Ictalurusnazas sp. nov., a new species of North American catfish (Siluriformes, Ictaluridae) from Mexico

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
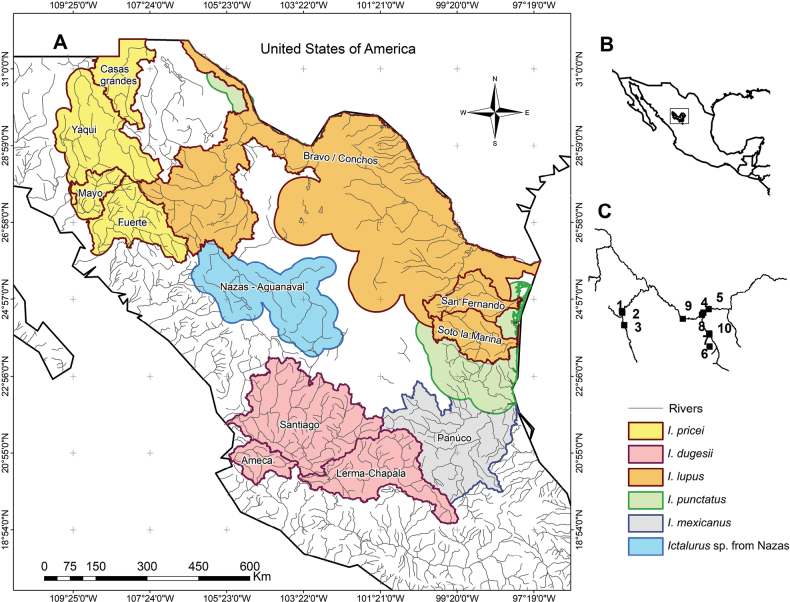 Figure 1
Figure 1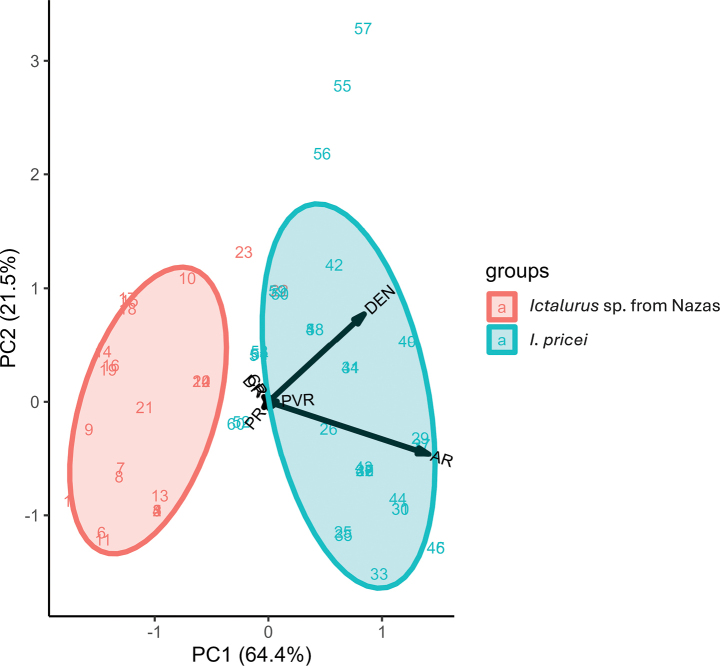 Figure 2
Figure 2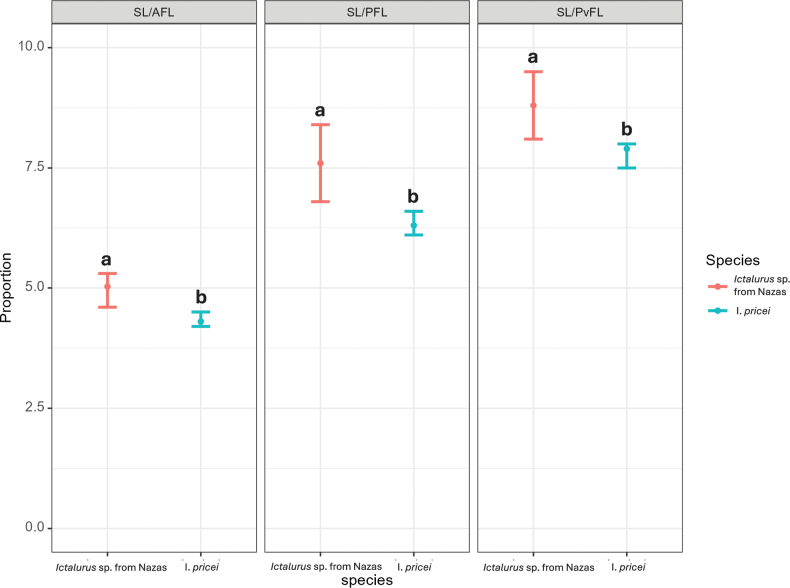 Figure 3
Figure 3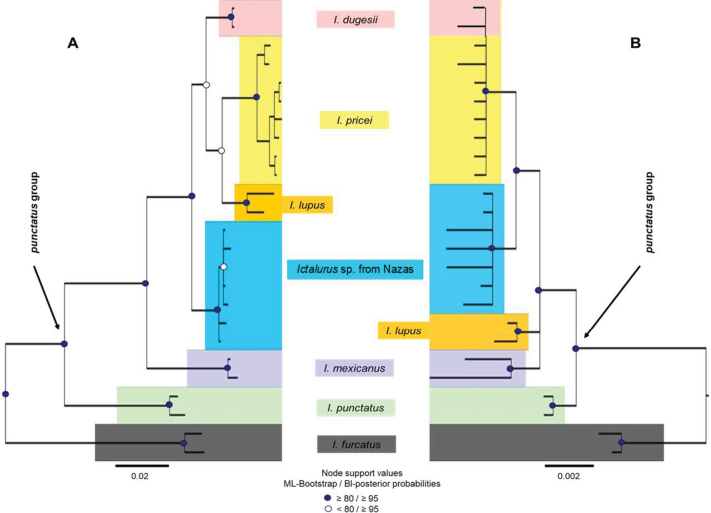 Figure 4
Figure 4 Figure 5
Figure 5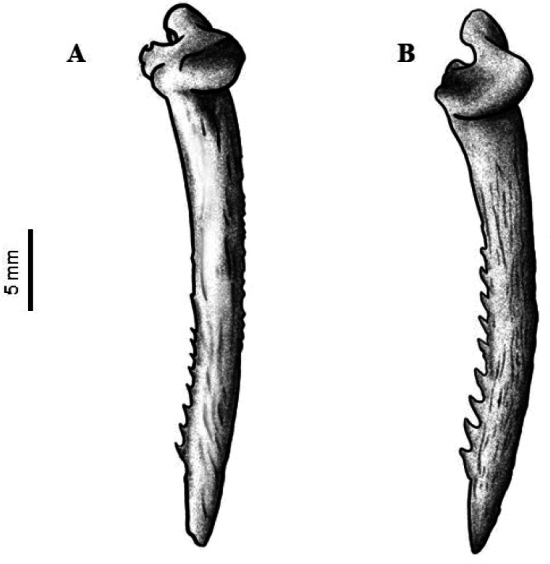 Figure 6
Figure 6 Figure 7
Figure 7| Locality | Drainage | Coordinates | Catalog | |
|---|---|---|---|---|
| 1 | 1.12 km north of El Olote, Santiago Papasquiaro, Durango | Ramos River |
| |
| 2 | El Olote, Santiago Papasquiaro, Durango | Ramos River |
| |
| 3 | River east of Papasquiaro, Durango | Ramos River |
| |
| 4 | Nazas River, main body, Nazas, Durango | Nazas River |
| CNPE-IBUNAM-17795 |
| 5 | Dolores Hidalgo, Nazas, Durango | Nazas River |
| CNPE-IBUNAM-17779 |
| 6 | Balneario “Belem” Peñón Blanco, Durango | Nazas River |
| CPUJED-0018 |
| 7 | El Ranchito, Peñón Blanco, Durango | Nazas River |
| CPUJED-0001 |
| 8 | Arroyo Covadonga, Nazas, Durango | Nazas River |
| CPUJED-0002 |
| 9 | Amoles, Nazas, Durango | Nazas River |
| CPUJED-0003 |
| 10 | Nuevo Covadonga, Peñón Blanco, Durango | Nazas River |
| CPUJED-0004 |
| Aros River, to the south of Natora, Son., Sahuipa, Sonora, Mexico | Yaqui River |
| CNPE-IBUNAM-5712 | |
| Bavispe River to 8.85 km east of Col. Morelos, Bavispe, Sonora, Mexico | Yaqui River |
| ||
| Moctezuma River south of Caserio de Terapa, Moctezuma, Sonora, Mexico | Yaqui River |
| ||
| Bavispe River 7.2 km northeast of the village de Huachinera, Bacerac, Sonora, Mexico | Yaqui River |
| ||
| Guerachi River to 27 km southeast of Guachochi city, Guadalupe y Calvo, Chihuahua | Fuerte River |
| ||
| Guerachi River 27 km southeast of Guachochi city, Guadalupe y Calvo, Chihuahua, Mexico | Fuerte River |
| ||
| Guerachi River 27 km southeast of Guachochi city, Guadalupe y Calvo, Chihuahua, Mexico | Fuerte River |
| ||
| El Sauzal, Urique, Chihuahua, Mexico | Fuerte River |
| CPUJED-0008 | |
| San Isidro Stream, Rosario, Sinaloa, Mexico | Fuerte River | – | CNPE-IBUNAM-20776 |
| Taxa | Locality | Voucher tissue |
|
|
|---|---|---|---|---|
|
| Angulo River at Sabino town; middle Lerma basin, Michoacan, Mexico |
|
| |
|
|
| |||
| Grijalva-Usumacinta River basin, Mexico |
|
| ||
| Apalachicola River, Florida, USA | UF 238203 |
|
| |
|
| San Juan-Bravo, Nuevo Leon, Mexico |
|
| |
| Conchos alto, Durango, Mexico |
|
| ||
| Nazas, Durango, Mexico |
|
| ||
|
|
| |||
| Ramos River, 1.4 km north of Ramos town; Nazas basin Durango, Mexico |
|
| ||
|
|
| |||
|
|
| |||
| El Olote town, Santiago Papasquiaro; Nazas basin, Durango, Mexico |
|
| ||
|
|
| |||
|
| Bavispe River, 7.2 km northeast of Huachinera town; Yaqui basin, Chihuahua, Mexico |
|
| |
| Bavispe River, east of San Miguelito town; Yaqui basin Chihuahua, Mexico |
|
| ||
| Guerachi River, 27 km southwest of Guachochi; Fuerte basin, Chihuahua, Mexico |
|
| ||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
|
|
| |||
| Bolanos-Santiago River basin, Mexico |
|
| ||
| Lake St Clair, Ontario, Canada |
|
| ||
| Gallinas River al Jabali town, San luis Potosi, Mexico | MNCN 2872 |
|
| |
| MNCN 2873 |
|
|
| PC1 | PC2 | |
|---|---|---|
