A new species of Galathea (Decapoda, Galatheidae) from the seamounts of the Easter Island area (Southeast Pacific Ocean Ridge) associated with a sea urchin

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1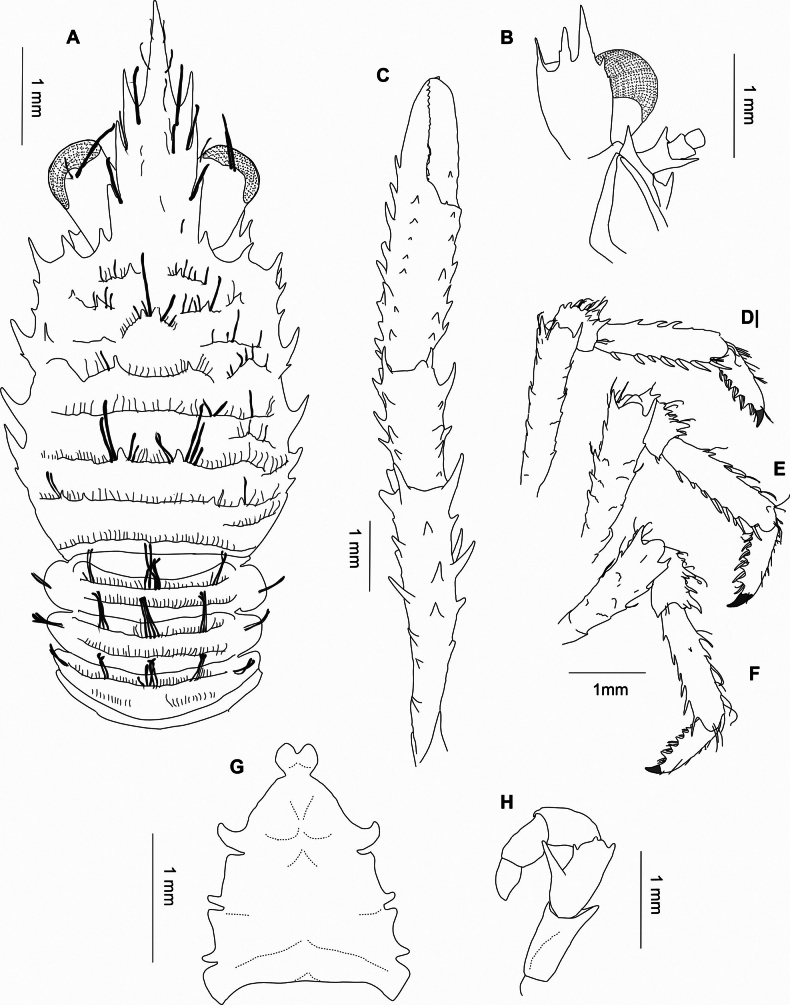 Figure 2
Figure 2 Figure 3
Figure 3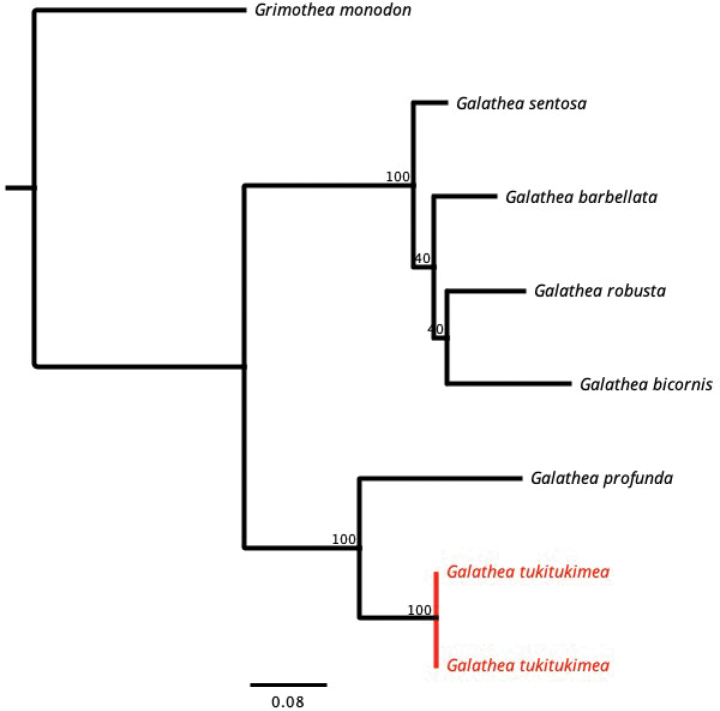 Figure 4
Figure 4| Species | GenBank accession no. ( | Locality | Biogeographic region | Depth (m) | Reference |
|---|---|---|---|---|---|
|
|
| Vanuatu and New Caledonia | Southwest Pacific Ocean | 182–270 |
|
|
|
| New Caledonia | Southwest Pacific Ocean | 81 |
|
|
|
| Vanuatu and New Caledonia | Southwest Pacific Ocean | 270–702 |
|
|
|
| Madagascar, La Réunion, Mauritius | Western Indian Ocean | 105–238 |
|
|
|
| Wallis and Futuna Islands | Southwest Pacific Ocean | 245–440 |
|
|
| Motu Motiro Hiva | Southeastern Pacific Ocean | 407 | This study | |
|
| Pukao seamounts | Southeastern Pacific Ocean | 348 | This study | |
|
| Chile Continental | Southeastern Pacific Ocean | 94–523 |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCrustacean biology and ecology · Marine Biology and Ecology Research · Coral and Marine Ecosystems Studies
Introduction
The family Galatheidae contains many species, primarily included in the genus Galathea Fabricius, 1793 (e.g. Baba et al. 2008; Macpherson and Baba 2011). The genus is one of the most species-rich of anomuran decapods, containing more than 170 species, most of which are distributed in the Indo-West Pacific Ocean (e.g. Baba et al. 2008; Schnabel et al. 2011; Macpherson and Robainas-Barcia 2015). A significant number of these species have been described in recent years (e.g. Macpherson and Robainas-Barcia 2015; Tiwari et al. 2022, 2024; Rodríguez-Flores et al. 2024; Macpherson et al. 2025) thanks to the increase in sampling efforts (e.g. Richer de Forges et al. 2013; Rodríguez-Flores et al. 2024; Gaymer et al. 2025). These results emphasize that despite the intensive taxonomic efforts, our knowledge gap is still evident in squat lobsters (De Grave et al. 2023).
