Sustained learned immunosuppression could not prevent local allergic ear swelling in a rat model of contact hypersensitivity
Yasmin Salem, Stephan Leisengang, Marie Jakobs, Kirsten Dombrowski, Julia Bihorac, Laura Heiss-Lückemann, Sebastian Wenzlaff, Lisa Trautmann, Tim Hagernacker, Manfred Schedlowski, Martin Hadamitzky

TL;DR
This study explores how learned immune responses in rats affect allergic reactions, finding that while some immune effects are preserved, local swelling remains unaffected.
Contribution
The study demonstrates that learned immunosuppression preserves some immune effects but fails to prevent local allergic swelling in a rat model.
Findings
Conditioned suppression of splenic cytokine production was preserved.
No conditioned effects were observed in draining lymph nodes or prevention of ear swelling.
Symptoms like itch may better reflect patient disease burden than swelling.
Abstract
Taste-immune associative learning has been shown to mimic immunopharmacological responses. Conditioned pharmacological effects may therefore be considered as controlled drug dose reduction strategy to maintain treatment efficacy. Against this background, the present study applied an established taste-immune associative learning protocol to a rat model of DNFB-induced contact hypersensitivity. After repeated pairings of a saccharin taste (conditioned stimulus, CS) with injections of the immunosuppressant cyclosporine A (CsA, unconditioned stimulus, UCS), animals were sensitized with the hapten. Retrieval started by presenting the CS together with sub-effective doses of CsA. This procedure preserved a conditioned suppression of splenic cytokine production. Compared to full dose treated animals, conditioned effects were neither observed in draining lymph nodes nor did it prevent ear…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
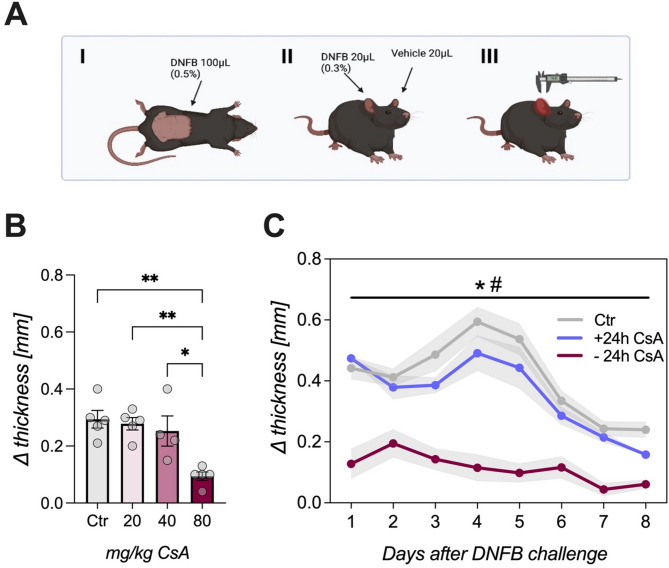 Figure 1
Figure 1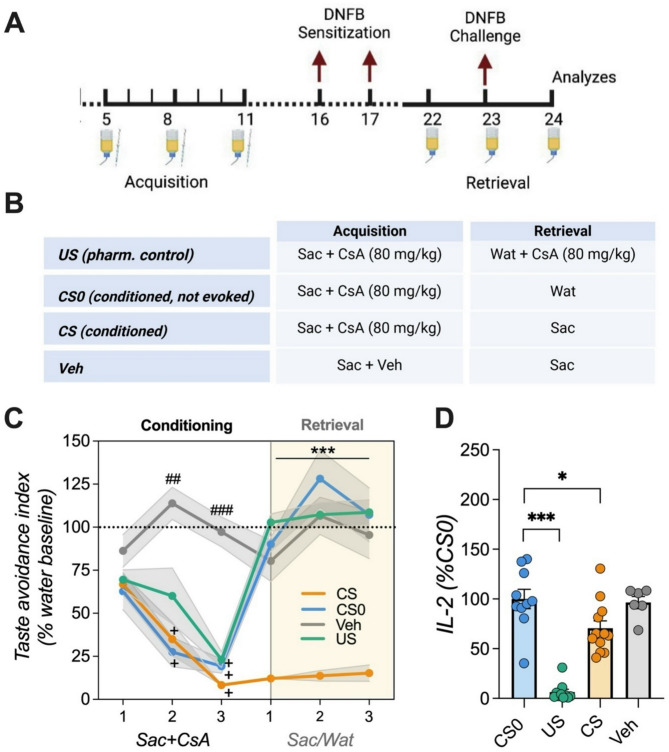 Figure 2
Figure 2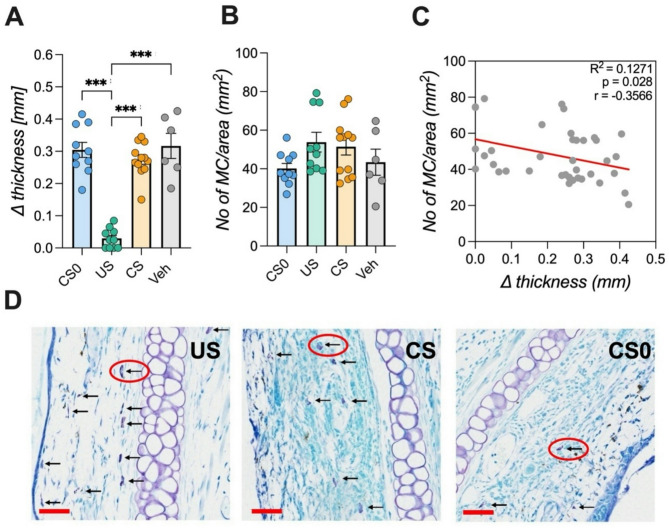 Figure 3
Figure 3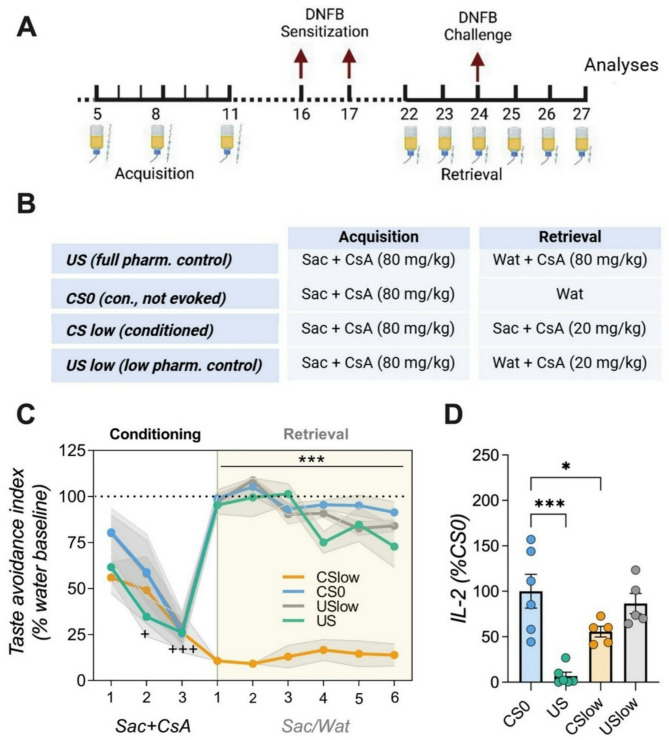 Figure 4
Figure 4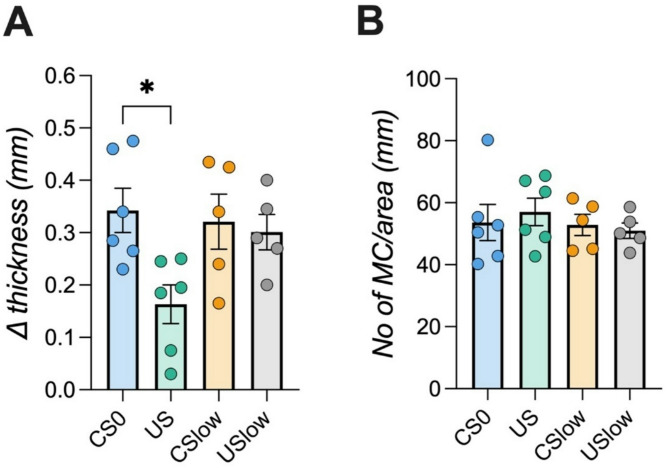 Figure 5
Figure 5- —Universitätsklinikum Essen (8912)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsOlfactory and Sensory Function Studies · Biochemical Analysis and Sensing Techniques · Immune Response and Inflammation
