The −10 region adjacent to open reading frames is a common expression pattern in Deinococcus-Thermus
Shitong Zhong, Linjia Wang, Shuang Song, Liangyan Wang, Yuejin Hua, Huizhi Lu

TL;DR
This study shows that a specific DNA region called the −10 motif is commonly used to start gene expression in the Deinococcus-Thermus phylum, often without a long RNA leader.
Contribution
The study identifies the −10 motif as a key promoter region for leaderless mRNA in the Deinococcus-Thermus phylum.
Findings
The −10 motif functions as a promoter for leaderless gene transcription in Deinococcus radiodurans.
The −10 motif is commonly found upstream of ORFs in the Deinococcus-Thermus phylum.
Adding a −35 region can enhance transcription levels in this system.
Abstract
This study reveals that the promoter −10 region adjacent to open reading frames for gene expression via leaderless mRNA constitutes an important mode of gene expression in the Deinococcus-Thermus phylum. Gene transcription and translation are fundamental processes that underpin the vitality of living organisms. In bacteria, the classic promoter elements for transcription include the −35 region and the −10 region, whereas the ribosome binding site (RBS) is essential for translation. Our investigation of the upstream sequences of ORFs in Deinococcus radiodurans has confirmed a −10 region-like motif, which appears as 5′-TANNNT-3′ (−10-motif). Through rigorous experimental validation, we have established that this −10-motif functions as the classical −10 region of the promoter during transcription, responsible for initiating transcription of leaderless genes (mRNA products that do not have…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
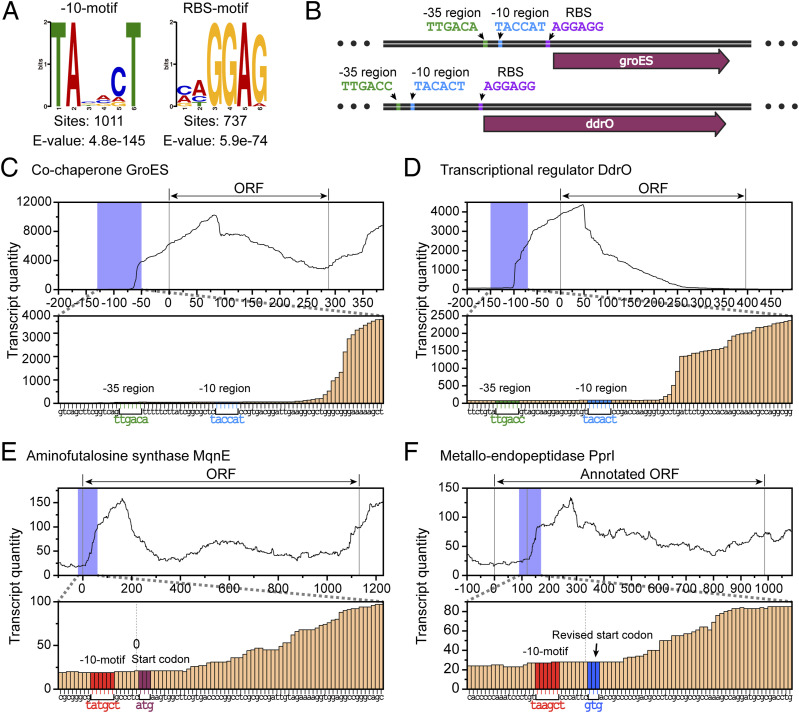 Figure 1
Figure 1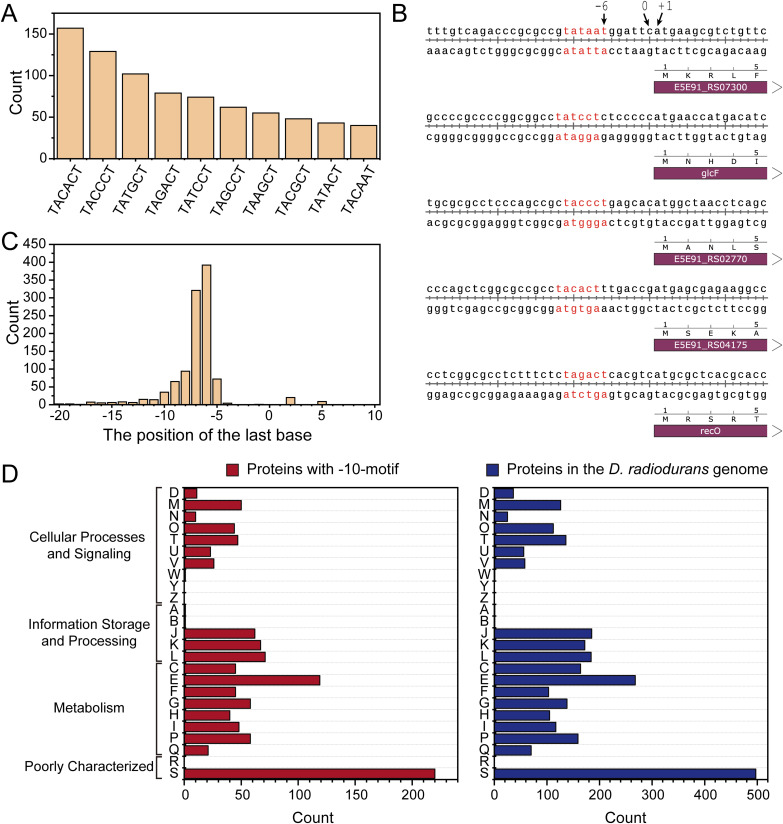 Figure S1
Figure S1 Figure S2
Figure S2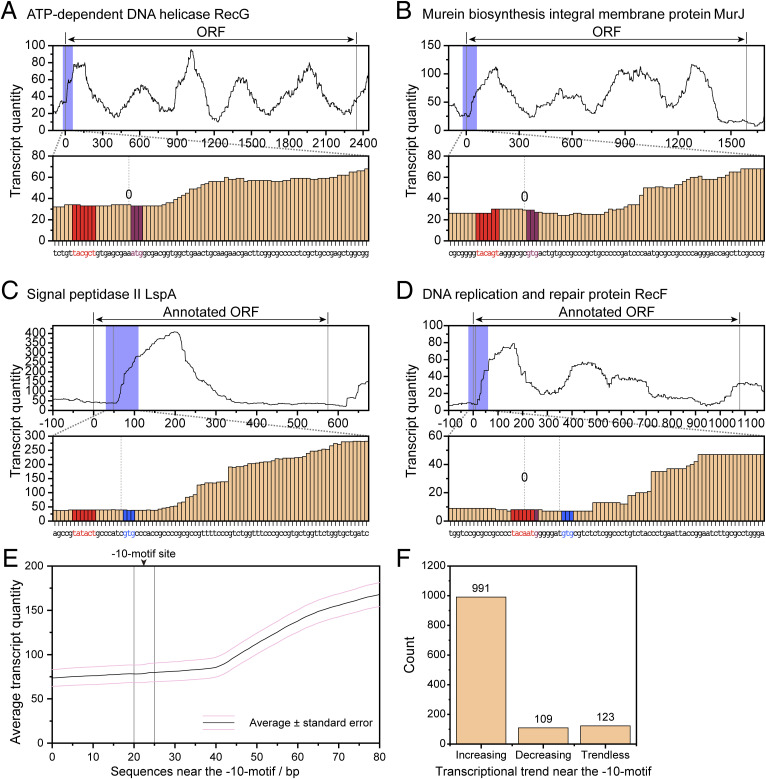 Figure S3
Figure S3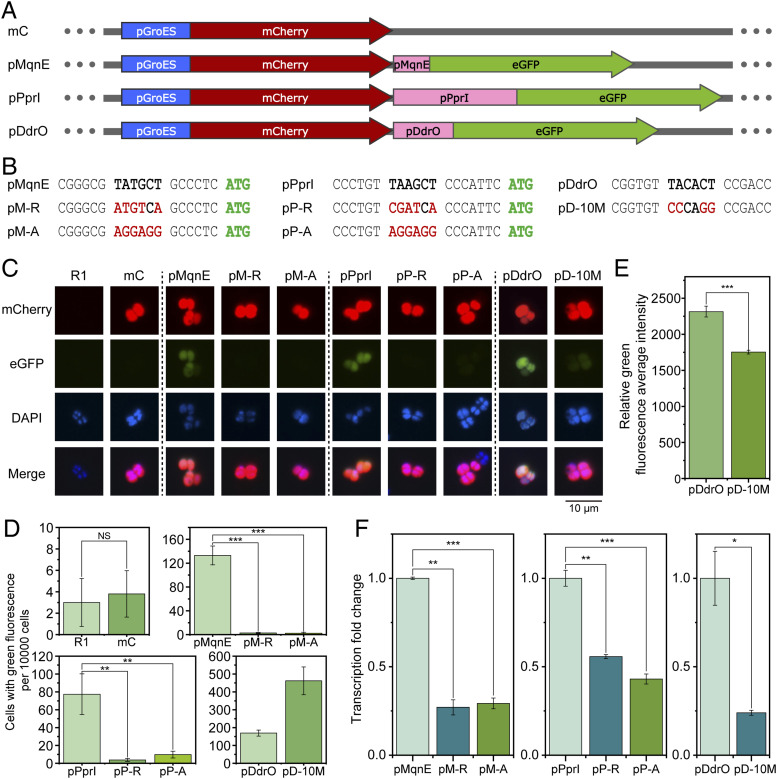 Figure 2
Figure 2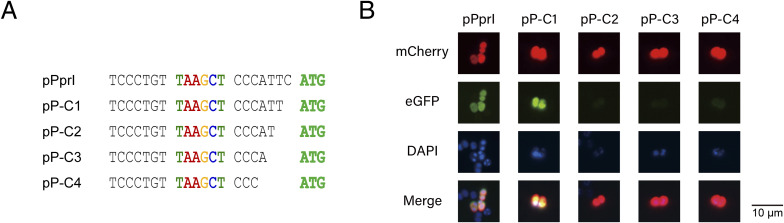 Figure S4
Figure S4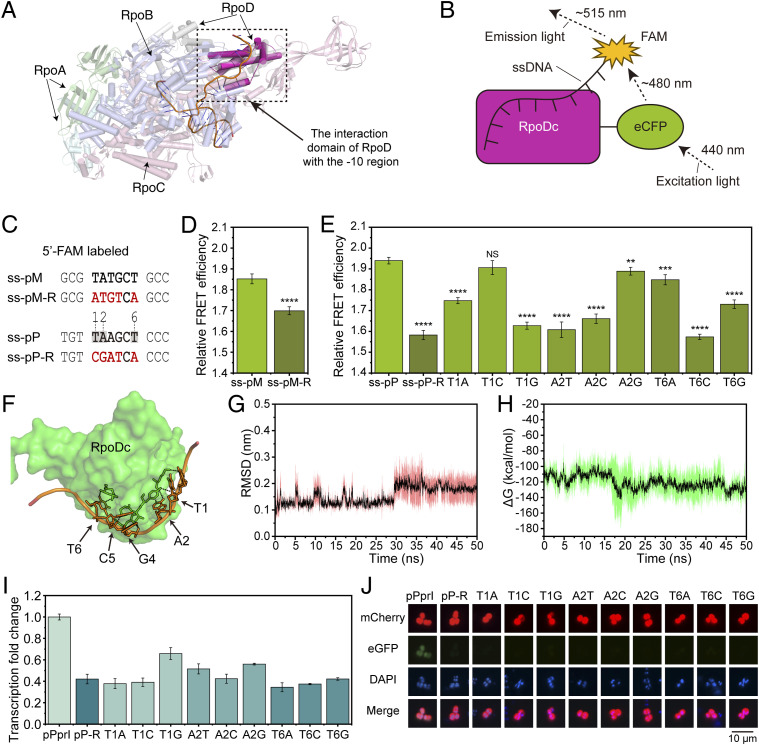 Figure 3