These species of squat lobsters are found mainly in the Western and Central Pacific but are absent in the Eastern Pacific. The citation of Galatheapaucilineata Benedict, 1902, from the Galapagos Islands probably is a species of Leiogalathea (Rodríguez-Flores pers. comm.). The easternmost occurrences of Galathea in the Pacific are G.senta Macpherson & Robainas-Barcia, 2015 from the Pitcairn Islands (Oeno Island, 23°55'S, 130°44'W) and another approximately 15 species from Gambier or the Austral Islands in French Polynesia (Macpherson and Robainas-Barcia 2015).
The seamounts of the Southeastern Pacific have been the focus of increasing sampling effort over the past decades (Fernández et al. 2014; Gaymer et al. 2025), which has facilitated the discovery of numerous new species, including squat lobsters and other groups of benthic invertebrates (Sellanes et al. 2019; Díaz-Díaz et al. 2020; Gallardo et al. 2021; Cañete et al. 2023; Tenorio et al. 2023; de Souza et al. 2024).
During 2024, aboard the RV Falkor (Too), a more thorough sampling of the Salas y Gómez Ridge was carried out in both international waters and within the Chilean Exclusive Economic Zone (EEZ) (FKt240224 Expedition). The technological capabilities of the ROV SuBastian, from the Schmidt Ocean Institute, allowed the discovery of new records and species, as well as the acquisition of HD images that will shed light on ecological associations and habitat use of benthic species. During the expedition, 24 dives were conducted between 200 and 1200 m depth, 11 of them within the Chilean EEZ, including dives around the islands of Rapa Nui and Motu Motiro Hiva, and six seamounts within the Rapa Nui Multiple Use Marine Coastal Protected Area (AMCP-MU Rapa Nui) and the Motu Motiro Hiva Marine Park (MMH Park).