| Eigenvalues | 7.16 | 2.39 |
| Percentage of variance explained | 64.44 | 21.51 |
| Eigenvectors | ||
| Dorsal fin rays | -0.0245 | 0.0468 |
| Caudal fin rays | -0.0204 | 0.0407 |
| Anal fin rays | 0.8716 | -0.4852 |
| Pectoral fin rays | -0.0226 | -0.0481 |
| Pelvic fin rays | 0.0451 | 0.0033 |
| Dentations | 0.4865 | 0.8708 |
| Dorsal rays | Caudal rays | Anal rays | |||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5 | 6 | 7 | 9 | 15 | 16 | 17 | 18 | 19 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | ||||||||||||||||||
| 2 | 16 | 5 | 1 | 2 | 16 | 1 | 5 | 8 | 6 | 9 | 1 | ||||||||||||||||||||||||
|
| 1 | 31 | 4 | 1 | 2 | 25 | 6 | 2 | 10 | 4 | 3 | 10 | 7 | 2 | |||||||||||||||||||||
| Pectoral rays | Pelvic rays | Dentations | |||||||||||||||||||||||||||||||||
| 6 | 7 | 8 | 9 | 10 | 6 | 7 | 8 | 9 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |||||||||||||||||
| 1 | 4 | 11 | 7 | 1 | 1 | 5 | 17 | 1 | 3 | 8 | 4 | 6 | 1 | 2 | |||||||||||||||||||||
|
| 1 | 3 | 30 | 2 | 1 | 34 | 1 | 5 | 15 | 10 | 3 | 1 | 1 | 1 | |||||||||||||||||||||
|
| |||||||
|---|---|---|---|---|---|---|---|
| Holotype | Paratype | ||||||
| Range | Mean | SD | Range | Mean | SD | ||
| Total length | 342 | 106.4-388.6 | 193.3 | 98.7 | 102.9-290 | 179.3 | 46.3 |
| Standard Length | 302 | 87.9-347 | 169.3 | 89.8 | 87.1-208 | 154.2 | 38.9 |
| Head length | 70.2 | 23.6-81.8 | 41.6 | 18.8 | 22.3-53.9 | 39.2 | 9.9 |
| 2.3 | 2.2-3.2 | 2.6 | 0.3 | 2.1-3 | 2.6 | 0.2 | |
| 8.2 | 4.6-9.6 | 6.6 | 1.4 | 4.1-7.2 | 5.9 | 0.7 | |
| 4.3 | 3.4-5.1 | 4.0 | 0.4 | 3.6-4.2 | 3.9 | 0.1 | |
| 5.4 | 4.2-9.4 | 5.6 | 1.4 | 3.8-6.1 | 4.7 | 0.6 | |
| 6.0 | 2.9-6.7 | 5.0 | 0.8 | 3.6-5.1 | 4.4 | 0.4 | |
| 8.7 | 5.8-11.4 | 7.6 | 1.8 | 4.4-7.6 | 6.4 | 0.7 | |
| 10.7 | 6.7-12.5 | 8.8 | 1.6 | 6.8-8.9 | 7.9 | 0.5 | |
|
|
|
|
|
|
| ||
|---|---|---|---|---|---|---|---|
|
| |||||||
|
| 9.4/1.6 | ||||||
|
| 2.5/0.3 | 9.5/1.3 | |||||
|
| 4.7/0.3 | 10.7/1.6 | 5.5/0.4 | ||||
| 2.1/0.6 | 9.6/1.7 | 2.9/0.5 | 5.9/0.7 | ||||
|
| 2.5/0.1 | 9.9/1.6 | 2.6/0.3 | 5.4/0.3 | 3.0/0.6 | ||
|
| 7.3/0.5 | 9.3/1.1 | 8.2/0.3 | 7.6/0.5 | 7.3/0.7 | 7.6/0.5 |
| 1 | Caudal fin deeply forked, with its distal margin in a U-shape and pointed tips; 26 to 32 anal fin rays |
|
| – | Caudal fin moderately forked, with its distal margin in a V-shape and semi-rounded to rounded tips; 27 or fewer anal fin rays |
|
| 2 | Pectoral spine with numerous strong dentations along its posterior margin; head length shorter than anal fin base length |
|
| – | Pectoral spine smooth or corrugated, with few weak posterior dentations; head length equal to or greater than anal fin base length |
|
| 3 | Insertion of the dorsal fin begins at the midpoint of the pectoral fin; 9 to 10 pectoral fin rays; 23 to 25 anal fin rays |
|
| – | Insertion of the dorsal fin begins at the posterior tip of the pectoral fin; 11 pectoral fin rays; 18 to 24 anal fin rays |
|
| 4 | 18 to 24 anal fin rays; maxillary barbels extend to the insertion of the pectoral fin or beyond pectoral spine with few or no weak dentations; the tip of the pectoral spine is sharp; the distal margin of the caudal fin is semi-rounded |
|
| – | 16 to 19 anal fin rays; maxillary barbels do not extend to the insertion of the pectoral fin; pectoral spine with few weak dentations visible from the middle to the tip; tip of the pectoral spine is blunt; distal margin of the caudal fin is completely rounded |