Figure 3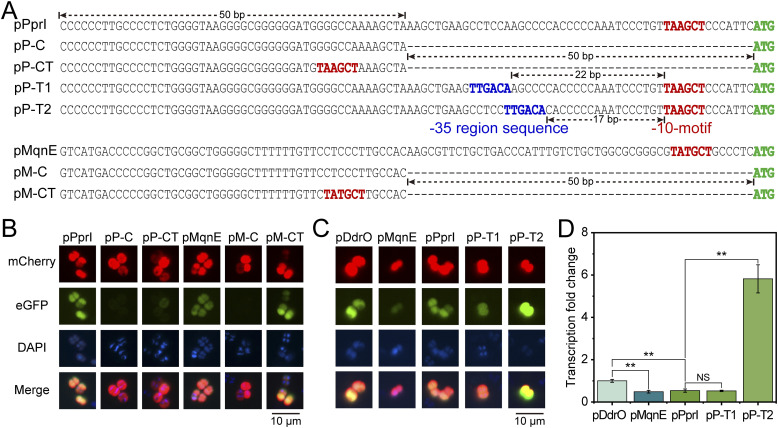 Figure 4
Figure 4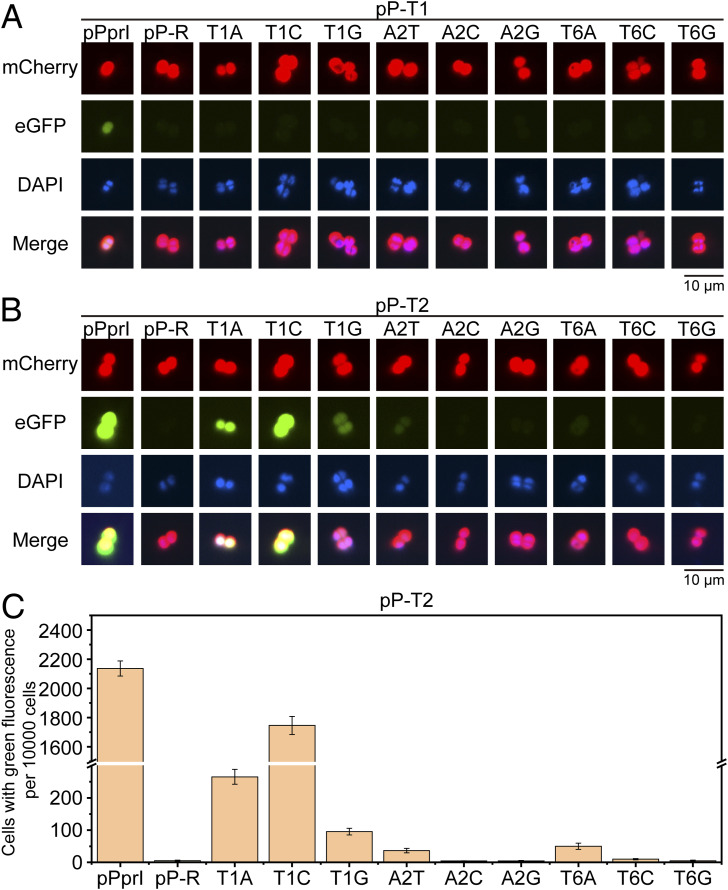 Figure S5
Figure S5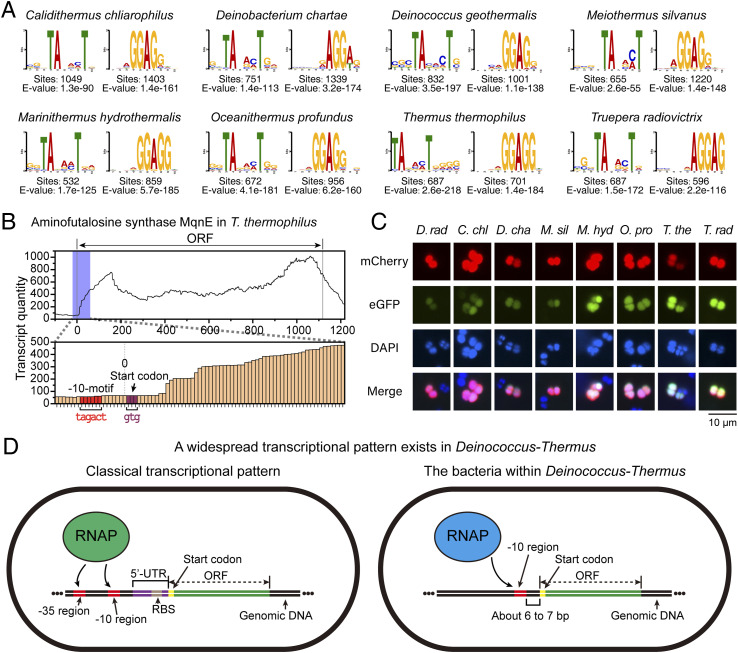 Figure 5
Figure 5 Figure S6
Figure S6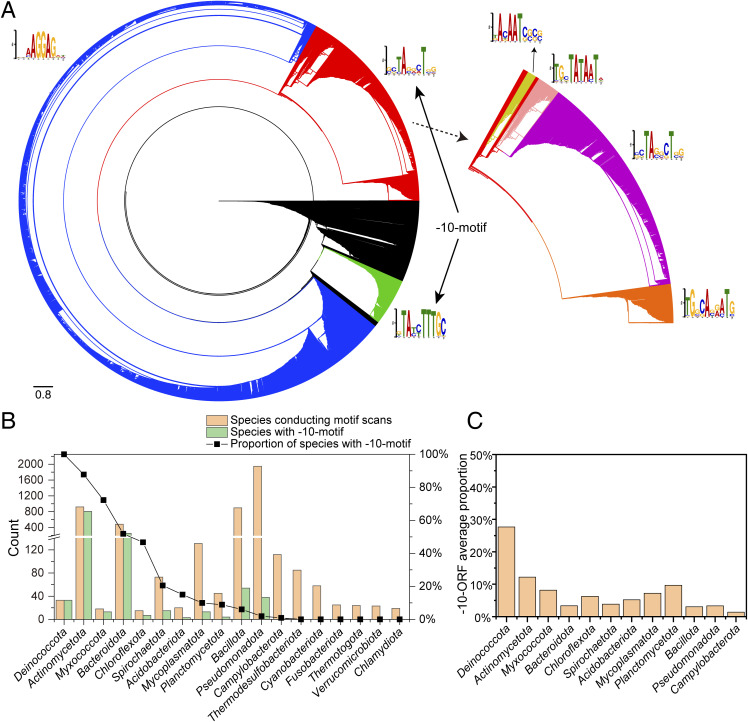 Figure S7
Figure S7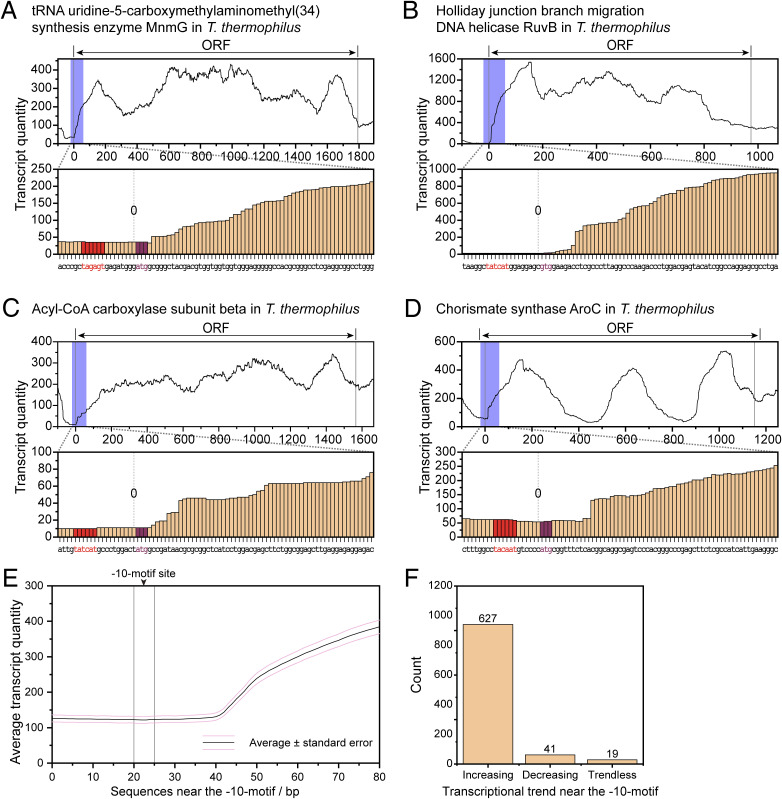 Figure S8
Figure S8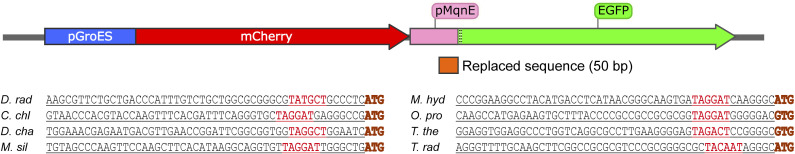 Figure S9
Figure S9 Figure S10
Figure S10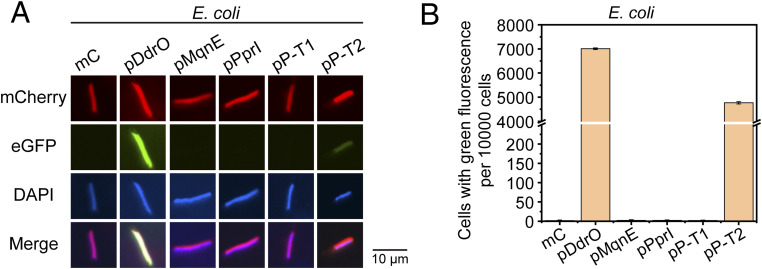 Figure S11
Figure S11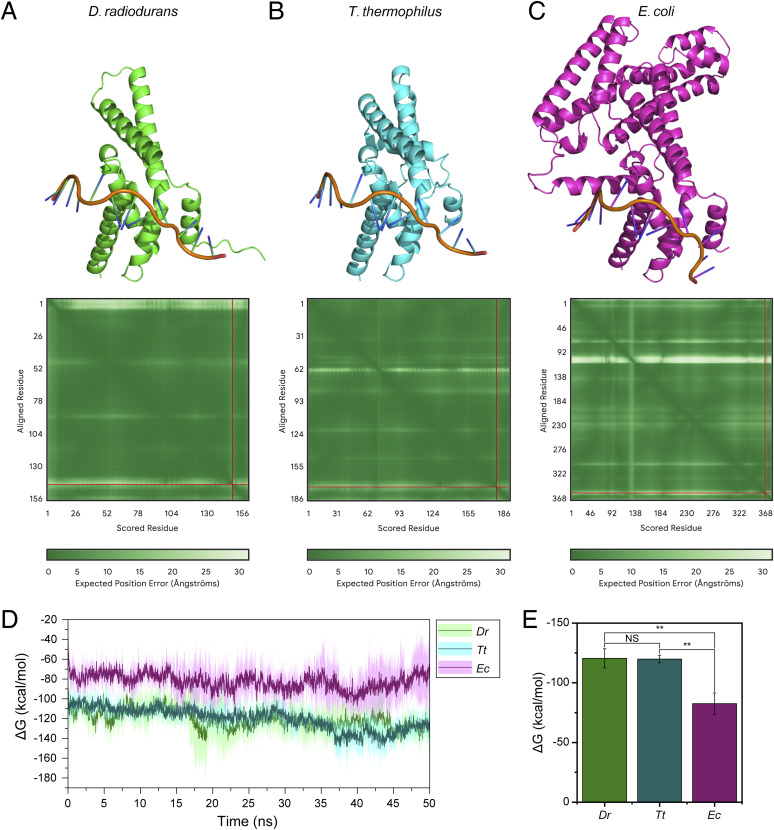 Figure S12
Figure S12 Figure S13
Figure S13 Figure S14
Figure S14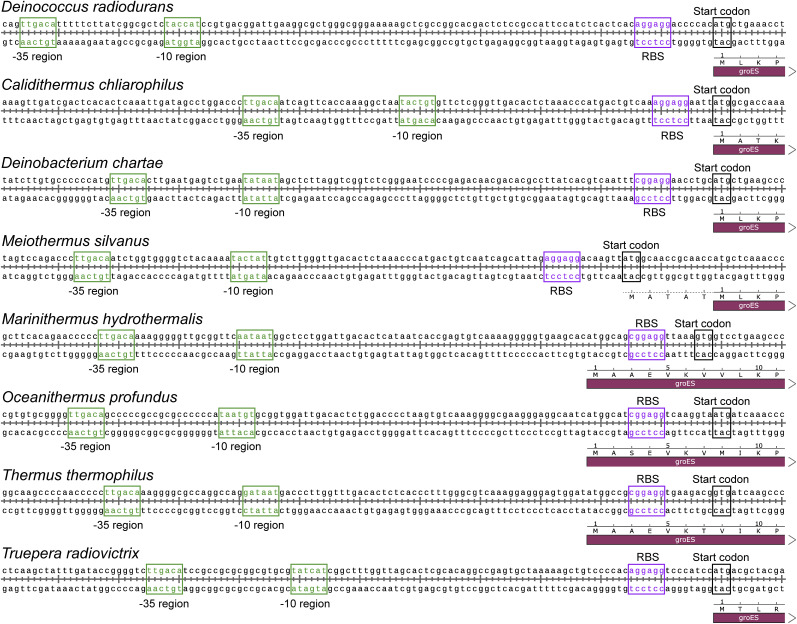 Figure S15
Figure S15- —MOST | National Natural Science Foundation of China (NSFC)http://dx.doi.org/10.13039/501100001809
- —MOST | National Natural Science Foundation of China (NSFC)http://dx.doi.org/10.13039/501100001809
- —MOST | NSFC | NSFC-Zhejiang Joint Fund | 浙江省科学技术厅 | Natural Science Foundation of Zhejiang Province (ZJNSF)http://dx.doi.org/10.13039/501100004731
- —MOST | NSFC | NSFC-Zhejiang Joint Fund | 浙江省科学技术厅 | Natural Science Foundation of Zhejiang Province (ZJNSF)http://dx.doi.org/10.13039/501100004731
- —MOST | NSFC | NSFC-Zhejiang Joint Fund | 浙江省科学技术厅 | Natural Science Foundation of Zhejiang Province (ZJNSF)http://dx.doi.org/10.13039/501100004731
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA and protein synthesis mechanisms · Bacterial Genetics and Biotechnology · Genomics and Phylogenetic Studies
Introduction
Gene transcription and translation are pivotal in the life cycle of an organism, governing the conversion of genetic information into functional proteins. Central to these processes are the promoter and the ribosome binding site (RBS), which are strategically positioned upstream of an ORF. RNA polymerase (RNAP) recognizes and binds to the promoter, thereby initiating transcription and resulting in the synthesis of mRNA. After transcription, the ribosome identifies the RBS on the mRNA molecule, marking the initiation for translation and ultimately leading to the production of polypeptides.
