ However, cats differ at the seventh residue, and it has been suggested this promotes more extensive Aβ accumulation.^64^ It is likely that this greater amyloid load triggers increased tau pathology, resulting in the NFT formation found in cats, but not in dogs.^64^
Farm animals
Farm animals are increasingly studied as potential models for human dementia due to their similar size and complexity with humans compared to rodents (Fig. 1).^25^
Pigs
The pig brain closely resembles the human brain,^13,65,66^ being gyrencephalic with similar development in late gestation and early postnatal life.^13,66^ Furthermore, the prefrontal cortex, a key region in human neurodegeneration, is ∼10% of the pig brain, similar to humans.^13^
Pigs live in small groups composed of a female and her offspring, and adult males are solitary, with interaction only occurring during the mating season. Pigs recognize familiar people and objects, but as the porcine visual system is less developed than non-human primates and humans, much of this recognition relies on auditory and olfactory senses.^13,67^ Additionally, like humans, pigs are able to relate cognition with emotion, displaying positive or negative reactions to situations in adapting to changing environments. For example, pigs recognize danger and associate negative emotions such as fear or anxiety with them, in the form of ‘freezing’, ‘standing alert’, defaecation and urination, and vocalizations.^13^ These complex cognitive abilities make pigs potentially better models for studying human dementia as they may be more permissive to cognitive testing.^13,65^
Pig behaviour, spatial discrimination and cognitive ability have been investigated using mazes.^13,66^ However, studying cognitive decline in pigs has limitations, including size, cost, space requirements and specialized housing,^65^ which are all significantly less in rodent studies. However, the larger brains and the existence of tau isoforms in pigs that are identical to human isoforms raise the potential of pigs as candidates for studying human dementia.^65^
Similar to the induction of the amyloid precursor protein (APP) gene to trigger the production of Aβ in the transgenic mouse model,^68^ Kragh et al.^69^ developed the first transgenic pig model using a mutation associated with early onset of Alzheimer's disease in humans to induce expression of human APP, and this induced increased expression of Aβ42 and accumulation,^65^ the primary component of human amyloid plaques.^61^ Although tau pathology and NFT formation have not been identified in ageing pigs, they do express four of the six isoforms found in humans,^26^ whereas wild-type rodents have no human tau isoforms.^65^ Compared with other animals, interest in pigs for cognitive research is relatively new^13^; therefore, more extensive studies on ageing pig pathology and behaviour are required to more accurately correlate pathology with cognitive decline.
Equids
Horses’ cognition is considered high due to their use in sport, work and human companionship,^70^ and so their relationship with humans is similar to dogs and cats rather than farm animals. This provides an opportunity to study the social interaction components of cognitive function and subsequent dysfunction.^70^ Evidence suggests horses have complex abilities of discrimination and memory. Lampe and Andre^71^ reported that horses could use visual, olfactory and auditory cues to recognize individual humans and associate positive or negative emotions with those individuals, suggesting a relationship between affect and cognition, as found in pigs.^13^ Although horses have not been systematically exposed to as many cognitive tests as cats or dogs, observations of memory loss indicators (e.g. confusion, disorientation and altered social behaviour) in aged horses have been reported.^6^
Two studies have investigated pathological lesions in ageing horses: one that studied 60 horses aged 7–23 years^12^ and the other that studied 100 horses aged 2–25 years.^11^ Both studies reported age-related increases in neuronal lipofuscin accumulation and age-related vascular degeneration and neuronal vacuolation.^11,12^ Lipofuscin pigment accumulation in neurons is associated with ageing in humans, commencing at age 9 and increasing with age,^72-74^ and abnormal aggregation and accumulation is present in many neurodegenerative diseases. While neither study in horses found amyloid plaques, possibly due to the use of Congo Red stain only rather than immunohistochemistry, or NFTs, Capucchio et al.^12^ reported tau-positive neurons and methenamine-positive diffuse plaques potentially indicating pre-NFT and pre-amyloid plaques, respectively, but only one of the horses displayed both. Along with horses, dogs and cattle have also shown specific aberrant lipofuscin accumulation in line with other neuropathology and signs of cognitive dysfunction,^6,75^ and this may be a potential biomarker for dementia-like pathology in non-human animals.