|
| 5 | 20 to 24 anal fin rays; pectoral spine with few weak dentations visible from the middle to the tip |
|
| – | 20 to 21 anal fin rays; posterior face of the pectoral spine lacks dentations |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFish biology, ecology, and behavior · Fish Biology and Ecology Studies · Fish Ecology and Management Studies
Introduction
Ictaluridae currently comprises 50 catfish species distributed across seven genera (Burr et al. 2020) and, together with Leuciscidae (191 species/33 genera; Schönhuth et al. 2018), Percidae (190 species/7 genera; Carlson and Wainwright 2010; Stepien and Haponski 2015), and Catostomidae (75 species/12 genera; Harris et al. 2014), it is considered one of the most species-rich families of freshwater fishes in North America. Since Taylor (1954) proposed the name Ictaluridae for North American catfishes and bullheads, this familial classification has undergone historical changes due to the instability caused by the addition of new information (morphological and molecular data), as well as its lack of robustness (resulting from substantial modifications with the inclusion of new entities). This is evidenced by the incorporation of new characters and entities, populations and/or taxa. Such is the case of the genus Ameiurus Rafinesque, 1820b, which was synonymized with Ictalurus Rafinesque, 1820b by Taylor (1954) and Taylor and Lundberg (1969) and then reinstated to the genus level by Lundberg (1992), who incorporated a phylogenetic approach and additional morphological characters. Subsequent phylogenies based on molecular (Hardman and Page 2003; Hardman and Hardman 2008; Arce-H. et al. 2016; Pérez-Rodríguez et al. 2023; Janzen et al. 2023) and morphological (Arce-H. et al. 2016) data corroborated this classification within Ictaluridae. Recently, the increased volume of molecular data and the application of several new phylogenetic approaches, incorporating mitochondrial and nuclear markers to provide a more detailed evolutionary framework, have revealed limitations of the classification based solely on morphological traits and outdated methodological approaches. For example, ictalurid cave species exhibit morphological similarities due to adaptations to the extreme conditions of their habitats (e.g., absence of light), which have historically led researchers to group them into a single clade. However, the polytypic genus Prietella Carranza, 1954 was later found to be polyphyletic, indicating that Prietellalundbergi Walsh & Gilbert, 1995 should be removed from this genus (Wilcox et al. 2004; Janzen et al. 2023).
Within the genus Ictalurus, questions have arisen from observations of comparative materials for taxonomic identification in ichthyofaunal catalogs, new records, or expert personal assessments regarding the validation of certain species, while also revealing the presence of undescribed species (Lundberg 1992). Several of these findings have been corroborated by recent phylogenetic hypotheses (Janzen et al. 2023; Pérez-Rodríguez et al. 2023). One such undescribed species is Ictalurus sp. from the Nazas River basin, which was originally recognized as a population of Ictaluruspricei (Rutter, 1896) (Meek 