Prokaryotic promoter research has largely been centered on Escherichia coli. In 1974, Schaller’s work elucidated the characteristics of the RNAP binding site (Schaller et al, 1975). Pribnow’s subsequent work identified the sequence 5′-TATPuATG-3′ as critical for the formation of a stable complex between the promoter and RNAP (Pribnow 1975), which contributed to the nomenclature of the −10 region as the Pribnow box. As research advanced, Takanami and colleagues identified a sequence ∼35 base pairs upstream of the transcription initiation site that is crucial for transcription recognition (Takanami et al, 1976). In E. coli, sequence alignment upstream of the transcription start site has revealed that the −35 region typically matches 5′-TTGACA-3′, and the −10 region aligns with 5′-TATAAT-3′ (Hawley & McClure 1983; Harley & Reynolds 1987). Extensive research has subsequently confirmed that the sigma factor (RpoD) plays a key role in interacting with the −35 and −10 regions, thereby facilitating the accurate initiation of transcription (Simpson 1979; Morett & Buck 1989; Bowers & Dombroski 1999).
In the exploration of the RBS, initial theories posited that translation initiation was dependent on the recognition of specific nucleotide sequences or the identification of secondary or tertiary structures in the mRNA (Hindley & Staples 1969; Steitz 1969). However, the discovery of the 3′-terminal sequence of 16S rRNA and its complementarity to a sequence upstream of the initiation codon implied that ribosomes recognize the initiation site through base pairing with the 5′-UTR of the mRNA (Shine & Dalgarno 1974; Steitz & Jakes 1975). Subsequent experimental validations confirmed that the ribosome recognition sequence on mRNA correlates with the 5′-CCUCCU-3′ sequence at the end of 16S rRNA (Hui & de Boer 1987; Jacob et al, 1987). This led to the identification of the classical RBS as the sequence 5′-AGGAGG-3′ or 5′-GGAGG-3′, which is commonly referred to as the Shine–Dalgarno (SD) sequence.