Introduction
Allergic contact dermatitis (ACD) is an inflammatory skin disease triggered by daily used objects such as detergents and fragrances that contain reactive low molecular weight chemicals (haptens)^1^. ACD accounts for 30% of all occupational diseases in industrialized nations^2^ and it is classified as type IV delayed type allergy. Disease progression involves two distinct phases^3^. In an initial sensitization phase, allergen exposure leads to a complex formation with self-proteins (haptenization) and a subsequent activation of the innate immune system. Upon re-exposure to the allergen (challenge phase), various activated immune cells and their mediators induce characteristic symptoms such as erythema, oedema and pruritus, caused by the dilation of blood vessels and increased capillary permeability^3–5^. Specifically, keratinocytes, neutrophils and macrophages secrete cytokines such as tumor necrosis factor (TNF)-α and interleukin (IL)-1), which recruit T cells^6^. These activated T cells, in turn, release other cytokines such as interferon (IFN)-γ, IL-2, IL-17 and IL-5 ^3,7^. Additionally, natural killer (NK) T cells produce IL-4, activating B cells and their secretion of immunoglobulin (Ig) M. This process triggers the complement system and subsequently mast cells, which release TNF-α and histamine^7,8^.
Frequently, the immunosuppressive drug cyclosporine A (CsA) is systemically used to alleviate ACD symptoms in humans but also in rat models of contact hypersensitivity (CHS) by modulating T cell activity^9^. However, chronic CsA use is accompanied by somatic and neuropsychiatric side effects including nephrotoxicity, depression, and/or anxiety^10–12^negatively impacting patients’ quality of life^1^. To overcome these drawbacks, learned immunosuppressive placebo responses have been proposed as a strategy to reduce drug dosages while maintaining therapeutic efficacy^13,14^. These approaches rely on classical conditioning and the intense bidirectional communication between the brain and the immune system^15,16^. Such strategies have been demonstrated effective in experimental animals, healthy subjects and patient populations^17,18^.