Although horses have been at the forefront of studies in equids, Malbon et al.^15^ conducted a post-mortem study on 13 donkeys aged between 33 and 44 years, with two younger controls (aged 8 and 9), to assess donkeys as a potential model for human neurodegeneration. Aβ plaques were identified in nine and NFT-like structures in seven of the aged donkeys, whilst neither of the younger donkeys displayed either pathology. Furthermore, because the oldest donkey in the sample (aged 44) displayed neither pathology, it is suggested that these lesions may not be part of the normal ageing process in donkeys.^15^ Additionally, this study highlighted the potentially significant association between Type II diabetes mellitus, obesity and Alzheimer's disease found in humans and the role of insulin dysregulation in these processes.^76^ Interestingly, equine metabolic syndrome is common in several equid species and is the most common endocrine disorder in donkeys.^15,77^ In equine metabolic syndrome, obesity causes reduced insulin clearance and adipokine (e.g. leptin and adiponectin) dysregulation, which can lead to hyperinsulinaemia.^15,77^ In humans, leptin and adiponectin are closely associated with Alzheimer's disease development^78^ and therefore present a promising opportunity for research in donkeys into their roles.
As the study by Malbon et al.^15^ remains the only published study that explored Alzheimer's disease-like pathology in donkeys and used brains from animals already dead, behavioural assessments were not obtained prior to death. Therefore, further investigation is required to fully understand how the pathology identified relates to human neurodegeneration.
Cattle and sheep
Like many other animals investigated, the Aβ sequence in cattle is identical to humans.^14^ A study using an anti-human Aβ antibody labelled human-like mature Aβ plaques in the cerebral cortices and hippocampi of 14 cattle over the age of 13 years (n = 63, age 13–23 years), and labelling was also present in the perivascular spaces, indicating cerebral amyloid angiopathy-like pathology (Fig. 4).^14^ All negative control cattle (n = 10, 10 months old) were devoid of labelling. The Aβ burden was much lower in aged cattle than humans with Alzheimer's disease, but the accumulation in cattle was age dependent (Fig. 4). However, another study in cattle (n = 102, age range 0–240 months) found intraneuronal and extra-neuronal deposits increased with age but also failed to find any Aβ plaques and suggested this may be due to a lack of β-sheet secondary protein structures in bovine Aβ.^79^
Comparison of amyloid load in cattle versus humans. (A) Anti-Aβ antibody labelling detecting parenchymal, perivascular and intracellular amyloid deposits in the temporal regions of aged cattle and aged human brains (scale bar = 50 µm). (B) Percentage of the brain occupied by Aβ plaques in the brains of old cattle (n = 63; age 13–23 years; Aβ burden 0.37 ± 0.07%) in comparison to healthy young cattle group (n = 10; age 10 months; Aβ burden not detected) and a human Alzheimer's disease group (Aβ burden 3.82 ± 1.0%). N/D denotes no Aβ aggregation detected.14
Sheep have been examined for suitability for genetic manipulation to produce a large animal model of Alzheimer’s disease due to the amino acid sequences of ovine and human Aβ40 and Aβ42 being identical, that of APP being highly conserved and the total levels of tau protein in cerebrospinal fluid being comparable.^80^ Furthermore, using anti-Aβ and phosphorylation-specific (AT8 and S396) anti-tau antibodies in the hippocampal and entorhinal regions of 30 sheep brains (estimated ages 1–2 years, 2–3 years and >5 years), Davies et al.^81^ reported both intracellular and vascular Aβ in sheep ≥2 years old, phosphorylated tau increasing with age and pre-NFT were present in sheep estimated to be >5 years old. Sheep express four isoforms identical to human tau, especially the 3R and 4R human isoforms, and in their pilot study, Davies et al.^81^ reported mature NFT formation in a 21-year-old sheep.
Although farm animals have the potential for developing spontaneous dementia-like pathology and cognitive decline, clinical investigation is more complicated than in laboratory or companion animals.^13^ The studies on horses,^11,12^ donkeys,^15^ cows^14^ and sheep^81^ were conducted on brains collected post-mortem with no ante-mortem assessment of cognitive decline undertaken. Farm animals are challenging to train for behavioural observations, making cognitive assessment difficult, but not impossible. However, given the social interactions between some equids and humans,^82^ further research on cognitive abilities in horses and donkeys may identify clinicopathological correlations. Also, as sheep show spontaneous development of Aβ deposition, pre-NFT and potentially mature NFT formation in old age^81^ like humans, therefore studying their behaviour may be beneficial too. It is worth noting that some transgenic pig^83^ and sheep^84^ models have been developed for neurodegenerative diseases other than Alzheimer's disease such as Huntington's disease.