The field of transcription and translation research continues to yield new insights that mRNAs lacking or possessing very short 5′-UTRs can still be normally translated (Shean & Gottesman 1992). These mRNAs, referred to as leaderless mRNAs, are widely distributed among bacteria, fungi, and archaea (Moll et al, 2002). This interesting phenomenon indicates that the upstream regions of these ORFs contain solely the promoter without RBS. Furthermore, studies have revealed a variety of promoter motifs in bacterial genomes (Ponnambalam et al, 1986; Keilty & Rosenberg 1987; Morett & Buck 1989; Gleghorn et al, 2008). The expansion of genomic databases has enabled bioinformatics analyses showing that sequences upstream of ORFs reveal a diversity in base composition across different bacterial species (Omotajo et al, 2015; Nakagawa et al, 2017). This suggests that the large amount of information contained in the DNA sequence upstream of the ORFs requires further investigation into its possible connection to the promoter or RBS. Given the intricacy of the upstream ORF sequences, it may be necessary to reconsider the definition/constitution of promoters (Mejía-Almonte et al, 2020).
Promoter research within the Deinococcus-Thermus (*D-*T) phylum has been largely centered on the genus Thermus, with a focus on understanding the role of transcriptional motifs (Feklistov et al, 2006; Sevostyanova et al, 2007; Miropolskaya et al, 2018) and their interactions with RNAP (Zhang et al, 2012; Weixlbaumer et al, 2013; Bae et al, 2015), as well as comparisons of promoter–RNAP interaction characters with E. coli (Mekler et al, 2012; Miropolskaya et al, 2012). Despite some efforts to summarize known promoter sequences (Sevostyanova et al, 2007), a comprehensive genomic overview of promoter organization within this phylum remains absent. In contrast, a research on Deinococcus promoters has been more piecemeal, with studies failing to converge on a unified set of distinct patterns (Chen et al, 2019). Nevertheless, analysis of the sequences upstream of ORFs in Deinococcus has uncovered a motif (5′-TANNNT-3′) that closely resembles the classical −10 region in E. coli, suggesting a potential role of this motif in transcription (Zheng et al, 2011; de Groot et al, 2014).
Consistent with a previous study, our analysis of the DNA sequence upstream of all ORFs in Deinococcus radiodurans (D. radiodurans) also revealed the conserved motif with the sequence 5′-TANNNT-3′, designated as the −10-motif. This motif displays a high degree of conservation within Deinococcus, suggesting its crucial role in gene expression. Through a series of experiments, we have demonstrated that the −10-motif is indeed associated with transcription and plays a classical −10 region role in RNAP recognition. Mutations at the conserved sites of the −10-motif led to abnormal gene expression. Our findings suggest that approximately one third of genes in D. radiodurans are transcribed as leaderless mRNA, raising concerns about the accuracy of some gene annotations. Notably, when a −10-motif appears at an appropriate position on a sequence, it can function as a promoter sequence in D. radiodurans. In addition, the presence of the −35 region can significantly enhance transcriptional expression levels. What’s more, our sequence alignment has revealed that the −10-motif is a widespread feature across various species within the D-T phylum, and the function assays demonstrated a conserved role of −10-motif in transcriptional regulation. Thus, we posit that the −10-motif is a significant promoter pattern in the D-T phylum.
In the classical expression model primarily studied in E. coli, gene transcription relies on promoters containing −35 and −10 regions to produce mRNA with 5′-UTR; and subsequent translation depends on the RBS located within this 5′-UTR. In contrast, our research reveals that a significant number of genes in the D-T phylum use transcription relying on a −10 region immediately upstream of the ORF to produce leaderless mRNA for protein expression. This pattern is markedly different from the classical expression model.
Results
−10-motif forms a nonclassical expression pattern in D. radiodurans
Through DNA sequence analysis on sequences 20 bp upstream of D. radiodurans ORFs using MEME software (Bailey et al, 2015), two primary motifs were identified (Fig 1A), consistent with findings in Deinococcus deserti (de Groot et al, 2014). One motif (−10-motif) presents in the form of TANNNT, similar to the classical −10 region of promoter, whereas the other (RBS-motif) presents in the form of GGAG, similar to classical RBS sequences. These two predicted motifs, although similar in sequence to known functional sequences, still require further validation for their specific functions. Because the RBS-motif not only shares similar features in sequence with classical RBS sequences but also appears a few bp upstream of the ORF, matching the positional characteristics of classical RBS, we infer that it is the RBS sequence in D. radiodurans. In contrast, −10-motif occupies a position similar to that of the RBS, but exhibits a distinct sequence. It is similar to the −10 region of classical promoters and is most frequently found in the form of 5′-TACACT-3′ (Fig S1A). Notably, the −10-motif predominantly occurs ∼6–7 bp upstream of the ORF (Fig S1B and C), present in nearly one-third of the D. radiodurans genes (Fig 1A), and does not show a bias toward specific pathways (Fig S1D), which is consistent with previous findings (de Groot et al, 2014).
Performance of motifs upstream of the ORFs and RNA transcription in D. radiodurans.(A) Two motifs were identified by scanning the 20-bp sequence upstream of the ORFs (the total number is 3,146) in the D. radiodurans genome. The left −10-motif closely resembles the −10 region of a classical promoter, whereas the right ribosome binding site (RBS)-motif corresponds to a classical RBS sequence. (B) Classical transcription and translation patterns in D. radiodurans, exemplified by groES and ddrO. The −35 region, −10 region, and RBS are highlighted in green, blue, and purple, respectively. (C, D, E, F) RNA transcription statistics of genome sequences. (C, D) Transcriptome data reveal that following the promoters (highlighted in green and blue) of GroES (C) and DdrO (D), a clear elevation in transcription levels of related sequences is observed, aligning with the expected function of promoters. (E, F) Similarly, after the −10-motif (highlighted in red) of MqnE (E) and PprI (F), a notable increase in transcription levels is evident, suggesting that the function of −10-motif parallels that of a promoter. (The bar chart below is created by enlarging the blue section from the line graph above. The annotated start codon of the ORF is indicated in dark purple, whereas the revised start codon is marked in blue).
Distribution of −10-motif in Deinococcus radiodurans.(A) −10-motif typically occurs ∼6–7 bp upstream of the ORF. (B) 10 most prevalent −10-motif variants in D. radiodurans. (C) Positioning of the −10-motif in relation to the ORF. (D) Comparison of the classification of genes containing −10-motif with the classification of genes in the entire genome. One letter code descriptions: [D] cell cycle control, cell division, chromosome partitioning; [M] cell wall/membrane/envelope biogenesis; [N] cell motility; [O] posttranslational modification, protein turnover, chaperones; [T] signal transduction mechanisms; [U] intracellular trafficking, secretion, and vesicular transport; [V] defense mechanisms; [W] extracellular structures; [Y] nuclear structure; [Z] cytoskeleton; [A] RNA processing and modification; [B] chromatin structure and dynamics; [J] translation, ribosomal structure, and biogenesis; [K] transcription; [L] replication, recombination, and repair; [C] energy production and conversion; [E] amino acid transport and metabolism; [F] nucleotide transport and metabolism; [G] carbohydrate transport and metabolism; [H] coenzyme transport and metabolism; [I] lipid transport and metabolism; [P] inorganic ion transport and metabolism; [Q] secondary metabolite biosynthesis, transport, and catabolism; [R] general function prediction only; [S] function unknown.
Notably, the −10-motif is present and highly conserved across various species within the Deinococcus genus, with the first T, second A, and sixth T remaining nearly invariant (Fig S2A–E). Interestingly, some −10-motifs appear within the reading frame or far upstream, which suggest inaccuracies in previous reading frame annotation, such as the −10-motif of the pprI (also known as irrE) (Fig S2E), which has been discussed by a previous study (Ludanyi et al, 2014). With the −10-motif ahead of the ORF, the transcript is assumed to lack 5′-UTR, which was identified as leaderless mRNA by previous studies (Zheng et al, 2011; de Groot et al, 2014).
−10-motif conservation in the Deinococcus genus homologous proteins.(A, B, C, D, E) −10-motif is marked in color; the start codon in brown indicates the start codon for the annotated ORF. The start codon in blue represents a corrected codon not used as a start codon by the original annotation, yet it aligns with the amino acid translation frame, hinting at a possible annotation discrepancy. The start codon in black also is not employed as a start codon by the initial annotation, nor does it align with the translation frame of the original annotation. A frameshift in the amino acid sequence suggests the possibility of genomic sequencing inaccuracies. (E) Within the ORF of PprI from D. radiodurans, −10-motif (highlighted in red) is found immediately followed by a start codon GTG (highlighted in blue). In the AlphaFold-predicted structure of PprI (AlphaFold DB: Q9RXY7), a long-disordered peptide chain is present at the N terminus (highlighted in purple). The −10-motif of pprI followed by the start codon is also observed in some other species within the Deinococcus genus, and these corrected start codons can match the original reading frame.
In D. radiodurans, some proteins such as GroES and DdrO are expressed through the traditional transcription and translation pathways, using classical promoter sequences that are followed by RBS sequences (Fig 1B). To determine whether there is any regulation preference of −10-motifs adjacent to ORFs versus classical promoters, we performed enrichment analysis on the transcriptome data of D. radiodurans (SRX2731648) to observe RNA transcription levels near the ORF. Analysis of RNA transcription levels reveals a substantial elevation following the classical promoter’s −10 region (Fig 1C and D), consistent with the known function of promoter in initiating transcription. Similarly, we observed a notable increase in transcription levels following the −10-motif (Figs 1E and F and S3A–F), suggesting that the −10-motif could be a regulatory element like classical promoters, which controls gene transcription in D. radiodurans.