In this established paradigm, animals are exposed to an unfamiliar gustatory stimulus as conditioned stimulus (CS, saccharin solution) paired with an injection of an immunosuppressive drug, such as CsA or rapamycin as unconditioned stimulus (UCS). Subsequent presentation of the CS alone at a later time point induces both behavioral and immunological changes. These include conditioned taste avoidance (CTA), where animals consume less of the CS, and a suppression of T cell function alongside decreased cytokine protein production ^20–24^. Importantly, recent findings demonstrated that the administration of low doses of the UCS (10–25%) shortly after CS re-presentation as reminder cues, not only strengthened but also sustained these learned immunosuppressive responses^25,26^. This strategy has been shown to prolong heart allograft survival and to attenuate disease progression in rat models of brain tumor and rheumatoid arthritis^23–25^. However, the potential of this learned drug dose reduction strategy within a CHS model has only sparsely been analyzed^29^. Moreover, findings point out that CHS symptom severity not only depend on the allergen, the animal species and the strain used, but also differs in response to administered CsA drug doses and the allergen which is used^26–32^. Against this background, the present study evaluated the potential effectiveness of sustaining taste-immune associative learning in an allergy-related experimental disease model with the aim of broadening the knowledge regarding this dose-reduction strategy as supportive treatment tool.
Materials and methods
Animals
Male Dark Agouti rats (DA/HanRj; nine to ten weeks old, 190–220 g; Janvier Labs, France) were group housed in a temperature (20° C) and humidity (55 ± 5%) controlled facility under a 12/12-h reversed light/dark cycle (lights off at 7:00 a.m.). After acclimatization for two weeks, rats were housed individually with ad libitum access to food and tap water. The animal facilities and experimental procedures were in accordance with National Institutes of Health and Association for the Assessment and Accreditation of Laboratory Animal Care guidelines and the ARRIVE guidelines and were approved by the Institutional Animal Care and Use Committee (LANUV TSG-Nr. G1884/21 Düsseldorf, North Rhine-Westphalia).