1904; Álvarez del Villar 1970), a widely distributed species with several geographically isolated populations that have recently been found to be molecularly well-differentiated (Castañeda-Rivera et al. 2014; Ballesteros-Córdova et al. 2015; Janzen et al. 2023; Pérez-Rodríguez et al. 2023). Among the undescribed populations of I.pricei, those of the Nazas River basin are notable since they are more closely related to Ictalurusdugesii (Bean, 1880) and Ictaluruslupus (Girard, 1858), within the punctatus group (Janzen et al. 2023; Pérez-Rodríguez et al. 2023). Furthermore, in contrast to other I.pricei populations, the origins of which date back to the Pleistocene, the cladogenesis of Ictalurus sp. from the Nazas River basin occurred during the early Pliocene (Janzen et al. 2023; Pérez-Rodríguez et al. 2023). Ictalurus sp. from the Nazas River has even been considered an undescribed species by some authors (Miller 1986; Smith and Miller 1986). However, no taxonomic revision or morphological comparison has been conducted to provide useful characters for the diagnosis of Ictalurus sp. from the Nazas River basin as a distinct taxon.
The aim of this study was therefore to perform a morphological comparison between Ictalurus sp. and I.pricei, the nominal species that has historically included Ictalurus sp. from the Nazas River basin, and to identify useful morphological characters to diagnose this undescribed species from other members of the genus Ictalurus and formally describe it as a new taxon.
Materials and methods
Specimens of Ictalurus sp. were collected throughout the Nazas River basin (Fig. 1, Table 1). All specimens were fixed in 5% formalin, preserved in 70% ethanol, and deposited in the fish collection of the Universidad Juárez del Estado de Durango (FCB-UJED). Specimens deposited in “Colección de Peces de la Universidad Michoacana (CPUM), Facultad de Biología, Universidad Michoacana de San Nicolás de Hidalgo” and “Colección Nacional de Peces (CNP), Instituto de Biología, Universidad Nacional Autónoma de México” were also analyzed.
Distributions of recognized species of the Ictaluruspunctatus group in Mexico. A. River basins in northern and central Mexico inhabited by species of the punctatus group (sensu Pérez-Rodríguez et al. 2023); B. Map of northern and central Mexico showing the distribution of Ictalurusnazas sp. nov.; C. Nazas River basin with sampling sites of the specimens used in this study. The numbers correspond to the locations listed in Table 1.
Meristic counts and linear measurements were taken from the left side of 24 specimens of Ictalurus sp. from the Nazas River basin (hereafter Ictalurus sp. from Nazas) and 36 specimens of I.pricei with a standard length exceeding 100 mm (Table 1), following Hubbs and Lagler (1958). Meristic counts included rays and spines in the dorsal (D), caudal (C), anal (A), pectoral (P), and pelvic (PV) fins. Moreover, three traits were considered derived from the characterization of the pectoral spine, as described by Arce-H. et al. (2016): the number and development of dentations, and their distribution along the posterior face of the pectoral spine. However, only the number of dentations was included in the analysis of meristic counts since it is the only one of these three traits that is discrete.