In the present paper, we describe and illustrate, using morphological and molecular data (COI), a new species of Galathea collected around AMCP-MU Rapa Nui and in MMH Park and associated with the sea urchin StereocidarisnascaensisAllison et al., 1967. This finding represents the first record of Galathea in the Southeastern Pacific and the first documented association between a sea urchin and a species of this genus. To facilitate taxonomic comparisons among morphologically closely related species, we also provide the phylogenetic relationships of some species within the genus Galathea.
Materials and methods
Sampling and measurements
The material examined is in the Museo Nacional de Historia Natural (MNHNC, Santiago, Chile) and Sala de Colecciones Biologicas UCN (SCBUCN, Coquimbo, Chile). The terminology and measurements follow Baba et al. (2011) and Macpherson and Robainas-Barcia (2015). The size of the specimens is indicated by the postorbital carapace length (CL), measured along the midline from the base of the rostrum to the posterior margin of the carapace. The length of the rostrum is measured from the tip to between the lateral basal incisions, the breadth is between left and right lateral basal incisions. Measurements of appendages were taken in dorsal (pereopod 1), lateral (antennule, pereopods 2–4) and ventral (antenna) midlines. Abbreviations used are: Mxp3, maxilliped 3; P1–P4, pereopods 1–4; M = male; F = female; ovig. = ovigerous.
Molecular analysis
The DNA extraction, amplification of the cytochrome c oxidase subunit (COI) of the two specimens, and subsequent sequencing were done following the workflow optimized in previous studies on squat lobster’s systematics (e.g. Rodríguez-Flores et al. 2021). DNA was extracted with the DNeasy Blood and Tissue kit (Qiagen), following the manufacturer’s protocol after an overnight digestion. The DNA integrity was assessed on a 2% agarose gel, and its purity was verified by the A260/280 ratio. We used the primers LCO1490 5′ GGTCAACAAATCATAAAGATATTGG-3′ HCO2198 5′-TAAACTTCAGGGTGACCAAAAAATCA-3′ (Folmer et al. 1994; Zuccon et al. 2012), for the partial amplification of the mitochondrial COI. PCRs (25 µL total volume) were performed using 2.5 µL of 10 × standard Taq reaction buffer (New England Biolabs), 2 µL of dNTP Mix (2.5 mM), 0.5 µL of each primer (10 mM), 0.125 µL of Taq DNA polymerase (New England Biolabs), 1 µL of BSA (0.06 mg/mL), 1 µL of MgCl_2_ (25 mM), 1 µL of template DNA, and nuclease-free water to complete the final volume. The PCR program consisted of an initial denaturation at 94 °C for 1 min, followed by 5 cycles of 94 °C for 40 s, annealing at 45 °C for 40 s, and extension at 68 °C for 1 min. This was followed by 35 cycles of 94 °C for 40 s, annealing at 57 °C for 40 s, and extension at 68 °C for 1 min, with a final extension step at 68 °C for 5 min. The resulting amplicons were visualized in agarose 1% gels, and the PCR products were sent to Macrogen, Inc. (Seoul, South Korea) for DNA Sanger sequencing.
Forward and reverse raw sequences were aligned, manually edited, and trimmed using Geneious Prime v. 2025.1.2 (https://www.geneious.com). Galatheatukitukimea sequences were aligned with other Galathea sequences available in GenBank from morphologically closely related species (G.barbellata, G.bicornis, G.profunda, G.robusta, and G.sentosa). Galatheatukitukimea consensus sequences were uploaded to GenBank with accession numbers PV448654–PV448655 (Table 1). Genetic distances for comparisons were estimated using uncorrected ‘p’ divergences in PAUP* v. 4.0 (build 169) (Swofford 2002). COI alignment was performed using MAFFT v. 7.490 (Katoh and Standley 2013) in Geneious Prime with default settings. The COI alignment was used to obtain a phylogenetic reconstruction with the PhyML v. 3.3.2 plugin with the following settings: substitution model = GTR, bootstrap = 1000, the proportion of invariable sites = estimated, gamma distribution parameter = estimate, optimize = Topology/length/rate (Guindon et al. 2010).