Drugs
A stock solution of the immunosuppressive drug CsA was prepared freshly every day by dissolving 200 mg CsA (LC Laboratories, Woburn, USA) in 200 µL 95% ethanol (Braun, Melsungen, Germany) and 1800 µL Miglyol (Caelo, Hilden, Germany). Corresponding to the animal’s individual weight, this stock solution was further diluted with 0.9% NaCl (Braun, Melsungen, Germany) to gain the respective drug dosages of 20, 40 and 80 mg/kg body weight ^26,27^. The Saccharin solution (10 mM; Sigma Aldrich) was prepared according to previous studies^24,28^.
CHS induction and symptom assessment
The chemical agent dinitrofluorobenzene (DNFB) is a potent contact allergen that induces cutaneous mast cell degranulation^33^. CHS was induced by using DNFB (Sigma-Aldrich, St. Louis, USA) in two phases^30^. In a sensitization phase, the hapten (100 µL of 0.5% DNFB dissolved in 4:1 acetone - olive oil) was applied on the animal’s shaved abdomen on two consecutive days. Following an incubation period of four days, the challenge phase was initiated, where animals were re-exposed to the allergen (20 µL of 0.3% DNFB) on one ear, counterbalanced left-and-right. The corresponding ear serves as control and was treated with a vehicle solution. 24 h later, ear thickness was measured (Fig. 1A). All procedures were performed under anesthesia to prevent stress for the animals (1.5–2% isoflurane mixture with oxygen; Isothesia, Piramal Critical Care B.V., Voorschoten, Netherlands). Ear thickness of the challenged and the control ear were measured at the middle of the ear lobe using a digital caliper (Kroeplin, Germany). Symptomatology of CHS was analyzed as ear thickness difference between both ears (∆ thickness).
Fig. 1. CsA treatment on CHS disease progression. (A) Schematic overview of CHS induction. Animals were sensitized on two consecutive days with DNFB (100 µL 0.5%) on the shaved abdomen. After an incubation time of four days, animals were challenged on one ear with DNFB (20 µL 0.3%), the other ear serves as control ear. 24 h later ear thickness was measured with a digital caliper. (B) Animals were sensitized as described and four days later CsA treatment was initiated for three consecutive days. On the third day, animals were re-exposed to the allergen and ear thickness was measured 24 h later. Treatment with 80 mg/kg resulted in a significant attenuation of thickness difference (ANOVA followed by Bonferroni post hoc analysis, p < 0.05 **p < 0.01; n = 4–5/group). (C) To assess the impact of the starting point of CsA (80 mg/kg) treatment, animals were subjected to continuous treatment regimen at different time points, either pre (-24 h CsA*) or post (+ 24 h CsA) DNFB challenge. Ear swelling was monitored over eight days. Bonferroni post hoc tests revealed at every time point a significantly less ear thickness difference of the − 24 h CsA group compared to the control group (*p < 0.05, n = 5/group), as well as between − 24 h and + 24 h CsA groups (#p < 0.05; n = 5/group). Data are shown as mean ± SEM.
Determining csa dose and treatment start of DNFB-induced CHS symptoms
Prior to the conditioning experiments, groups of rats were injected with three different doses of CsA (20, 40 and 80 mg/kg) intraperitoneally (i.p.) to determine treatment efficacy on CHS symptoms (ear swelling). Animals were sensitized as described and four days later CsA treatment was initiated for three consecutive days. On the third day, animals were re-exposed to the allergen and ear thickness was measured 24 h later (Fig. 1B). To determine the optimal starting point of treatment with CsA (80 mg/kg), different groups of sensitized animals were subjected to a continuous treatment regimen, starting at two different time points: 24 h before the challenge with the allergen (-24 h CsA group) and 24 h after the challenge (+ 24 h CsA). Ear thickness was measured over eight days and compared with a non-treated control group (Fig. 1C).