The linear measurements comprised the standard length (SL), head length (HL), caudal peduncle length (CPL), minimum (minH) and maximum (maxH) height, eye diameter (ED), interorbital distance (IO), anal fin base length (AFL), pectoral fin length (PFL), pelvic fin length (PvFL), body width (BW), and lower jaw length (LJL). All measurements were taken using digital calipers and recorded in mm. The data were then used to calculate the following proportions: head length to standard length, (SL/HL), interorbital distance to head length (HL/ID), eye diameter to head length (HL/ED), lower jaw length to head length (HL/LJL), lower jaw length to standard length (SL/LJL), caudal peduncle length to standard length (SL/CPL), maximum height to standard length (SL/MaxH), minimum height to standard length (SL/MinH), anal fin base length to standard length (SL/AFL), pectoral fin length to standard length (SL/PFL), pelvic fin length to standard length (SL/PvFL), and body width to standard length (SL/BW).
Multivariate and univariate approaches were employed to analyze both meristic and linear trait data to determine whether Ictalurus sp. from Nazas and I.pricei are morphologically disparate. A principal component analysis (PCA) was performed using the prcomp function built into R, based on correlation matrices. To determine significant differences in the proportions of linear measurements between Ictalurus sp. from Nazas and I.pricei, a Student’s t-test was performed using the t-test function built into R. All analyses were conducted in R v. 4.2.1 (R Core Team 2022).
To depict the independent evolutionary history and demonstrate, under a phylogenetic criterion (monophyly), the differentiation level exhibited by Ictalurus sp. from Nazas in relation to the hypothesized species of the genus (sensu Pérez-Rodríguez et al. 2023), particularly its relationship with I.pricei, we conducted a phylogenetic analysis of the genus Ictalurus. Phylogenetic analyses were based on the mitochondrial cytochrome b gene (Cytb) and the nuclear recombination-activation gen 1 (RAG1). All gene sequences were retrieved from GenBank and corresponded to specimens deposited in the Auburn University Museum Fish Collection (AUM), the Academy of Natural Sciences Ichthyology Collection (ANSP), the Ichthyology Collection of the Royal Ontario Museum (ROM), the Mississippi Museum of Natural Science (MMNS), the Florida Museum of Natural History (UF FLMNH), and the fish collection of the Universidad Michoacana de San Nicolás de Hidalgo (UMSNH). This highlights the fact that several sequences for both loci belong to voucher specimens of Ictalurus sp. from Nazas and I.pricei (Table 2).
The obtained sequences were aligned using MEGA X (Kumar et al. 2018) and subsequently checked and manually corrected. The substitution models that best fitted the mitochondrial and nuclear datasets were determined using the program JModelTest 2 (Darriba et al. 2012), based on the Bayesian Information Criterion (BIC). Phylogenetic reconstruction analyses were conducted for the Cytb and RAG1 gene matrices using both the Bayesian inference and maximum likelihood methods. These analyses were performed with MrBayes v. 3.2 (Ronquist et al. 2012) and RAxML v. 8.2.12 (Stamatakis 2006), as implemented in CIPRES (Miller et al. 2010). Moreover, Bagrusubangensis Boulenger, 1902 Cranoglanishenrici (Vaillant, 1893), Noturusgladiator Thomas & Burr, 2004, Pylodictisolivaris (Rafinesque, 1818), and Ameiurusmelas (Rafinesque, 1820a) were used as outgroups (Suppl. material 1: table S1) for both analyses. Finally, uncorrected (p) genetic distances between species of the Ictalurus were calculated for both loci using MEGA X (Kumar et al. 2018).
Results
Principal component analysis using meristic traits revealed two distinct groups corresponding to Ictalurus sp. from Nazas and I.pricei, when plotting the first and second components. The first and second components explained 64.4% and 21.5% of the explained variance, respectively (Fig. 2, Table 3). The characters with the greatest loads, mainly in the first component (C1), were the anal fin rays and the number of posterior dentations on the pectoral spine (Table 3). Ictalurus sp. from Nazas presented fewer anal fin rays (16–19) compared to I.pricei (19–24). Similarly, the number of posterior dentations on the pectoral spine was lower in Ictalurus sp. from Nazas (5–10) than in I.pricei (8–14) (Table 4). The remaining traits (development and distribution of dentations) of the pectoral spine serve to further differentiate the two species. The development of the dentations was weaker in Ictalurus sp. from Nazas compared to that of I.pricei (Fig. 6), and the distribution of the dentations in Ictalurus sp. from Nazas was concentrated towards the apical region of the pectoral spine whereas in I.pricei, these dentations extended from the apical region to beyond the median part of the pectoral spine (Fig. 6).
Principal Components Analysis of the meristic characters used for species discrimination between Ictalurus sp. from Nazas (N = 24) and I.pricei (N = 36).
In contrast to the meristic data, the PCA based on linear measurements did not indicate a clear differentiation pattern (Suppl. materials 1, 2: table S2, fig. S1). However, for the proportions of linear measurements, the Student’s t-test revealed significant differences in three proportions: SL/AFL (t = 3.4, p = 0.0009), SL/PFL (t = 3.8, p = 0.0003) and SL/PvFL (t = 3.1, p = 0.002). In all three cases, Ictalurus sp. from Nazas presented the lowest proportion values (Fig. 3).
Three significant proportional characters distinguishing Ictalurus sp. from Nazas and I.pricei. Values are means with 95% confidence intervals. Different lowercase letters above the error bars indicate significant differences (P < 0.05).
The phylogenetic analyses included sequences from 11 recognized taxa, comprising six recognized species of Ictalurus, including Ictalurus sp. from Nazas, and five outgroups. The final amplicon lengths were 1125 bp for Cytb and 1404 bp for RAG1. The HKY+F+G4 model was selected as the best fit for the Cytb dataset, while the K2P model was chosen for RAG1. Using Bayesian inference and maximum likelihood, the phylogenetic analyses revealed seven groups in the topology based on Cytb (Fig. 4A), recovering the six recognized species as monophyletic groups, plus Ictalurus sp. from Nazas. In contrast, the topology based on the nuclear locus revealed six groups, recovering only four recognized species as monophyletic groups, a further group formed by I.pricei and I.dugesii, and another comprising Ictalurus sp. from Nazas (Fig. 4B).