Systematic account
Superfamily Galatheoidea Samouelle, 1819
Family Galatheidae Samouelle, 1819
Genus Galathea Fabricius, 1793
Galathea
tukitukimea
sp. nov.
Taxon classificationAnimaliaDecapodaGalatheidae
F2ED9100-5771-5124-B666-B92C1A642810
https://zoobank.org/906F5BDA-B2FD-43F4-94C3-2E31E7219838
Material examined.
Salas and Gómez Ridge. Holotype: • Motu Motiro Hiva (Salas and Gómez Island). Cruise FKt240224, Stn 671-BB1A-005, 26°28'35"S, 105°13'58"W, 407 m, 20 March 2024: M 3.3 mm (MNHNC DEC-15582). Paratype: • Pukao seamount. Cruise FKt240224, Stn 667-BB1D-002, 26°54'52"S, 110°14'26"W, 348 m, 16 March 2024: ovig. F 4.4 mm (SCBUCN6722).
Etymology.
The specific epithet tukitukimea derives from the Rapa Nui words tuki tuki mea, meaning “red dots”, in reference to the vivid reddish spots on the carapace and pereiopods. The name was proposed by Serafina Moulton Tepano, a Rapa Nui artist who accompanied the FKt240224 expedition. It is treated as a noun in apposition.
Description.
Carapace: longer than broad; anterior and posterior cervical grooves distinct; dorsal surface with scale-like and interrupted ridges in all regions; mid-transverse ridge laterally interrupted and not scale-like, preceded by distinct cervical groove; transverse groove before cardiac spines; ridges not densely setose, with short simple setae (sometimes with a few short setules) and with some long and median thick, plumose setae on protogastric and cardiac ridges. Epigastric region with 2 spines and 1 or 2 minute acute granules; 2 median protogastric spines; 1 lateral protogastric and 1 parahepatic spine on each side; 2 median metagastric spines. Cardiac region with 2 median spines. Branchial regions each with 1 anterior branchial 2 posterior spines. Lateral margins slightly convex, with 8 spines: 3 spines in front of and 5 spines behind anterior cervical groove; first anterolateral, well developed, posterior to level of lateral limit of orbit; second and third spines small, situated at midlength between anterolateral spine and anterior cervical groove; 2 spines on anterior branchial region, second very small, and 3 spines on posterior branchial margin, third very small; 1 additional spine below lateral margin, between first and second anterolateral spines. Outer orbital angle acute; infra-orbital margin with 1 or 2 spines. Rostrum 2.7 × as long as broad, length 0.8 that of, breadth 0.3 that of carapace; distance between distalmost lateral incisions 0.4 distance between proximalmost lateral incisions; dorsal surface nearly horizontal in lateral view, with minute setiferous ridges; lateral margin with 4 sharp spines.
Pterygostomian flap rugose with sparse setae, anteriorly rounded; some granules on upper margin near linea anomurica.
Pleon: somites 2 and 3 each with 2 uninterrupted transverse ridges on tergite; somite 4 with 1 uninterrupted and 1 medially interrupted transverse ridge; somites 5 and 6 each with scale-like ridges, each tergite with some long and thick plumose setae; posteriormedian lobe of somite 6 indistinct. Telson incompletely subdivided. Two pairs of male gonopods.
Thoracic sternum: 1.2 × as long as wide. Sternite III with median shallow notch. Sternite IV with anterior part narrower than sternite III, with some short striae. Sternites IV–VI with a few striae. Sternite III ~1.7 × as wide as long; sternite IV nearly 2.1 × as wide as long, and 3.5 × as wide as sternite III.