Behavioral conditioning protocol - experiment 1
Following the establishment of CsA dose and treatment start, the first conditioning experiment was conducted, where new animals underwent a training phase with access to water in the morning (9 a.m.) and evening (5 p.m.) for 15 min. After each drinking session, the bottles were weighed to measure fluid intake. Individual mean water consumption of all sessions over a period of five days was taken as baseline level (100%) for ‘‘normal” fluid intake. After these five training days, acquisition started and animals were randomly allocated into four treatment groups. During three acquisition trials, separated by 72 h, all animals received saccharin and an i.p. injection with CsA (except handling control animals (Veh)) in the morning session for 15 min. In the evening session, all animals received water. Five days after the last acquisition all animals were immunized (induction of CSH as described above) and received water during the morning and evening drinking sessions until day 22 before retrieval was initiated. At retrieval, rats of the conditioned group (CS) received saccharin in the morning sessions and water in the evening. To compare conditioning effects with a standard pharmacological treatment, animals of the US group received i.p. injections of CsA (80 mg/kg) during the morning session of the retrieval phase. Control animals for residual effects of CsA administration (CS0) were neither re-exposed to the saccharin (CS) nor did they receive additional CsA at retrieval. Three retrieval trials were performed with DNFB challenge on the second trial, whereas ear swelling was measured 1 h after the 3rd trial, before ear samples and spleens were collected (Fig. 2A, B). The total amount of liquid consumed was assessed after each morning and evening drinking session by weighing the bottles. Fluid consumption and taste avoidance (CTA) was calculated as a percentage of the individual baseline water consumption (Fig. 3).
Fig. 2. Taste-immune associative learning (A) Schematic representation of conditioning paradigm and (B) group allocation. Animals underwent three acquisition trials. During these trials the CS,* US*, and CS0 groups received saccharin and an injection with CsA (80 mg/kg) prior to sensitization with DNFB. After the incubation time, retrieval started with presentation of the saccharin solution (CS and Veh group) or water (US and CS0 group). The US group additionally received 80 mg/kg CsA. On the second day of retrieval, animals were challenged with DNFB and 24 h later ear thickness was measured. (C) Taste avoidance index. Conditioned animals showed a pronounced CTA on the second and third acquisition day and over the course of retrieval, reflected by a significantly lower saccharin consumption (ANOVA followed by Bonferroni post hoc analysis; ##p < 0.01; ###p < 0.001 = Veh vs. all groups; +++p < 0.001 = all groups vs. acquisition day 1; ***p < 0.001 = CS vs. all groups; n = 6–12/group). (D) IL-2 levels of ex-vivo anti-CD3 stimulated splenocytes. Compared to the CS0 group, US and CS animals showed reduced IL-2 amounts. Data are presented as percentage of CS0 group (ANOVA followed by Bonferroni post hoc analysis; *p < 0.05, ***p < 0.001; n = 6–12/group). Data are shown as mean ± SEM.
Fig. 3. Local CHS inflammation following taste-immune associative learning. (A) Ear thickness was measured 1 h after the last retrieval trial. Animals receiving the full pharmacological dose of CsA (US group) showed significantly attenuated ear swelling compared to the other groups (ANOVA; Bonferroni post hoc analysis, ***p < 0.001; n = 6–12/group). (B) Mast cell infiltration in the ear was counted and standardized to the area. US and CS groups showed a tendency towards higher numbers of mast cells per area but did not reach level of significance. (C) Correlation analysis between ear thickness difference and mast cell infiltration showed a significant negative interaction (Pearsons’s correlation, R squared = 0.127; p = 0.028). Data are shown as mean ± SEM. (D) Representative images of a toluidine-stained ears (5 μm section, 50 μm red scale bar). Mast cells (arrows) and cartilage are visible in violet. Sections of US and CS animals demonstrate a higher amount of mast cell infiltration into the site of local allergic reaction compared to the CS0 group.