Bayesian inference and maximum likelihood hypothesis of the genus Ictalurus. Hypothesis based on A. Mitochondrial gene Cytb, and B. Nuclear gene RAG1
P-distances between the members of Ictalurus ranged from 2.1 to 9.9% for Cytb (Table 6) and from 0.1 to 1.6% for RAG1 (Table 6). Based on the meristic, morphometric, and molecular evidence, a new species of the genus Ictalurus was described. Descriptive statistics of the meristic data, linear measurements, and proportions for the holotype and paratypes are presented in Tables 4, 5.
Ictalurus
nazas
sp. nov.
Taxon classificationAnimaliaSiluriformesIctaluridae
2512F819-1611-54AB-ACC1-953C9DE8D558
https://zoobank.org/A26FD24E-C28C-4C72-8D82-BD3995E470D2
Type locality.
Ramos River, 1.12 km north of El Olote, Santiago Papasquiaro, Durango, Mexico.
Type materials.
Holotype. • UMSNH-2654, adult, 302 mm SL, Ramos River 1.12 km north of El Olote, Santiago Papasquiaro, Durango, Mexico, endorheic drainage Ramos River; 25°14'13.2"N, -105°27'0.4"W, collected on June 2, 2008. Paratypes. • UMSNH-7266 (N = 1), El Olote, Santiago Papasquiaro, Durango, Mexico, endorheic drainage Ramos River; 25°13'17.04"N, -105°26'37.79"W, collected on May 10, 2004.; • UMSNH-1642 (N = 3) river to the east of Santiago Papasquiaro, Durango, Mexico, endorheic drainage Ramos River; 25°2'26.12"N, -105°24'54.67"W, collected on September 19, 2005; • CNPE-IBUNAM-17795 (N = 5) Nazas River, main body, Nazas, Durango, Mexico, endorheic drainage Nazas River; 25°12'57"N, -104°10'25"W, collected on April 4, 2004; • CNPE-IBUNAM-17779 (N = 1) Dolores Hidalgo, Nazas, Durango, Mexico, endorheic drainage Nazas River; 25°15'59"N, -104°05'24"W, collected on April 4, 2004; • CP-UJED-0018 (N = 1) Balneario “Belem” Peñón Blanco, Durango, Mexico, endorheic drainage Nazas River; 24°44'36"N, -104°04'36"W, collected on September 18, 1989; • CPUJED-0001 (N = 4 El Ranchito, Peñón Blanco, Durango, Mexico, endorheic drainage Nazas River; 24°59'19"N, -104°19'39"W, collected on August 5, 2015; • CPUJED-0002 (N = 2) Arroyo Covadonga, Nazas, Durango, Mexico, endorheic drainage Nazas River, 25°11'31"N, -104°11'41"W, collected on December 13, 2022; • CPUJED-0003 (N = 1) Amoles, Nazas, Durango, Mexico, endorheic drainage Nazas River, 25°07'47"N, -104°29'47"W, collected on December 1, 2022; • CPUJED-0004 (N = 4) Nuevo Covadonga, Peñón Blanco, Durango, Mexico, endorheic drainage Nazas River, 24°54'35"N, -104°04'47"W; collected on May 22, 2023; • CPUJED-0005 (N = 1) El Ranchito, Peñón Blanco, Durango, Mexico, endorheic drainage Nazas River; 24°59'19"N, -104°19'39"W, collected on May 21, 2023.
Diagnosis.
Ictalurusnazas sp. nov. differs from I.pricei by the following combination of characters: 16–18, rarely 19, anal fin rays (vs. 19–24); shorter premaxillary barbel, reaching only to the opercle margin (vs. extending beyond the margin of the opercle); shorter base of anal fin, 5.0 times into the SL (vs. larger base, 4.4 times into the SL); shorter length of pectoral fin, 7.6 times into the SL (vs. larger pectoral fin, 6.4 times into the SL); shorter length of pelvic fin, 8.8 times into the SL (vs. larger pelvic fin, 7.9 times into the SL); lower number of posterior dentations, 5–8, rarely 9 and 10, (vs. higher number of dentations, 8–11, rarely 12–14 dentations) (Fig. 6); weaker development of dentations (vs. stronger development) (Fig. 6), and dentations concentrated in the apical part of the pectoral spine (vs. dentations extended from apical to median part of the pectoral spine) (Fig. 6).
Ictalurusnazas sp. nov. holotype, UMSNH-2654, adult, SL 302 mm, Ramos River, 1.12 km north of El Olote, Santiago Papasquiaro, Durango, Mexico, endorheic drainage Ramos River.
View of the pectoral spine of Ictalurus species: A. I.nazas sp. nov., 33 mm. and B. I.pricei, 28 mm. Ictalurusnazas sp. nov. exhibits fewer posterior dentations, which are underdeveloped and can only be distinguished from the middle of the spine towards the apex.
In relation to other recognized species of Ictalurus (sensu Pérez-Rodríguez et al. 2023), Ictalurusnazas sp. nov. can be distinguished from its congeners, I.lupus (Miller et al. 2005), I.dugesii (Arce-H. et al. 2016), I.mexicanus (Arce-H. et al. 2016), I.punctatus (Miller et al. 2005), I.furcatus (Miller et al. 2005), and I.balsanus (Mejía et al. 2013), by presenting an anal fin base shorter than the head length, with 16 to 19 anal fin rays. Additionally, the premaxillary barbels do not extend beyond the posterior margin of the opercle (see Description for further details). Adult specimens of Ictalurusnazas sp. nov. also differ from most congeners, except I.dugesii, by exhibiting fewer and less developed posterior dentations on the pectoral spine, which are restricted to its apical portion.