RNA transcription levels of −10-motif–containing proteins in D. radiodurans.(A, B, C, D) Randomly selected examples. The blue box in the curve graph represents the transcription levels spanning from −20 to +60 bp relative to the ORF, which is further detailed in the bar graph below. In the bar graph, the −10-motif is denoted in red, the originally annotated start codon is in purple, and the inferred correct start codon is highlighted in blue. (E) Average transcriptional trends near the −10-motif. (F) Counting results of transcriptional trends.
−10-motif is important for gene expression
Previous sequence analysis has shown that many species have their ORFs closely followed by RBSs upstream (Omotajo et al, 2015; Nakagawa et al, 2017), and therefore, it can be inferred that the classical promoter −10 regions for these ORFs should be located dozens bp upstream. In contrast, the genomic location of −10-motif in D. radiodurans is very close to the ORF, which markedly differs from that of classical promoters, pointing out the possibility of the −10-motif acting as a RBS. To investigate the role of the −10-motif, we constructed a reporter plasmid using the upstream sequence of aminofutalosine synthase mqnE (pMqnE) and the modified upstream sequence of metalloprotease pprI (pPprI), followed by eGFP as a reporter gene (Fig 2A). The upstream sequence of groES (pGroES) and mCherry was fused for background referencing, and the upstream sequence of ddrO (pDdrO) coupled with eGFP for control (Fig 2A). We introduced random mutation into the −10-motif of pMqnE and pPprI, or substituted them with a classical RBS sequence. In addition, random mutation was also introduced into the −10 region of pDdrO (Fig 2B). The constructed plasmids were transformed into D. radiodurans R1 strain, and the expression of eGFP protein was monitored using fluorescence microscopy. As well as the mutations, the substitution of −10-motif with classical RBS also led to the loss of green fluorescence, indicating the different role of −10-motif compared with RBS. In contrast, mutation in the −10 region of pDdrO resulted only in a diminished fluorescence intensity, suggesting a different transcription mode between the −10-motif and classical −10/−35 promoter (Fig 2C). The fluorescent observations were further substantiated by cell flow cytometry data (Fig 2D and E). Using the transcription level of the mCherry protein as a baseline, we quantified the transcription level of eGFP through qPCR. The results indicate that occupation of the −10-motif by alternate sequences leads to a marked reduction in RNA levels, similar to the behavior observed in pDdrO with mutations in the −10 region (Fig 2F). Consequently, we affirmed the role of −10-motif as a promoter. Nevertheless, the −10-motif necessitates a specific proximity to the initiation codon, whereas an excessively close distance would lead to aberrant expression (Fig S4A and B).
*Impact of −10-motif substitution on protein expression.(A) Construction of four fluorescent reporter vectors, where mCherry served as a reference linked to the upstream sequence of the groES (pGroES), and eGFP acted as an indicator linked to the upstream sequences of the mqnE (pMqnE), pprI (pPprI, new ORF start position was used), and ddrO (pDdrO). (B) Replacement of key sites in the pMqnE, pPprI, and pDdrO, in which the −10-motif of pMqnE and pPprI was replaced with either a random sequence (pM-R and pP-R) or a classical ribosome binding site (pM-A and pP-A), and the −10 region of the DdrO promoter was substituted with a random sequence (pD-10M). (C) Expression of fluorescent proteins mCherry and eGFP in D. radiodurans. Results showed that the replacement of the −10-motif in MqnE and PprI resulted in the disappearance of eGFP fluorescence, whereas the replacement of the −10 region in DdrO promoter resulted in the reduced eGFP fluorescence. (D, E) Green fluorescence expression captured by flow cytometry. (F) Transcription level of eGFP was assessed relative to the WT sequences, revealing that the substitution of key sites resulted in decreased transcription levels. NS: P ≥ 0.05, *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001.
Impact of decreasing the −10-motif to start codon distance on eGFP expression.(A) Sequential base deletions from the start codon side to diminish the gap between the −10-motif and the start codon. (B) eGFP expression outcomes subsequent to the distance reduction.
The −10-motif functions as the classical −10 region
Previous research on the crystal structure of RNAP from Thermus thermophilus (PDB: 4G7H) has identified a crucial domain within RpoD that interacts with the −10 region. Given that the predicted structure of D. radiodurans’s RpoD aligns well with this domain and the −10-motif closely resembles the classical −10 region, we hypothesized that −10-motif may interact with the RpoD protein (Fig 3A). To test the interaction between the −10-motif and RpoD, we constructed a truncated RpoD (RpoDc) and linked it to the eCFP protein at the C terminus via a flexible peptide linker. Meanwhile, we labeled a 5′-FAM tag to the single-stranded DNA (ssDNA) containing the −10-motif (from pMqnE and pPprI), and assessed the interaction between −10-motif and RpoD through fluorescence resonance energy transfer (FRET) between FAM and eCFP (Fig 3B and C). Taking the intensity ratio of 515-nm emission light to 480-nm emission light as an indicator of FRET efficiency, we observed strong interaction between −10-motif and RpoD, whereas the mutated −10-motif exhibited a significant decrease in FRET efficiency (Fig 3D and E), suggesting that the −10-motif interacts with the RpoD and indeed functions as the −10 region. We further used AlphaFold to predict the complex structure of RpoDc with −10-motif, exhibiting multihydrogen bonds and salt bridges participating in the interaction (Fig 3F). The binary model was then validated by molecular dynamics simulations, with the root mean square deviation (RMSD < 0.2 nm) of the −10-motif showing the stability of the binding between RpoDc and −10-motif (Fig 3G). The calculated interaction free energy ΔG further demonstrated the high affinity between the protein and DNA (Fig 3H).
*Interactions between the −10-motif and RpoD’s corresponding domain, and the effect of point mutations on interaction and protein expression.(A) Interaction between RNA polymerase and the −10 region. The crystal structure of RNA polymerase was from T. thermophilus (annotated in light color, PDB: 4G7H), and the truncated RpoD (RpoDc) from D. radiodurans was predicted by AlphaFold (indicated in purple). (B) Design diagram of fluorescence resonance energy transfer (FRET) experiments, in which the C terminus of RpoDc is conjugated to eCFP and ssDNA is tagged with 5′-FAM, enabling FRET upon molecular interaction. (C) Four ssDNAs labeled with 5′-FAM were designed based on −10-motifs in pMqnE and pPprI. The −10-motif of pPprI was modified to introduce point mutations at the first, second, and sixth nucleotide positions, respectively. (D, E) FRET results of four ssDNAs and ss-pP point mutations with RpoDc. (F) Interaction structure of RpoDc with −10-motif predicted by AlphaFold. Hydrogen bonds are indicated by dashed lines. The numbered bases belong to the −10-motif, with the exception of A2 and T6, where all other bases interact via the phosphate backbone. (G, H) Results from molecular dynamics simulations of the predicted structure: the root mean square deviation of the −10-motif (sequence: TAAGCT) (G) and the interaction free energy ΔG between protein and DNA (H). (I, J) Transcription-level alterations (I) and expression results (J) of eGFP in D. radiodurans after point mutations in the −10-motif of PprI. NS: P ≥ 0.05, *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001.
Except for T1C, the substitution of bases at positions 1, 2, and 6 with any other bases decreased the interaction significantly (Fig 3E), emphasizing the participation of the bases in interacting with RpoD. Meanwhile, in vivo mutations of these bases significantly reduced the transcription levels of eGFP compared with the WT (Fig 3I), leading to the decreased expression of the reporter eGFP (Fig 3J), suggesting that these three bases are extremely important for gene transcription, as well as the interaction with RpoD.
Sequences containing the −10-motif can serve as promoters
Notably, the upstream sequences of most −10-motif genes do not exhibit the presence of a classical −35 region. To test whether there exists a sequence similar to the −35 region to aid −10-motif in normal gene expression, we excised the 50-bp segment upstream of the start codon and reinserted the −10-motif at the new position (Fig 4A). Analysis of fluorescent protein expression revealed that the 50-bp truncation eliminated the normal expression of fluorescent proteins, which could be restored by reintroducing the −10-motif (Fig 4B). This indicates that the presence of a −10-motif in the sequence is sufficient to function as a promoter in D. radiodurans, and also suggests that the RNAP in D. radiodurans has weak dependence on sequences other than the −10-motif. However, variations in fluorescence intensity upon reintroduction imply that the sequences adjacent to the −10-motif still have an impact on transcription strength (Fig 4B).
*Dependency of the −10-motif on other sequences.(A) pPprI and pMqnE were truncated by 50 bp (pP-C, pM-C), and the −10-motif was added to the truncated sequences (pP-CT, pM-CT). The −35 region sequences were introduced at 22 bp (pP-T1) and 17 bp (pP-T2) upstream of the −10-motif in pPprI, respectively. (B) Impact of a 50-bp truncation adjacent to the start codon and the restoration of the −10-motif on the expression of fluorescence. (C, D) Effects of −35 region sequence on fluorescence expression. NS: P ≥ 0.05, *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001.
Generally, classical promoter in organisms like E. coli comprises the −10 region and the −35 region, both of which contribute to the recognition by sigma factor (Zuo & Steitz 2015). Through transcriptional and translational levels, we have demonstrated that the −10-motif upstream of ORFs functions as a classical −10 region in D. radiodurans. Furthermore, simply introducing the −10-motif sequence alone is sufficient to enable a segment of DNA to be recognized by RpoD (sigma factor) and function as a promoter (Fig 4A and B). Because both the −35 region and −10 region are required in classical promoters, it is possible that adding a −35 region sequence upstream of the −10-motif in D. radiodurans might stabilize or assist in protein expression. To evaluate the impact of the −35 region on the −10-motif, we integrated the −35 region into pPprI (followed by the eGFP gene) to assess its effect on fluorescence expression (Fig 4C and D). Compared with pDdrO, pMqnE and pPprI led to markedly lower transcription levels (Fig 4D). However, with the −35 region positioned appropriately upstream (17 bp) of the −10-motif (pP-T2), the promoter significantly enhanced gene transcription and expression levels, even for the variants of the −10-motif (Figs 4C and D and S5B and C). When the −35 region is positioned inappropriately upstream (22 bp) of the −10-motif (pP-T1), the enhancement was gone (Figs 4C and D and S5A). In fact, existing studies have shown that the optimal distance between the −35 region and the −10 region is 17 ± 1 bp in E. coli (Aoyama et al, 1983) and B. subtilis (Yeh et al, 2011), which is consistent with our results.