Eye: ocular peduncles 1.8 × longer than broad, maximum corneal diameter 0.7 of rostrum width.
Antennule: article 1 with 3 distal spines, distomesial smaller than distolateral, distodorsal spine larger than others; lateral margin with 2 small spines.
Antenna: article 1 with distomesial spine not reaching distal margin of article 2. Article 2 with distomesial shorter than distolateral spine, not reaching end of article 3. Articles 3 and 4 unarmed.
Mxp3: ischium with spine on extensor and flexor distal margins. Merus subequal in length to ischium, with strong median and small distal spine on flexor margin; extensor margin with small distal spine. Carpus smooth along extensor margin.
P1: 3.3 × carapace length, relatively slender, somewhat depressed on palm, more so on fingers. Merus 1.4 × as long as carapace, 3 × as long as carpus, with spines arranged roughly in rows, distal spines prominent. Carpus 3.4 × as long as palm, 1.6 × longer than broad; dorsal surface with small spines arranged roughly in longitudinal rows; mesial margin with 2 or 3 strong spines. Palm 4.3 × longer than broad, lateral and mesial margins subparallel; spines arranged roughly in rows; dorsolateral row continuing onto lateral margin of fixed finger. Fingers 0.4 × as long as palm; movable finger ending in a slightly curved spine; flexor margin with 6 or 7 serrated distal teeth, and a strong subtriangular tooth proximally, finely setose.
P2–4: moderately slender. P2 1.8 × carapace length. Meri successively shorter posteriorly (P3 merus 0.8 length of P3 merus, P4 merus 0.7 length of P3 merus); P2 merus 0.7 of carapace length, 4.5 × as long as broad, 1.5 × longer than P2 propodus. Extensor margin with row of 5 or 6 proximally diminishing spines on P2 and P3, 2 or 3 distal spines on P4; ventral margins distally ending in strong spine followed proximally by 2 or 3 spines and several eminences. Carpi with 5 or 6 spines on extensor margin on P2–4; lateral surface with 3 or 4 spines or acute granules subparallel to extensor margin; flexor distal margin acute. Propodi 3.5–4.0 × as long as broad; extensor margins with 3 or 4 proximal spines; flexor margin with 6–8 slender movable spines, terminal spines paired. Dactyli distally ending in well-curved, strong spine, length 0.5–0.6 that of propodi; flexor margin with 5 proximally diminishing teeth, terminal tooth prominent.
Epipods present on P1, absent on P2 and P3.
Genetic data.
COI. 671-BBIA-005, GenBank code: PV448654. 667-BBID-002, GenBank code: PV448655.
Remarks.
The new species belongs to the species group having the carapace with paired median protogastric and cardiac spines. This group now contains eight species: G.barbellata Macpherson, 2012 from New Caledonia, G.bicornisMacpherson et al., 2025 from New Caledonia, G.echinata Macpherson, 2012 from New Caledonia, G.profunda Macpherson, 2012 from New Caledonia and Vanuatu, G.robusta Baba, 1990 from Madagascar, G.sentosa Macpherson & Robainas-Barcia, 2015 from Wallis and Futuna, and the new species from Salas and Gómez Ridge. The new species can be distinguished from these species by the following characters:
Galatheabicornis, G.echinata, G.robusta, and G.sentosa have the antennular article 1 with 2 well-developed distal spines, with the distomesial spine minute or obsolescent, whereas the new species has 3 well-developed distal spines. Only 3 species of the group (G.barbellata, G.robusta, and the new species) have 3 well-developed distal spines on the antennular article 1: G.barbellata and G.profunda both have the metagastric region unarmed, whereas there are 2 spines in the new species. Furthermore, the branchial regions are unarmed in G.barbellata and G.profunda, whereas these regions have some spines in the new species. Finally, the epigastric region is armed with numerous spines in G.barbellata and unarmed in G.profunda, whereas this region has only 2 spines. The rostrum is much larger in the new species than in G.barbellata.