Moreover, Ictalurusnazas sp. nov. can be diagnosed by the following characters: from I.punctatus by having a moderately forked caudal fin with a distal margin forming an “U” shape and fully rounded (vs. deeply forked caudal fin with a “V” shaped margin and pointed tips); from I.lupus, by having 7 to 9 (rarely 6 and 10) pectoral fin rays (vs. 9 to 10 rays); from I.mexicanus, by the presence of posterior dentations in the pectoral spine (vs. absence); from I.dugesii by presenting a blunt tip on the pectoral spine (vs. sharp tip); from I.furcatus by presenting fewer anal fin rays (16 to 19 vs. 28–38 rays), and from I.balsanus by presenting 17 to 19 caudal fin rays (vs. 22 rays).
Description.
Elongated body; large head, ventrally flattened, tapering anteriorly. Eyes small, diameter, 9.6 times into head the length, interorbital distance, 3.2. times into the head length. Terminal mouth position with eight barbels: four maxillary, two premaxillary, and two nasals. The premaxillary barbels do not reach the edge of the operculum.
Dorsal fin with 5 to 7, rarely 9 rays, its origin positioned at the midpoint of the pectoral fins. Pectoral fin length 8.4 times into the SL and contains 7 to 9 rays, though, on occasions, specimens may exhibit 6 or 10 rays. The pectoral spine features a series of 5 to 8 posterior dentations with weaker development; in rare cases, 9 or 10 dentations may be present. Pelvic fin is short, 9.5 times into the SL, with 7 to 8 rays, rarely 6 and 9; origin posterior to the base of the dorsal fin. Anal fin base shorter than head length, its length fitting 5.0 times into the SL; fin composed of 16 to 19 rays. Caudal peduncle short and moderately forked, with a rounded distal margin forming a “U” shape, with 17 to 19 rays.
Color. In life, the body is olive green, with a slightly darker pigmentation on the dorsal part of the head, fins, and premaxillary barbels. Ventral region, along with the maxillary and nasal barbels, appears noticeably lighter. The body features a few dark spots, while the caudal fin margin is distinctly black. In alcohol, the body takes on a gray hue, with the fins and ventral region significantly lighter. The fin margins remain black, and the premaxillary barbels appear darker than the maxillary and nasal barbels.
Habitat and ecology.
The species inhabits slow-moving waters with temperatures ranging from 17 to 22 °C. It is generally found in areas with exposed bedrock slabs and crevices that provide refuge.
Distribution.
Ictalurusnazas sp. nov. is only known from the endorheic basin of the Nazas-Aguanaval System, including its main tributaries: the Rivers Peñón Blanco and Ramos (Fig. 7).
Type locality of I.nazas sp. nov. Ramos River, 1.12 km north of El Olote, Santiago Papasquiaro, Durango, Mexico, endorheic drainage, Ramos River; 25°14'13.2"N, -105°27'0.4"W.
Etymology.
The specific epithet nazas is derived from the name of the basin in which the species is distributed and to which it is endemic.
Discussion
Although Ictalurusnazas sp. nov. was considered an undescribed species during the last century (sensu Miller 1986; Smith and Miller 1986), the studies that were conducted were limited to lists and regionalization of the ichthyofauna from biogeographical provinces of North America, without performing a detailed morphological comparison or description. The strength of the molecular evidence, combined with increased taxonomic sampling within Ictalurus, has provided more evidence for the recognition of this new taxon and further contributed to resolving the systematics of this genus. The diagnostic morphological characters of I.nazas sp. nov. are distinct and differentiate this new taxon, particularly from I.pricei, the nominal species to which it was originally assigned. These same characteristics, in combination with others, have also proved useful for clearly distinguishing this new taxon from the remaining species of Ictalurus (see Dichotomous key). Although the two phylogenetic trees showed minor inconsistencies with each other, both loci consistently recovered I.nazas sp. nov. within the punctatus group as a well-supported, reciprocally monophyletic lineage (Fig. 4). Uncorrected p-distances based on Cytb between I.nazas sp. nov. and its closest congeners ranged from 2.1% to 2.9% (Table 6), which falls within the minimum interspecies divergence limits for Cytb within Ictaluridae (1.8–3.6%) (Hardman and Page 2003; Hardman 2004; Egge and Simons 2006; Egge et al. 2015; Pérez-Rodríguez et al. 2023). Both the phylogenetic relationships and molecular divergence estimates indicate an independent evolutionary history that began approximately five million years ago (Janzen et al. 2023; Pérez-Rodríguez et al. 2023). This history is consistent with the isolation of I.nazas sp. nov. in the ancient Nazas River paleosystem during the Pliocene (Domínguez-Domínguez et al. 2011) and constitutes strong evidence to support its recognition as a new and independent taxonomic entity.