Fluorescence expression results under point mutations of the −10-motif when the −35 region appears at different positions.(A) eGFP expression outcomes when the −35 region appears at 22 bp upstream of the −10-motif. (B, C) eGFP expression outcomes when the −35 region appears at 17 bp upstream of the −10-motif.
Ubiquity and functional consistency of the −10-motif in the D-T phylum
We also analyzed the upstream motifs of ORFs in species within the D-T phylum (also known as Deinococcota) and observed that the results are similar to those in D. radiodurans, with the clear presence of the −10-motif and RBS-motif (Figs 5A and S6). Furthermore, we conducted an analysis of upstream sequences of ORFs in bacterial species across the entire domain, where the predominant motifs identified were again the −10-motif and RBS-motif (Fig S7A). Interestingly, all analyzed species within the D-T phylum exhibited the presence of the −10-motif (Fig S7B), with the average proportion of ORFs carrying the −10-motif reaching ∼30% (Fig S7C), making it the most prominent among all phyla studied.
Distribution and performance of −10-motif across D-T phylum.(A) Results of motif discovery within the upstream sequences of ORFs across various species within the D-T phylum. (B) Transcription-level statistics for the mqnE gene in T. thermophilus indicating a marked increase in transcription subsequent to the −10-motif. (C) Outcomes of substituting the 50-bp sequence proximal to the start codon in the mqnE ORF upstream sequence with the corresponding 50-bp sequence from other D-T phylum species. These sequences enabled the normal expression of eGFP in D. radiodurans, although there might be differences in fluorescence intensity. (D) Widespread transcriptional pattern in the D-T phylum.
Motif scanning results for species in the D-T phylum.
Motif scanning results for species in the bacterial domain.(A) Clustering results of the scanned motifs mainly show two categories: ribosome binding site sequences and −10-motifs, with −10-motifs further divided into five forms. (B) Proportion of −10-motif carrying species (−10-species) under common phyla, with the proportion in the D-T phylum (Deinococcota) being 100%. (C) Average proportion of −10-motif carrying ORFs (−10-ORFs) in −10-species under common phyla. Among them, the average proportion of −10-ORFs in the D-T phylum (Deinococcota) is the highest.
Taking T. thermophilus as an example, the transcription levels elevate following the −10-motif, which is similar to D. radiodurans, suggesting that the −10-motif across the D-T phylum uniformly acts as a promoter sequence (Figs 5B and S8A–F). Besides, the substitution of promoter sequence with 50-bp sequences upstream of the ORFs from other species before the eGFP-encoded sequence led to the normal expression of eGFP in D. radiodurans (Figs 5C and S9), demonstrating the universality of the −10-motif in the D-T phylum. In addition, high resemblance in sequence alignment of the RpoD protein also suggested the similar interaction between −10-motif and RpoD (sigma factor) throughout the D-T phylum (Fig S10).
RNA transcription-level analysis of −10-motif–containing proteins in T. thermophilus.(A, B, C, D) Randomly selected examples. The blue box in the curve graph signifies the transcription levels from −20 to +60 bp of the ORF, which is expanded in the bar graph below. In the bar graph, the −10-motif is indicated in red, and the annotated start codon is marked in purple. (E) Average transcriptional trends near the −10-motif. (F) Counting results of transcriptional trends.
50-bp sequence proximal to the start codon in the MqnE ORF upstream sequence was substituted with the corresponding 50-bp sequence from other D-T phylum species.
Amino acid sequence alignment of the RpoD protein in the D-T phylum.
Discussion
Sequences upstream of the ORFs are significant for normal transcription and translation of genes. In prokaryotes, the classical promoter for the recognition and binding of RNAP consists of the −35 region and −10 region. After transcription, the resulting mRNA typically contains 5′-UTR, where the RBS, related to translation, is located upstream of the start codon. However, researchers have identified leaderless mRNAs lacking or possessing very short 5′-UTRs, which can still be normally translated. Studies suggest that the initiation of leaderless mRNA translation is triggered by the interaction between the 70S particle and the start codon (O’Donnell & Janssen 2002; Udagawa et al, 2004; Beck et al, 2016), which is different from the conventional translation process that relies on initiation factors (Cummings & Hershey 1994; Tedin et al, 1999; Yamamoto et al, 2016).
This article reveals that apart from the RBS-motif, a conserved motif (−10-motif) similar to the classical −10 region is located ahead of the start codon for almost one-third ORFs in D. radiodurans, functioning as −10 region of classical promoter in transcription and resulting in disappeared or shortened 5′-UTR for the following leaderless mRNA.
Because the −10-motif is located ahead of the start codon like classical RBS, we designed experiments to investigate whether it functions like RBS. The results turned out that the −10-motif cannot be replaced by a classical RBS sequence, and the interaction between −10-motif and RpoD (sigma factor) further indicated that the −10-motif acts like −10 region of the classical promoter. We further confirmed that the 1/2/6 bases of the −10-motif are irreplaceable, consistent with structural findings of RpoD interacting with the −10 region (Feklistov & Darst 2011; Zhang et al, 2012; Zuo & Steitz 2015), confirming the −10-motif is a −10 region. Our experiments also demonstrated that in Deinococcus, a sequence with the −10-motif introduced at an appropriate position can function as a promoter. In contrast, different to that in Deinococcus, the introduction of −10-motif–containing plasmids constructed above into E. coli does not achieve eGFP expression, suggesting the disparate activity of RpoD in other species (Fig S11A and B). Meanwhile, free energy calculations also indicated that RpoD from E. coli has a weaker binding affinity to the −10 region (−10-motif) compared with D. radiodurans and T. thermophilus (Fig S12A–E). Besides, the diminished expression restored after the addition of −35 region into the proper position before the −10-motif (Fig S11A and B), confirming that RpoD from E. coli requires both the −10 and −35 region of the classical promoter to complete its mission of transcription initiation. However, for the leaderless genes in Deinococcus, the addition of the appropriately positioned −35 region led to enhancement of gene expression levels. In addition, −10-motifs appearing in different sequences exhibit varying expression strengths, and we speculate that this might be influenced by their adjacent sequences, which has also been discovered in Thermus aquaticus (Feklistov et al, 2006). Furthermore, the binding motifs of some transcriptional regulators, such as DdrO binding site characterized by previous studies (Makarova et al, 2007; Wang et al, 2015), may overlap with the −10-motif or appear nearby (Fig S13), thereby regulating expression strength.
Expression results of fluorescent proteins in E. coli with different upstream sequences of ORFs.(A, B) Fluorescence microscopy images (A) and cell flow cytometry results (B) show that besides pDdrO with the classical form, only pP-T2 carrying the −35 region sequence at an appropriate position can also function in E. coli.
*Results of interaction structure prediction and molecular dynamics simulations for truncated RpoD proteins from E. coli, D. radiodurans, and T. thermophilus with the −10 region (−10-motif).(A, B, C) Complex structures predicted by AlphaFold3. (D) Binding free energy (ΔG) results during the molecular dynamics simulation process. (E) Statistical results and significance tests of the binding free energy (ΔG). NS: P ≥ 0.05, *P < 0.05, **P < 0.01, ***P < 0.001, ***P < 0.0001.
In D. radiodurans, the binding site of DdrO occupies a sequence near the −10-motif, thereby affecting gene expression.
ORFs were used to be characterized through the identification of the start codon. However, in Deinococcus, proteins such as PprI, LspA, MurD, and RecF, which are encoded after the −10-motif, exhibit disordered peptide chains at the N terminus in the predicted or crystal structures (Figs S2E and S14A–C). The peptide chains float around the periphery of the tight tertiary structure, suggesting its possible nonfunctionality and the incorrect annotation for previous leaderless genes. The enzymatic activity of PprI without the peptide was intact (Lu et al, 2019, 2024), further confirming the annotation error. Thus, the identification of the −10-motif as the promoter also enabled us to reannotate the leaderless genes.
Some proteins encompass −10-motifs within their ORFs or proximal to their start codons.(A, B, C) Protein examples include LspA (A), MurD (B), and RecF (C) in D. radiodurans. −10-motifs are highlighted in red, and the predicted start codons are indicated in blue. The structures of LspA (AlphaFold DB: Q9RRU7), MurD (AlphaFold DB: Q9RRJ4), and RecF (PDB: 2O5V) were sourced from the AlphaFold database and Protein Data Bank, respectively. Protein structures exhibit irregular coils at their N termini, located upstream of the predicted start codons, which are marked in purple. −10-motifs show high conservation across related species, with the following annotations: −10-motifs are colored, original annotated start codons are in brown, start codons with suspected annotation errors are underlined, and presumed correct start codons are in blue.
In addition, we found that the −10-motif is not only prevalent in Deinococcus but also observed across various species within the D-T phylum (Figs 5A and S6). The conservation of the −10-motif and its associated transcription processes (Figs 5B and S8A–F) are consistent with those found in Deinococcus (Figs 1E and F and S3A–F). Moreover, the upstream sequences of ORFs containing the −10-motif from these species are fully functional within D. radiodurans, suggesting a uniform role of the −10-motif as a promoter within the D-T phylum (Fig 5C and D). Beyond the −10-motif, classical gene expression paradigms also exist within the D-T phylum (Fig S15).