Galatheatukitukimea showed the lowest COI genetic divergence with G.profunda (12.0%). Concerning the other morphologically similar species, the divergences ranged from 16.8% (G.sentosa and G.barbellata), 17.8% (G.bicornis) to 19.9% (G.robusta) (unfortunately no COI data are available for G.echinata).
Coloration.
Dorsal surface of the cephalothorax, chelipeds, and pereiopods white, with yellow-orange blotches bearing red spots. Dorsal regions covered with iridescent setae. Ventral surfaces of pereiopods and chelipeds exhibit the same coloration pattern. Sternite white (Fig. 1).
Galatheatukitukimea sp. nov., holotype, male (MNHNC DEC-15582) and paratype, female (SCBUCN6722). Color in life, dorsal view.
Distribution.
Chilean Exclusive Economic Zone; Rapa Nui Multiple-Use Coastal Marine Protected Area (AMCP-MU Rapa Nui); on the Pukao seamounts; Motu Motiro Hiva Marine Park (MMH Park); around to Motu Motiro Hiva Island (Salas and Gómez Island), at depths of 348–407 m.
Galatheatukitukimea sp. nov., MNHNC DEC-15582, holotype, male. A. Carapace and pleonal tergite 1–4, dorsal view; B. Left antennular and antennal peduncles, ventral view; C. Right P1, the setation of all articles is omitted; palm, merus, and carpus bear long plumose setae on the extensor margin, and the distal region of the manus is finely setose; D–F. Left P2-P4, lateral view; G. Sternal plastron, ventral view; H. Left third maxilliped, ventral view.
Habitat and ecology.
Specimens of G.tukitukimea were living among the perianal spines of the sea urchin Stereocidarisnascaensis (Fig. 3). This urchin is white with a reddish tone at the base of its spines. The sea urchins were found on rocky bottoms, and on the Pukao seamounts, the substrate was covered with red crustose algae at a depth of 348 m.
Galatheatukitukimea sp. nov. on Stereocidarisnascaensis in habitat. A. Pukao seamount, depth 348 m; B. Motu Motiro Hiva Island, depth 407 m; C. G.tukitukimea on the spine of S.nascaensis. Image credit ROV SuBastian/Schmidt Ocean Institute, FKt240224 expedition.
Ocean census species number.
OC186.
Discussion
The discovery of Galatheatukitukimea in the southeastern Pacific represents the first record of the genus from this region of the ocean and from Chilean territory. This finding supports the hypothesis pointed out in other studies (e.g. Gallardo et al. 2021) of a biogeographic connectivity and faunal affinity between the Salas and Gómez Ridge and the subtropical communities of the western and central Pacific. In fact, during the preliminary study of the squat lobsters found in the recent expeditions (e.g. FKT240224) we have observed a clear overlap among the species found in Salas and Gómez and Nazca Ridges and the southwestern and central Pacific fauna (manuscript in preparation).
Sampling carried out in the seamounts of the southeastern Pacific are demonstrating the existence of a little-known fauna (Mecho et al. 2021; Wagner et al. 2021; Gaymer et al. 2025). Likewise, these samples carried out with ROV also allow us to know quite reliably the habitat where many of these species are found (Asorey et al. 2020; Gallardo et al. 2021; Tapia-Guerra et al. 2021; Cañete et al. 2023). The new species found in the seamounts of Salas and Gómez Ridge has always been observed associated with the sea urchin Stereocidarisnascaensis. This sea urchin is distributed in the southern Desventuradas Islands, the intersection of the Nazca and Salas and Gómez Ridges, and along Salas and Gómez ridges (Mecho et al. 2021). This expedition extended the distribution range of S.nascaensis westward, within the Easter Island ecoregion. This potential mimicry-based association is uncommon in squat lobsters, which warrants further study. Previous records of S.nascaensis were obtained using Agassiz trawls (Parin et al. 1997; Mecho et al. 2021), a more aggressive sampling method that may have prevented the simultaneous collection of Galathea specimens. In contrast, the new records were made using the manipulator arm of the ROV SuBastian, a more precise and less disruptive technique.