Marine mammals
Marine mammals have been subjected to the least amount of neurodegenerative research due to the difficulty in studying the progression of neurodegeneration and cognitive decline during the animals’ lifetimes. Thus, much of the research has focused on brains recovered after death due to stranding. Nonetheless, evidence for dementia-like pathology has been observed in several studies of odontocetes (toothed whales and dolphins)^18,85^ and pinnipeds (sea lions, seals and walruses),^16^ raising their importance as potential natural spontaneous models of dementia.^29^
Odontocetes
Odontocetes have grown in interest in cognitive dysfunction research for various reasons including comparable lifespans to humans, identical Aβ amino acid sequence to human Aβ in several odontocete species^29,85^ and the cooperative living practices of numerous species.^18^ Of particular interest is their epimeletic, or caregiving, behaviour towards injured or diseased individuals such as carrying an individual, keeping it afloat or protecting it from further danger that have all been observed more frequently in odontocetes than in other marine mammals.^18,86^ This behaviour is important in the study of neuropathology as the disease can progress further, allowing more advanced lesions to develop compared with solitary individuals that are less likely to survive to more advanced stages of the disease.^18^ Furthermore, epimeletic behaviour aligns more with human behaviour, allowing comparative evaluation of the social interaction components of cognition. The Scottish Marine Animal Stranding Scheme (SMASS)^87^ investigates stranded cetaceans, pinnipeds, marine turtles and sharks along the Scottish coast (Fig. 5) and was a source of material for a comprehensive study.^18^
Cetacean, pinniped, seas turtle and basking shark strandings in 2021 in Scottish coastal waters. A map of 17 species of cetacean (red), pinniped (blue), turtle (yellow), basking shark (green) and other shark species (purple) reported stranded along the coast of Scotland in 2021 (Taken from SMASS, 2021—Map of Strandings).87
A recent study by Vacher et al.^18^ investigated brains of 22 individuals from five different odontocete species that had stranded in Scotland. Using evidence of old age, such as worn or lost teeth, relative increases in cerebral white matter or life history-related photo identification, 18 of the 22 dolphins were designated as aged, including Risso's dolphins (Grampus griseus), long-finned pilot whales (Globicephala melas), white-beaked dolphins (Lagenorhynchus albirostris), harbour porpoises (Phocoena phocoena) and a bottlenose dolphin (Tursiops truncatus), and four were designated as young adults or subadults. All 18 aged odontocetes showed intracellular Aβ accumulation, especially in the limbic and paralimbic lobes that, in the odontocete brain, overlay the basal ganglia and the thalamus, respectively.^18,88,89^ Additionally, in the oldest animals, significant amounts of both intra- and extracellular deposits of Aβ were present in the brain regions corresponding to those in humans with Alzheimer's disease.^18^ Of the 18 aged odontocetes, three individuals (one L. albirostris, one G. melas and one T. truncatus) showed Aβ plaques, probable cerebral amyloid angiopathy (vascular Aβ accumulation) and hyperphosphorylated tau. This suggested a pre-NFT biomarker, and further investigation of these aged odontocetes showed both Aβ and tau aggregations in additional brain regions that were not observed in the four young adults or subadults, suggesting spontaneous, age-related protein aggregation and progression.^18^
Comparing odontocete brains with human and terrestrial animal brains is difficult as much of the odontocete brain has structurally adapted to an aquatic environment.^18,88,89^ However, the Aβ plaques found in the odontocetes’ brains by Vacher et al.^18^ were predominantly in the rostrolateral cortices, called the orbital lobe in odontocetes. The orbital lobe contains the limbic and paralimbic lobes and is considered homologous with the human frontal lobe.^18,89^ This aligns with the initial accumulation of amyloid plaques in the frontal, temporal and occipital lobes of human brains in Alzheimer's disease.^18,90^ Despite the similar overall size and complexity of the odontocete brain and the inclusion of all the human hippocampal regions (dentate gyrus, subiculum, hippocampus and entorhinal cortex) in the odontocete hippocampus, the latter is much smaller than in humans, estimated at only 10% of the size.^88,89^ These disparities in regional size raise curiosity about functional localization in odontocete brains. A reduction in the size of the hippocampus suggests a smaller reliance on this structure, with other more developed structures necessary for aquatic survival. Oelschläger^88^ suggested the reduced hippocampal size may correspond to the reduced olfactory system in cetaceans. Alternatively, as the hippocampus is associated with movement and spatial memory, a reduced hippocampus may be less vulnerable to injury, thereby reducing behaviours that may cause stranding.^18,88^ Unfortunately, in the archived brain samples used by Vacher et al.,^18^ the hippocampi were not sampled.