Classical gene expression sequences in the D-T phylum.The −35 and −10 regions of the promoter are highlighted in green. Ribosome binding sites are indicated in purple. Inferred start codons are outlined with black boxes.
There is speculation that leaderless genes represent an ancient mode of gene expression (Zheng et al, 2011; Beck & Moll 2018), and bacteria within the D-T phylum are considered to be ancient (Zheng et al, 2011; Coleman et al, 2021). Besides, many species within the D-T phylum are extremophiles, exhibiting heat, desiccation, and radiation resistance. Therefore, whether the −10-motif expression pattern is simply a remnant of an ancient expression mode in the course of evolution or contributes to the survival in the extreme environment remains a question.
In summary, species within the D-T phylum, exemplified by D. radiodurans, possess a substantial number of genes that use the −10-motif as the −10 region of a promoter for the expression of leaderless mRNA through RNAP recruitment (Fig 5D). A sequence with a −10-motif at the proper site is sufficient to ensure gene transcription in the D-T phylum. And the distinct preference of −10 sequence and −35 sequence among RNAPs from different species is worth further study.
Materials and Methods
Bacterial genomes and sequence analysis
Initially, the complete reference genome file (GCF_020546685.1) of D. radiodurans (Repar et al, 2021) was downloaded from NCBI on 30 March 2023. A 20-bp DNA sequence upstream of each annotated ORF was extracted and encapsulated into a FASTA file. MEME software (Bailey & Elkan 1994; Bailey et al, 2015) was then used to scan for potential hidden motifs within these sequences (command: meme upORFseqs.txt -dna -w 6 -mod zoops -nmotifs 2 -brief 50,000 -p 1). The scan results are shown in Table S1. The same procedure was followed for D-T phylum species listed in Table S2 (command: meme upORFseqs.txt -dna -w 10 -mod zoops -nmotifs 2 -brief 50,000 -p 1).
Table S1. MEME scan results of upstream sequences of ORFs in the D. radiodurans genome.
Table S2. D-T phylum reference genomes used in this article and MEME scan results.
Furthermore, for the identification of the potential −10-motif, the 450-bp sequence starting from 300 bp upstream of each ORF in D. radiodurans was extracted, and FIMO software (Bailey et al, 2015) was used to scan for possible −10-motifs (command: fimo --thresh 0.015 --norc meme_out/meme.html upORFseqs_300_150.txt). Suitable sequence information for statistical analysis was then filtered out, and the filtered results are shown in Table S3. Amino acid sequences of all ORFs in D. radiodurans and ORFs carrying −10-motif were further clustered using EggNOG (Huerta-Cepas et al, 2019), with detailed results shown in Tables S4 and S5, respectively.
Table S3. Fimo results of potential −10-motifs.
Table S4. Clustering results of all ORFs in the D. radiodurans genome.
Table S5. Clustering results of ORFs containing the −10-motif.
For motif scanning in the bacterial domain, reference genomes of species with complete sequencing were downloaded from NCBI on 10 December 2024, and species that were not clearly classified or symbiotic were excluded. The species information used is listed in Table S6. Motif scanning was then conducted using MEME software (command: meme upORFseqs.txt -dna -w 10 -mod zoops -nmotifs 3 -brief 50,000 -p 1) for 20-bp upstream ORFs (results are shown in Table S7). The clustering analysis of high-confidence motifs was performed using the motifStack package in R (Ou et al, 2018).
Table S6. Genomic information of bacterial species conducting motif scanning.
Table S7. Results of motif scanning in bacterial species.
Strains, culture media, primers, and reagents
The E. coli strains used in the study were E. coli Trans5a and E. coli BL21(DE3), both acquired from TransGen Biotech Co., Ltd. The strains used (D. radiodurans R1 and T. thermophilus HB8) were both sourced from the laboratory stock. Primers and ssDNA substrates used in the study (Table S8) were synthesized by Tsingke Biotech Co., Ltd. Unless indicated otherwise, all chemical reagents used in the study were obtained from Sangon Biotech (Shanghai) Co., Ltd. The culture medium for E. coli was Luria–Bertani (LB) medium (1% tryptone, 1% NaCl, 0.5% yeast extract), with solid media supplemented with 1.5% agar. The final concentrations of kanamycin and ampicillin used were 40 and 50 μg/ml, respectively. For D. radiodurans and T. thermophilus, the culture medium was TGY medium (0.5% tryptone, 0.1% glucose, 0.3% yeast extract), with solid media supplemented with 1.5% agar. The final concentration of chloramphenicol in the antibiotic selection medium for D. radiodurans was set at 4 μg/ml.
Table S8. Primers and ssDNA substrates used in the article.
The construction of the vectors
The plasmids used in this study are listed in Table S9. The PCR system employed in the article was Tks Gflex DNA Polymerase from Takara Biomedical Technology (Beijing) Co., Ltd. For the construction of the expression vector 28a-eCFP, the pET-28a(+) plasmid was linearized using primers pet-f and pet-r, followed by the linearization of ecfp using primers fp-pet-f and fp-pet-r. Subsequently, linearized plasmid and ecfp were treated with Exnase II (Nanjing Vazyme Biotech Co., Ltd.) together and then transformed into E. coli Trans5a, thereby completing the construction of 28a-eCFP. For 28a-rpoDc-eCFP, the construction was further enhanced by connecting a truncated rpoD fragment to the 28a-eCFP. For the construction of plasmid pK-groES::mCherry, the plasmid pRADK was first linearized using primers pragdk-fp-r and pgroes-fp-r, and the mCherry was amplified using primers fp-f and fp-r. After treatment of the linearized plasmid and mCherry with Exnase II together, the product was transformed into E. coli Trans5a, thereby completing the plasmid construction. For pK-pprI::eGFP, the plasmid pRADK was first linearized using primers pragdk-f and pragdk-fp-r, followed by the amplification of the pprI upstream sequence (the upstream sequence refers to the sequence between this ORF and the previous one) using primers pppri-pr-f and pppri-eg-r, and the amplification of the eGFP gene using primers egfp-f and fp-r. The three linearized sequences were then treated with Exnase MultiS (Nanjing Vazyme Biotech Co., Ltd.) together and then transformed into E. coli Trans5a, thereby completing the plasmid construction. For pK-gC-pI::eG, the obtained plasmid pK-pprI::eGFP was first linearized using primers pragdk-f and pppri-mc-f, followed by the extraction of groes::mCherry sequence from pK-groES::mCherry using primers pgroes-pk-f and fp-r. The above method was then employed to obtain the desired plasmid. For pK-gC-mE::eG and pK-gC-dO::eG, pK-gC-pI::eG was linearized using mCherry-r and egfp-f, with the mqnE upstream sequence and ddrO upstream sequence amplified using the respective primers. Following the same procedure, the desired plasmids were obtained. For other derivative plasmids, the corresponding plasmids were linearized using the corresponding primers, and were then directly transformed it into E. coli Trans5a to complete the construction.
Table S9. Plasmids used in the article.
The preparation and transformation of D. radiodurans–competent cells
The D. radiodurans R1 strain was inoculated into the TGY culture medium and cultured at 30°C with shaking at 200 rpm (Stab S2 Stackable Incubator Shaker, Radobio Scientific Co., Ltd.) until the optical density (OD_600_) reached ∼1.0. 1 ml of the bacterial solution was centrifuged at 6,000 rpm (Centrifuge 5424 R, Rotor FA-45-24-11, Eppendorf) for 3 min to remove the supernatant. The cells were resuspended after adding 250 μl of 4 × TGY and 250 μl of 60 mM CaCl_2_ solution and centrifuged to remove the supernatant. After repeating the resuspension and centrifugation steps once more, 250 μl of 4 × TGY and 250 μl of 60 mM CaCl_2_ solution were added to resuspend the cells, and the cells were incubated at 30°C with shaking for 90 min to complete the preparation of D. radiodurans–competent cells. Subsequently, 500 μl of 50% glycerol was added, and the cells were aliquoted into 1.5-ml EP tubes at 100 μl per tube and stored at −80°C for future use. For transformation, the prepared D. radiodurans–competent cells were thawed on ice, and then, 5 μl of plasmid (∼100 ng/μl) was added. The mixture was incubated on ice for 30 min, followed by the addition of 900 μl of TGY culture medium and further incubation at 30°C with shaking at 200 rpm (Stab S2 Stackable Incubator Shaker, Radobio Scientific Co., Ltd.) for 6 h. After incubation, the cells were centrifuged at 8,000 rpm (Centrifuge 5424 R, Rotor FA-45-24-11, Eppendorf) for 1 min, leaving ∼200 μl of the supernatant. The cell suspension was then spread onto an antibiotic-containing plate, which was then inverted and incubated in a 30°C incubator (LRH-250 Incubator, Shanghai Yiheng Technology Instrument Co., Ltd.). Once colonies reached the appropriate size, they were selected and cultured in TGY liquid medium.