Most of the symbiotic relationships between squat lobsters and macro-invertebrates have been observed with soft and black corals (see the review by Baeza 2011). In the species of Galatheidae, the association with echinoderms is common among crinoids and species of the genus Allogalathea (Cabezas et al. 2011) and Galathea (Baba and Fujita 2008; Baba et al. 2009). An association between squat lobsters and sea urchins has only been observed between Munidopsisabdominalis (family Munidopsidae) and echinoids of the genus Cidaris (Rice and Miller 1991).
Morphologically, Galatheatukitukimea is related to other species of the genus distributed from Madagascar to New Caledonia and Wallis and Futuna, e.g. G.barbellata, G.bicornis, G.echinata, G.profunda, G.robusta, and G.sentosa (e.g. Baba 1990; Macpherson 2012). These species belong to the same molecular clade (Fig. 4), but only a single gene (COI) is available, so their phylogenetic relationships cannot be known reliably. Most of the morphologically related species inhabit deep waters between 100 and 700 m depth, although most species of the genus Galathea inhabit shallow waters (<100 m) (Baba et al. 2008; Schnabel et al. 2011), which suggests some common ecological characteristics. The genetic divergence in the COI gene between G.tukitukimea and six of these seven species ranges from 12% to 20.5%, which falls within the range previously reported for the genus Galathea (Macpherson and Robainas-Barcia 2015). These genetic distances support the distinctiveness of G.tukitukimea within this group, though further molecular markers and ecological data are necessary to resolve their phylogenetic relationships more robustly. Obviously, more data, both molecular and ecological, are needed to establish a more complete phylogeny of this group of species.
Phylogenetic tree obtained after the maximum-likelihood analysis based in COI sequences.
Supplementary Material
XML Treatment for Galathea tukitukimea
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Allison EC Durham JW Mintz LW (1967) New Southeast Pacific echinoids.Occasional Papers of the California Academy of Sciences 62: 1–23.
- 2Asorey CM Sellanes J Easton EE Bieler R Mecho A (2020) Architectonicakarsteni Rutsch, 1934 (Gastropoda: Architectonicidae) in seamounts of the Nazca-Desventuradas Marine Park: first record in Chilean waters since the Miocene.The Nautilus 134(2): 61–70.
- 3Baba K (1990) Chirostylid and galatheid crustaceans of Madagascar (Decapoda, Anomura). Bulletin du Muséum National d’Histoire Naturelle, Paris, 4e série, Section A 11: 921–975. 10.5962/p.288276 · doi ↗
- 4Baba K Fujita Y (2008) Squat lobsters of the genus Galathea (Decapoda: Anomura: Galatheidae) associated with comatulid crinoids from the Ryukyu Islands, Japan.Crustacean Research 37(0): 43–62. 10.18353/crustacea.37.0_43 · doi ↗
- 5Baba K Macpherson E Poore GCB Ahyong ST Bermudez A Cabezas P Lin CW Nizinski M Rodrigues C Schnabel KE (2008) Catalogue of squat lobsters of the world (Crustacea: Decapoda: Anomura – families Chirostylidae, Galatheidae and Kiwaidae).Zootaxa 1905(1): 1–220. 10.11646/zootaxa.1905.1.1 · doi ↗
- 6Baba K Macpherson E Lin CW Chan T-Y (2009) Crustacean Fauna of Taiwan: squat lobsters (Chirostylidae and Galatheidae). National Science Council (Taiwan), Taipei, [ix +] 312 pp.
- 7Baba K Ahyong ST Macpherson E (2011) Morphology of the marine squat lobsters. In: Poore GCB Ahyong ST Taylor J (Eds) The Biology of Squat Lobsters.CSIRO Publishing, Melbourne and CRC Press, Boca Raton, 1–37.
- 8Baeza JA (2011) Squat lobsters as symbionts and in chemo-autotrophic environments. In: Poore GCB Ahyong ST Taylor J (Eds) The Biology of Squat Lobsters.CSIRO Publishing, Melbourne and CRC Press, Boca Raton, 249–270.