Memory-updating of protocol - experiment 2
For the second experiment, new subjects were conditioned as described above using identically treated US and CS0 control groups. Contrary to Experiment 1, the handling control group (Veh) was omitted and two experimental groups (CSlow, USlow) were now included in the protocol. Both groups were conditioned with three acquisition trials receiving saccharin as CS and an i.p. injection with CsA (80 mg/kg) as US as described in Experiment 1. During each of the six retrieval trials, however, the CSlow animals received an i.p. injection with sub-effective CsA (20 mg/kg) as “reminder cues immediately after saccharin exposure in the morning session. The USlow group served for controlling the pharmacological efficacy of the sub-effective doses by receiving water at both drinking sessions together with an injection of CsA (20 mg/kg) in the morning session. Retrieval was conducted over six consecutive days, with a DNFB challenge on the second day. Like in Experiment 1, ear thickness was measured.
ear swelling was measured 1 h after the 3rd retrieval trial, before ear samples and spleens (Fig. 4A, B). The total amount of liquid consumed was assessed after each morning and evening drinking session by weighing the bottles. Fluid consumption and taste avoidance (CTA) was calculated as a percentage of the individual baseline water consumption.
Fig. 4. Taste-immune associative learning with reminder cues. (A) Schematic representation of conditioning paradigm and (B) group allocation. During three acquisition trials (CS/UCS parings), animals of all groups (CSlow, USlow, US, CS0) received saccharin and an injection with CsA (80 mg/kg). Subsequently, animals were sensitized with DNFB. 5 days following sensitization, conditioned animals (CSlow) received saccharin paired with injections of 25% (20 mg/kg) of the full therapeutic CsA dose during retrieval. Control groups received either water together with injections of either 20 mg/kg CsA (USlow group) or 80 mg/kg CsA (US group), or vehicle (CS0 group). Animals were challenged with DNFB on the second day of retrieval. (C) Taste avoidance index. Compared to all other groups, conditioned animals (CSlow) displayed a pronounced CTA, over the course of retrieval (ANOVA followed by Bonferroni post hoc analysis; +p < 0.05, +++p < 0.001 = all groups vs. acquisition day 1; ***p < 0.001 = CSlow vs. all groups; n = 5–6/group). (D) US and CSlow groups also significantly differed in IL-2 production of ex-vivo anti-CD3 stimulated splenocytes compared to CS0. Data are presented as percentage of CS0 group. (ANOVA followed by Dunnett’s post hoc analysis, +p < 0.05; ++p < 0.001; *p < 0.05, ***p < 0.001; n = 5–6/group; Data are shown as mean ± SEM).
Splenocytes isolation
One hour after the last CS re-exposure, deeply anaesthetized animals (isoflurane, 4–5%) were sacrificed by decapitation, spleens were removed and splenocytes were isolated by disrupting the spleen in a Petri dish containing cold HBSS with a 20 ml syringe plunger. Red blood cells were then lysed with BD Pharm Lyse (BD Pharmingen, Heidelberg, Germany) and splenocytes were washed in cell culture medium (RPMI + 10% FBS + 0.1% gentamycin) before being filtered through a 70 μm nylon cell strainer. After washing the suspension, cell concentrations were determined via an automated cell counter (Vet abc, Medical Solution, Steinhausen, Switzerland) and cell numbers were adjusted to a final concentration of 5 × 10^6^ cells/mL. Splenocytes were seeded in a 96 well plate (250.000 cells/well) and stimulated with 1 µg/mL of mouse anti-rat CD3 monoclonal antibody (clone: G4.18, BD Biosciences) for 24 h in a humidified incubator (37 °C, 5% CO_2_).
IL-2 ELISA
IL-2 amounts of the supernatants were measured using a sandwich ELISA (Quantikine^®^ELISA Rat IL2, R&D systems, Minneapolis, USA) according to the manufacturer’s instructions. Optical density was assessed at 540 and 570 nm using Fluostar OPTIMA Microplate Readers (BMG Labtech, Offenbach, Germany). Cytokine concentrations were calculated using a log-log curve-fit standard curve.
Histology
Histological analysis of the challenged ear was performed to quantify the mast cell infiltration. Ears were collected, cut longitudinally, and fixed in 4% paraformaldehyde for one week. Tissues were embedded in paraffin and cut serially into slices of 5 μm thickness using a microtome. Sections were stained with 0.01% toluidine blue for one minute and scanned using Scope Scanner (Aperio CS2 Scanner, Leica Biosystems, Deer Park, USA). Three sections per animal with a total of nine areas were counted using ImageJ (version: 2.9.0/1.53t; National Institutes of Health) and evaluated in a blinded manner.