Evidence of Alzheimer's disease-like pathology, amyloid plaques and NFT, in odontocetes is insufficient to diagnose Alzheimer's disease as this requires identification of significant clinical cognitive dysfunction. However, studying odontocete behaviour is challenging as samples from free-ranging animals are only available after stranding. Although there are no definitive reports of cognitive decline in ageing odontocetes, the animals studied by Vacher et al.^18^ and others reported by SMASS^87^ (Fig. 5) all stranded. Live-strandings could result from pathology-associated disorientation, aligning with the first signs of dementia in humans and commonly presenting as confusion and poor sense of direction.^18,91^ The ‘sick-leader hypothesis’ has been proposed as a possible cause of mass-strandings in marine mammals, whereby an impaired leader erroneously leads a group into shallow waters.^18,91^ These conclusions, however, are purely speculative.
To investigate this hypothesis, further studies must compare Alzheimer's disease-like pathology with behavioural disparities, and this may be possible in captive odontocetes. Stylianaki et al.^92^ investigated Alzheimer's disease-like pathology in a 40-year-old captive bottlenose dolphin (T. truncatus) finding parenchymal (in the equivalent of the human frontal, temporal and parietal lobes) and vascular Aβ plaques. No mature NFTs were reported, but this study showed apolipoprotein E (APOE) immunoreactivity, similar to human Alzheimer's disease, suggesting a human-like involvement of the APOE gene in Aβ plaque formation.^92^ This dolphin showed no signs of cognitive dysfunction but was found with undigested fish in the stomach (usually indicative of sudden death as there is rapid transit of ingesta in cetaceans) and evidence of drowning, which may indicate some form of memory loss. Again, these conclusions are highly speculative and require further investigation.
Another suggestion for the observed neuropathology in odontocetes relates to their life histories. Some odontocetes are known to be deep-diving, frequently reaching depths of 1000 and even 3000 m, which a Cuvier's beaked whale (Ziphius cavirostris) surpassed for over 2 h.^85^ Diving to such depths repeatedly increases the risk of significant episodes of cerebral hypoxia, despite adaptation to such conditions, and hypoxia increases the risk of neurodegenerative pathology.^85^ Neuroglobin, a haem protein expressed within neurons with a higher affinity for oxygen than haemoglobin, enables oxygen to move from the vascular compartment to the neural tissue, providing oxygen stores to afford some protection under hypoxic conditions.^85^ However, diving mammals have the same amount of neuroglobin as non-diving mammals, so this mechanism does not negate the risk of hypoxia.^85^ Similar to ageing, cerebral hypoxia can lead to hypometabolism, which is known to trigger APP overexpression and prevent homeostatic clearance of Aβ aggregates in humans with dementia and can cause inflammation, oxidative stress and neuronal loss.^93^ These processes may be implicated in the neuropathologies reported in odontocete species.