Expression and purification of proteins
The recombinant plasmids 28a-eCFP and 28a-rpoDc-eCFP were transformed into E. coli BL21(DE3). After verification of protein expression, the expressing strain was cultured in 500 ml of LB medium at 37°C with shaking at 200 rpm (Stab S2 Stackable Incubator Shaker, Radobio Scientific Co., Ltd.). This was continued until the optical density at 600 nm (OD_600_) reached ∼0.6. IPTG was then added at a final concentration of 0.2 mM, and the culture was incubated at 30°C with shaking for an additional 6 h to induce protein expression. After the induction, the cells were harvested by centrifugation at 8,000 rpm (Centrifuge 5910 R, Rotor FA-6x250, Eppendorf) for 5 min and washed with PBS. The cell pellet was stored at −80°C for later use. For protein purification, the cells were resuspended in 30 ml of buffer A (1 M NaCl, 5% glycerol, 20 mM Tris–HCl, pH 7.5) and lysed using a cell homogenizer for 40 min, and the lysate was centrifuged to collect the supernatant. The supernatant was then filtered through a 0.22-μm filter. Protein purification was carried out using an AKTA pure 25 system (GE Healthcare) equipped with a nickel column. The system was first equilibrated with buffer A, after which the filtered supernatant was loaded onto the nickel column. The column was washed with 10% buffer B (1 M NaCl, 5% glycerol, 20 mM Tris–HCl, pH 7.5, 500 mM imidazole) to remove nonspecifically bound proteins, followed by a wash with 40% buffer B to elute the target protein. A final wash with 100% buffer B was performed to clean the column. The eluted protein was then subjected to buffer exchange into buffer C (250 mM NaCl, 5% glycerol, 20 mM Tris–HCl, pH 7.5) using a desalting column. The purified protein was stored at 4°C for short-term use or frozen in liquid nitrogen and stored at −80°C.
Fluorescence microscopy observation and flow cytometry counting
The D. radiodurans R1 strain receiving plasmid transformation was inoculated into a 5 ml TGY culture medium and cultured with shaking until the OD_600_ reached ∼5.0. 20 μl of the bacterial culture was stained with DAPI for 5 min and then photographed from the red (filter cube: Texas Red, excitation filter: 560/40, dichroic mirror: 595, barrier filter: 630/60), green (filter cube: FITC, excitation filter: 480/30, dichroic mirror: 505, barrier filter: 535/45), and blue (filter cube: DAPI, excitation filter: 362/396, dichroic mirror: 415, barrier filter: 432/482) channels using 100× oil objective of ECLIPSE Ti2-U (Nikon). In addition, 1 ml bacterial culture was analyzed for eGFP expression using a CytoFLEX flow cytometer (Beckman Coulter, Inc.). Fluorescence intensity below 800 is considered nonfluorescent, and if fewer than 10 out of every 10,000 cells express fluorescence, it is considered that there is no fluorescence expression. E. coli Trans5a harboring the target plasmid was cultured overnight in LB medium, followed by observation and photography under the fluorescence microscope.
Quantitative real-time PCR (qRT–PCR)
The corresponding D. radiodurans strains were inoculated into 50 ml of TGY culture medium and cultured with shaking until the OD_600_ reached ∼1.0, after which the cells were centrifuged and preserved. Total RNA was extracted using TransZol Up Plus RNA Kit (TransGen Biotech), and reverse transcription was carried out on 1 μg of RNA using Reverse Transcriptase M-MLV (Takara). Quantitative real-time PCR was subsequently conducted using TB Green Premix Ex Taq (Takara) and the Mx3005P fluorescence quantitative PCR instrument (Stratagene), following a two-step protocol. The gene of interest was eGFP (primers: egfp-rt-f 5′-CGGCAAGCTGACCCTGAAGT-3′ and egfp-rt-r 5′-TTCATGTGGTCGGGGTAGCGG-3′), whereas the mCherry was used for normalization (primers: mc-rt-f 5′-CAGGACTCCTCCCTGCAGGA-3′ and mc-rt-r 5′-GCGTCGTAGTGGCCGCCGTC-3′).
FRET assays
The 5′-FAM–labeled ssDNA was prepared in a 10 μM solution with double-distilled water (ddH_2_O), whereas the purified eCFP and RpoDc-eCFP were formulated in a 10 μM solution with buffer C. The ssDNA and proteins were added in a 1:1 volume ratio, and the resulting reaction mixture was incubated at 37°C for 30 min. After incubation, the sample was then transferred to a black-bottom, opaque 384-well plate (Corning), and the emission spectra from 460 to 600 nm were recorded using the FlexStation 3 multimode microplate reader (Molecular Devices) with an excitation wavelength of 440 nm. FRET efficiency is obtained by calculating the ratio of emission intensities at 515 and 480 nm. Relative FRET efficiency is obtained by calculating the ratio of FRET efficiency of the same ssDNA incubated with RpoDc-eCFP and eCFP, respectively.
Transcriptome sequencing and transcript quantity statistics
The T. thermophilus HB8 strain was cultured in 50 ml of TGY medium at 70°C in a water bath incubator (THZ-82A Water Bath Incubator Shaker, Changzhou Jintan Liangyou Instrument Co., Ltd.), with shaking at 220 rpm until the OD_600_ reached 1.0. The cells were then pelleted by centrifugation at 8,000 rpm (Centrifuge 5910 R, Rotor FA-6x250, Eppendorf) for 5 min. The harvested bacteria were sent to Novogene Co., Ltd. for transcriptome sequencing. Briefly, sequencing libraries were generated using NEBNext Ultra RNA Library Prep Kit for Illumina (NEB), and library quality was assessed on the Agilent 5400 system (Agilent). Qualified libraries were pooled and sequenced on Illumina platforms with a PE150 strategy. Subsequently, the transcriptome of D. radiodurans (SRX2731648, WT stain without treatment) and T. thermophilus (SRR28238428) was processed for library construction and sequence alignment using the makeblastdb tool. For quantifying transcript levels of target genes, the ORF and 400-bp regions upstream and downstream of the ORF were selected. The target sequence was aligned with the established sequence alignment libraries using the BLAST tool, and results containing gaps or inversions were excluded. Transcript levels for each nucleotide were calculated based on the alignment data, and the statistical results of the ORF and 100-bp regions upstream and downstream of the ORF were documented.
Structure prediction and molecular dynamics simulation
Truncated RpoD proteins from D. radiodurans (WP_227086025.1, Met168-Ile311), T. thermophilus (WP_011228005.1, Tyr84-Ile258), and E. coli (WP_000437376.1, Thr95-Ile450) that interact with the −10 region (−10-motif) were selected for structure prediction. Using the DNA strand 5′-TGTTAAGCTCCC-3′ as a template, the AlphaFold server (Abramson et al, 2024) was employed to predict their structures. After obtaining the predicted structures, the phosphate group at the 5′ end of the DNA strand was removed for molecular dynamics simulations. For the molecular dynamics simulation, the size of the solution system was set to 8 × 10 × 11 nm^3^. Sodium ions and chloride ions were added to reach a salt concentration of 125 mM. The simulations were performed using GROMACS 2024.3 (Abraham et al, 2015) with the AMBER14SB_ParmBSC1 force field (Maier et al, 2015; Ivani et al, 2016) and TIP3P water molecules (Jorgensen et al, 1983). After initial equilibration, all systems were simulated at 310 K and 1 atm for 50 ns with a time step of 2 fs. The molecular dynamics simulations were repeated three times, and the binding free energy (ΔG) between protein and DNA was calculated using the gmx_MMPBSA method (Valdes-Tresanco et al, 2021). The root mean square deviation and energies were recorded every 0.02 ns during the simulation process.
Statistical analysis and reproducibility
The predicted protein structures presented in the article were sourced from the AlphaFold DB, and the protein crystal structures were obtained from the PDB. Because of the absence of a predicted structure for RpoD from D. radiodurans, its structure was predicted using the AlphaFold2 prediction server (Jumper et al, 2021). Structural alignments were conducted using PyMOL v2.6.0a0. DNA sequence diagrams were prepared using SnapGene v6.0.2, and statistical graphs were created with Origin 2019b. All experimental results were successfully replicated at least three times. Unless indicated otherwise, the results represented the means and SD of independent experiments. To compare significant differences, a one-way ANOVA followed by Tukey’s post hoc test was conducted.
Supplementary Material
Reviewer comments
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abraham MJ, Murtola T, Schulz R, Páll S, Smith JC, Hess B, Lindahl E (2015) GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. Software X 1-2: 19–25. 10.1016/j.softx.2015.06.001 · doi ↗
- 2Abramson J, Adler J, Dunger J, Evans R, Green T, Pritzel A, Ronneberger O, Willmore L, Ballard AJ, Bambrick J, (2024) Accurate structure prediction of biomolecular interactions with Alpha Fold 3. Nature 630: 493–500. 10.1038/s 41586-024-07487-w 38718835 PMC 11168924 · doi ↗ · pubmed ↗
- 3Aoyama T, Takanami M, Ohtsuka E, Taniyama Y, Marumoto R, Sato H, Ikehara M (1983) Essential structure of E. Coli promoter: Effect of spacer length between the two consensus sequences on promoter function. Nucleic Acids Res 11: 5855–5864. 10.1093/nar/11.17.58556310517 PMC 326322 · doi ↗ · pubmed ↗
- 4Bae B, Feklistov A, Lass-Napiorkowska A, Landick R, Darst SA (2015) Structure of a bacterial RNA polymerase holoenzyme open promoter complex. Elife 4: e 08504. 10.7554/e Life.0850426349032 PMC 4593229 · doi ↗ · pubmed ↗
- 5Bailey TL, Elkan C (1994) Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc Int Conf Intell Syst Mol Biol 2: 28–36.7584402 · pubmed ↗
- 6Bailey TL, Johnson J, Grant CE, Noble WS (2015) The MEME suite. Nucleic Acids Res 43: W 39–W 49. 10.1093/nar/gkv 41625953851 PMC 4489269 · doi ↗ · pubmed ↗
- 7Beck HJ, Moll I (2018) Leaderless m RN As in the spotlight: Ancient but not outdated! Microbiol Spectr 6: 6.4.02. 10.1128/microbiolspec.RWR-0016-2017 PMC 1163360830006995 · doi ↗ · pubmed ↗
- 8Beck HJ, Fleming IM, Janssen GR (2016) 5′-Terminal AU Gs in Escherichia coli m RN As with shine-dalgarno sequences: Identification and analysis of their roles in non-canonical translation initiation. P Lo S One 11: e 0160144. 10.1371/journal.pone.016014427467758 PMC 4965119 · doi ↗ · pubmed ↗