This finding is consistent with other endemic fish species from the Nazas River, such as the leuciscids Notropisnazas Meek, 1904 (Contreras-Balderas et al. 2005), Gilaconspersa Garman, 1881 (Contreras-Balderas et al. 2005; Schönhuth et al. 2014), Cyprinellagarmani Jordan, 1885 (Contreras-Balderas et al. 2005), and Cyprinellaalvarezdelvillari Contreras-Balderas & Lozano-Vilano, 1994 (Contreras-Balderas et al. 2005), the catostomid Pantosteusnebuliferus (Garman, 1881) (Contreras-Balderas et al. 2005; Corona-Santiago et al. 2018), and the cyprinodontid Cyrpinodonnazas Miller, 1976 (Echelle et al. 2005). These findings have important biogeographic and conservation implications. Although the Nazas River basin is currently an endorheic system, it has been considered a historical transitional zone between freshwater fish lineages originating from the Gulf of Mexico and Pacific slope drainages in North America. This is supported by evidence of the colonization of the genus Moxostoma Rafinesque, 1820b from the Gulf slope into the Pacific slope drainages during the early Pliocene (Pérez-Rodríguez et al. 2016). The two phylogenetic hypotheses, based on Cytb and RAG1 presented herein, suggest that the ancestor of Ictalurusnazas sp. nov. and the species currently distributed in the Pacific slope drainages (I.pricei and I.dugesii) likely inhabited the region corresponding to the paleo-Nazas system. Thus, this area also appears to have served as a transitional zone between Ictalurus species from the Gulf of Mexico slope and those from the Pacific slope drainages. Indeed, previous phylogenetic hypotheses and divergence time estimates (Janzen et al. 2023; Pérez-Rodríguez et al. 2023) indicate that the ancestor of Ictalurusnazas sp. nov. originated in the early Pliocene, supporting a biogeographic transition similar to that hypothesized for Moxostoma species. The addition of this new taxon increases both the richness of the genus Ictalurus in Mexico and the level of endemism for the Nazas River, suggesting that the Nazas River basin should be considered an important biogeographic province in North America that merits additional conservation protections.
We further highlight the fact that this study is part of an extensive investigation throughout the Nazas basin, with a high completeness of sampling within the basin and its tributaries. Despite this, only specific populations of I.nazas sp. nov. were found at the sites mentioned above. Immediate management and conservation measures are therefore recommended, together with further research on the biology and ecology of this species. These studies could ultimately support the inclusion of I.nazas sp. nov. in vulnerability categories at both national and international levels, ensuring that conservation efforts are appropriately directed, given its status as a microendemic species.
Dichotomous key to the Ictalurus species of the punctatus group
**: **
Comparative material
Ictaluruspricei: CNPE-IBUNAM-5712 (N = 2) Aros River, to the south of Natora, Son., Sahuipa, Sonora, Mexico; UMSNH-2931, (N = 1), Bavispe River, at 8.85 km east of Col. Morelos, Bavispe, Sonora, Mexico; UMSNH-7763, (N = 3), Moctezuma River, to the south of Caserio de Terapa, Moctezuma, Sonora, Mexico; UMSNH-2221, (N = 5), Bavispe River, at 7.2 km northeast of the village de Huachinera, Bacerac, Sonora, Mexico; UMSNH-2642, (N = 5), Guerachi River, at 27 km southeast of Guachochi city, Guadalupe y Calvo, Chihuahua, Mexico; UMSNH-2534 (N = 5), Guerachi River, to southeast of Guachochi city, Guadalupe y Calvo, Chihuahua, Mexico; UMSNH-7764 (N = 6), Guerachi River, at 27 km southeast of Guachochi city, Guadalupe y Calvo, Chihuahua, Mexico; CPUJED-0008 (N = 10), El Sauzal, Urique, Chihuahua, Mexico; CNPE-IBUNAM-20776 (N = 2) San Isidro Stream, Rosario, Sinaloa, Mexico.
Supplementary Material
XML Treatment for Ictalurus nazas
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Álvarez del Villar J (1970) Peces Mexicanos (claves). Servicio de investigación pesquera. Instituto nacional de Biología Pesquera. Mexico. 166 pp.
- 2Arce-HM Lundberg JGO’Leary MA (2016) Phylogeny of the North American catfish family Ictaluridae (Teleostei: Siluriformes) combining morphology, genes and fossils.Cladistics 33(4): 406–428. 10.1111/cla.1217534715724 · doi ↗ · pubmed ↗
- 3Ballesteros-Córdova A Castañeda-Rivera M Grijalva-Chon JM Castillo-Gámez RA Gutiérrez-Millán LE Camarena-Rosales F Ruíz-Campos G Varela-Romero A (2015) Complete mitochondrial genome of Ictaluruspricei (Teleostei: Ictaluridae) and evidence of a cryptic Ictalurus species in Northwest Mexico. Mitochondrial DNA.Part A, DNA Mapping, Sequencing, and Analysis 27(6): 4439–4441. 10.3109/19401736.2015.108956126405008 · doi ↗ · pubmed ↗
- 4Bean TH (1880) Descriptions of two species of fishes collected by Prof. A. Dugès in central Mexico.Proceedings of the United States National Museum 2(95): 302–305. 10.5479/si.00963801.2-95.302 · doi ↗
- 5Boulenger GA (1902) Additions à la faune ichthyologique du bassin du Congo. Annales du Musée du Congo (Série Zoologique) 2(2): 19–57 [pls 7–16].
- 6Burr BM Warren Jr ML Bennett MG (2020) Ictaluridae: North American Catfishes. In: Warren ML Burr BM Echelle AA Kuhajda BR Ross ST (Eds) Freshwater fishes of North America.Johns Hopkins University Press, Maryland, 23–100.
- 7Carlson RL Wainwright PC (2010) The ecological morphology of darter fishes (Percidae: Etheostomatinae). Biological Journal of the Linnean Society.Linnean Society of London 100(1): 30–45. 10.1111/j.1095-8312.2010.01417.x · doi ↗
- 8Carranza J (1954) Descripción del primer bagre anoftalmo y depigmentado encontrado en aguas mexicanas. Ciencia (México, D.F.)14(7–8): 129–136.