Pinnipeds
Although odontocetes have been more extensively investigated, several studies have assessed dementia-like pathology in aged pinnipeds, including seals (Phocidae), sea lions (Otariidae) and walruses (Odobenidae), which have also an identical Aβ peptide sequence to humans (Table 1). Takahashi et al.^16^ investigated the brains of 10 pinnipeds (four seals, five sea lions and one walrus) aged 0–35 years, and amyloid plaque pathology was present in eight of them (aged 27–35) in various combinations of the frontal, parietal, temporal and occipital lobes of the cerebral cortex, but none in the white matter, brain stem or cerebellum. Plaques were also present in the blood vessels, indicating cerebral amyloid angiopathy-like pathology (Fig. 6). Additionally, both neuritic plaques, with a distinct compact core, and diffuse plaques, devoid of a distinct core, were present, and both had a higher composition of Aβ42 than of Aβ40,^16^ aligning with Alzheimer's disease in humans.^61^ The depositions were considered to be age dependent as none were found in the two infant or juvenile pinnipeds examined, whereas all eight aged pinnipeds had amyloid plaques. Takaichi et al.^16^ also found an increased number and size of astrocytes and microglia with enlarged pre-synapses around amyloid deposits, a feature found commonly around amyloid plaques in humans suggestive of increased inflammation.^94^ Pinnipeds express five of the six human tau isoforms (1N3R, 2N3R, 0N4R, 1N4R and 2N4R), and significant NFT formation with fibrillar aggregation was reported in these pinnipeds.^16^
Neuropathology in aged cetaceans and pinnipeds. Amyloid plaques (brown pigment) in (A) a striped dolphin (Stenella coeruleoalba)29 (scale bar = 100 μm); (B) diffuse plaques in an aged bottlenose dolphin (T. truncatus)18 (scale bar = 200 μm); (C) amyloid plaque deposits around blood vessels indicating cerebral amyloid angiopathy in a white-beaked dolphin (L. albirostris)18 (scale bar = 200 μm); (D) axons and dendrites containing hyperphosphorylated tau (brown pigment) in an aged long-finned pilot whale (G. melas)18 (scale bar = 100 μm); (E–I) pathology in a 32-year-old Australian sea lion (Neophoca cinerea) showing (E) neuritic plaques (dark red pigment) with a dense core16 (scale bar = 20 μm); (F) diffuse plaques without a distinct core16 (scale bar = 20 μm); (G) blood vessels in the meninges with amyloid aggregation (red pigment)16 (scale bar = 50 μm); (H) blood vessels in the cerebral cortex with amyloid aggregation (red pigment)16 (scale bar = 100 μm); and (I) NFTs (black pigment) in the neurites of the cerebral cortex (scale bar = 50 μm).16
Pinnipeds have also shown human-like vascular Aβ deposits colocalized with APOE in the arterial and capillary walls.^16^ The eldest sea lions (aged 27–32 years) showed more severe cerebral amyloid angiopathy pathology compared to the younger pinnipeds and followed a similar regional progression in the frontal, temporal, parietal and occipital lobes^16^ as humans with dementia.^46^ Although the findings of this report seem promising, it was the first study to report spontaneous Alzheimer's disease-like pathology in aged pinnipeds and further investigation is required.
Discussion
With an ageing population and an increase in dementia cases worldwide, the search for a cure or even a treatment to slow the progression of cognitive decline is critical. Animal models are vital in scientific research to study and understand many human diseases and their pathogeneses. Although rodents have been used extensively for cognitive research, due to ease of use, accessibility, cost and ethical and legal restrictions, recent studies have suggested that larger animals with brain anatomy and lifespans more similar to humans should be considered. Through analysis of the published literature, this review has identified animal species that spontaneously develop age-related neurodegeneration. Youssef et al.^6^ published a review of the evidence for dementia-like pathology in cats, dogs, horses and primates, discussing some of the behaviours likely to be associated with this pathology. However, this review provides a deeper understanding of the pathological features and several additional animal species and highlights the clinicopathological correlations in all these animal species.
Ambiguity remains with respect to the pathological lesion(s) most representative of neurodegeneration. Several studies propose it is the sequence of onset and progression of amyloid and tau pathologies,^95-97^ whilst others suggest that neuronal loss is the most important pathological feature.^41,98-100^ Furthermore, loss of synapses is considered to be the strongest correlate of cognitive decline in Alzheimer's disease.^101-104^ As the primary proteinopathies in Alzheimer's disease, amyloid and tau accumulations represented by Aβ plaques and NFTs, respectively, have been the primary focus of studies of pathology relating to neurodegeneration in ageing animals (Table 3). However, to fully assess these non-laboratory animal species as potential models of wider human dementia, it is essential that the other proteinopathies present in humans, such as α-synuclein, Lewy bodies and trans-activation response element DNA binding protein 43 (TDP-43), are also investigated.^3,6,7^ Therefore, the significant proteinopathy and vascular pathologies present in several species of ageing animals represent promising subjects for further investigation (Table 3). Ultimately, to identify the animal(s) that model human diseases most accurately, all aspects of Alzheimer's disease pathology must be thoroughly studied, which will include determining other diseases responsible for causing cognitive dysfunction in these various animal species.
Studies of non-laboratory animal pathology have been limited in sample size, especially marine mammals due to their habitats limiting investigations of their brains to, primarily, those of stranded animals post-mortem.^16-18,85^ Furthermore, age is a critical factor in dementia, and the ages of the marine mammals available are usually estimated via subjective features that may lack accuracy.^18^ Impaired cognition is a critical component of human dementia diagnosis. Therefore, understanding impaired cognition and behaviour in animals is essential to identify the animal species that best models human dementia. As the conclusions drawn from many of these studies are based on extrapolations and speculations, larger sample sizes, ante-mortem assessments and full post-mortem investigations are required to validate any conclusions based on the cause of death, age at death and evidence of behavioural change and cognitive decline, and this must correlate with the types and extents of the pathology present.
However, there are several ethical and logistical limitations to studying behaviour in non-laboratory animals to the same extent possible in humans, resulting in dementia research in animals focusing primarily on post-mortem studies as behavioural observations were unavailable. Animal welfare concerns in several of the discussed candidate models pertain mainly to pain and suffering, general health and cognitive welfare. Although some animal research requires only observation of an animal's behaviour in their natural habitat, some can be invasive, harmful and/or painful to the subjects. The UK Animal Welfare Act 2006 holds owners legally accountable for ensuring the welfare of livestock and companion animals and preventing unnecessary pain and suffering.^105^ Additionally, the UK Parliament has recently dedicated discussion to the ethics and practicality of using animals in scientific and medical research, covered by the Animals (Scientific Procedures) Act 1986^106^ and its various amendments, and if legislation changes, some candidate animal species may become less available for scientific research. Moreover, 41 000 species are currently threatened by extinction,^107^ including many marine mammals, and this raises ethical considerations in the search for an animal model for better understanding human dementia, irrespective of how valuable such a model may be.
Conclusion
Identifying and validating animal models of human diseases is complex and time-consuming. Human Alzheimer's disease and other dementias are, at present, devoid of any valid animal models representing of all the associated pathophysiological processes and behaviours of any individual disease, and also for assessing potential therapeutics. Although no single animal species develops all the characteristics of any human dementia, many develop proteinopathies, resulting in dementia-like pathology, and some have comparable and correlated cognitive dysfunction. Further and more comprehensive investigation of the pathological features and their correlations with clinical signs, especially cognitive dysfunction, in complex animals will identify the most appropriate model for studying this devastating, yet increasingly prevalent, group of diseases.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alzheimer’s Disease International . World Alzheimer Report 2015 The Global Impact of Dementia An analysis of prevalence, incidence, cost and trends . Accessed 23 January 2023. https://www.alzint.org/u/World Alzheimer Report 2015.pdf
- 2Alzheimer’s Research UK. Prevalence and incidence of dementia . Accessed 21 January 2023. https://dementiastatistics.org/about-dementia/prevalence-and-incidence/
- 3Alzheimer’s Disease International. Dementia facts & figures . Accessed 03 April 2023. https://www.alzint.org/about/dementia-facts-figures/
- 4Dementia UK. What is dementia? . Accessed 21 January 2023. https://www.dementiauk.org/information-and-support/about-dementia/what-is-dementia/
- 5World Health Organization (WHO). ICD-11 for Mortality and Morbidity Statistics . Accessed 30 March 2023. https://icd.who.int/browse/2024-01/mms/en#213458094
- 6Youssef SA, Capucchio MT, Rofina JE, et al Pathology of the aging brain in domestic and laboratory animals, and animal models of human neurodegenerative diseases. Vet Pathol. 2016;53(2):327–348.26869150 10.1177/0300985815623997 · doi ↗ · pubmed ↗
- 7Pendlebury ST, Rothwell PM. Prevalence, incidence, and factors associated with pre-stroke and post-stroke dementia: A systematic review and meta-analysis. Lancet Neurol. 2009;8(11):1006–1018.19782001 10.1016/S 1474-4422(09)70236-4 · doi ↗ · pubmed ↗
- 8Phillips NLH, Roth TL. Animal models and their contribution to our understanding of the relationship between environments, epigenetic modifications, and behavior. Genes (Basel). 2019;10(1):47.30650619 10.3390/genes 10010047 PMC 6357183 · doi ↗ · pubmed ↗