Statistical analysis
Statistical analyses were performed using SigmaPlot (Version 12.3, Systat Software San Jose, CA, USA) and Prism (Version 9.5.1, Graph Pad Software, San Diego, CA, USA). Normality was tested via Shapiro-Wilk-Test and data were log transformed when necessary (Experiment 1). P value was considered significant at < 0.05. Two-way analysis of variance (ANOVA) with group (treatment) as one factor and time (days) as a within-subjects factor was used to analyze behavioral data as well as ear thickness measurements. When appropriate, post-hoc individual comparisons between groups were determined by Bonferroni t-tests. One-way ANOVA followed by Bonferroni or Dunnett’s Method post hoc analysis was used for all other experiments. The numbers of animals per treatment group, which differ marginally due to technical reasons, are reported in the figure legends.
Results
Pre-treatment with high doses of csa attenuates CHS symptoms
To determine the effective dose of CsA for preventing allergic DNFB-induced ear swelling, a dose response study was conducted, revealing differences between the applied dosages (ANOVA: F(3,15) = 9.552; p < 0.001; η^2^=0.656). Only animals treated with 80 mg/kg CsA for three consecutive days showed a significant attenuation of ear swelling compared to controls and animals treated with 20 or 40 mg/kg CsA (p < 0.05; Fig. 1B). Subsequently, different time points for a therapeutic intervention with 80 mg/kg CsA were examined by observing ear swelling over eight days. ANOVA revealed a main effect for the factor time (F(7,112 = 54.24; p < 0.001 η^2^=0.832), group (F(3,112) = 23.02; p < 0.001; η^2^=0.772) and a time x group interaction (F(21,112) = 4.734; p < 0.001; η^2^=0.470). Notably, only pre-treatment that started 24 h before challenge led to a significantly diminished ear thickness compared to untreated controls and the group receiving CsA treatment 24 h post challenge (Ctr vs. -24 h CsA, p < 0.05; + 24 h CsA vs. – 24 h CsA, p < 0.05). In control animals, ear swelling peaked on day four (Fig. 1C).
Taste-immune associative learning in CHS
In a first setup (Experiment 1), animals underwent three conditioning sessions where a saccharin solution (CS) was paired with CsA injections (UCS; 80 mg/kg), before being immunized with DNFB on two consecutive days. Since pre-treatment (24 h before immunization) was necessary to prevent ear swelling, retrieval of conditioned immunosuppression began one day prior to DNFB challenge (Fig. 2A, B). Conditioned animals showed a pronounced CTA across all three retrieval trials (ANOVA: for group (F(3,68) = 80.486; p < 0.001; η^2^=0.944) and time (F(2,68) = 6.375; p < 0.01; η^2^=0.158), reflected by significantly diminished fluid intake in the CS group (Fig. 2C; p < 0.001). Concurrently, IL-2 cytokine production of ex-vivo anti-CD3 stimulated splenocytes differed significantly among groups (ANOVA F(3,34) = 33.54; p < 0.001; η^2^=0.747; Fig. 2D). Post-hoc analysis revealed that IL-2 levels in conditioned (CS) and fully treated (US) animals were significantly lower than in the CS0 group (p < 0.05, p < 0.001). Analysis moreover revealed significant groups differences of allergic symptoms (ANOVA: F(3,34) = 44.46; p < 0.001; η^2^=0.797), whereas only CsA treated animals treated exhibited reduced ear swelling (p < 0.001; Fig. 3A). Histological staining showed a tendency towards higher numbers of mast cell infiltration at the inflammation site in US and CS groups compared to the controls but did not reach level of significance (Fig. 3B, D). However, correlation analysis demonstrated a significant negative interaction between ear thickness and mast cell density (two-tailed t-test, p = 0.028; Fig. 3C). Analysis of immune cell subsets such as B cells, MHC-II + cells, and APCs in cervical and axillary lymph revealed reductions only in the US but not conditioned group (Supplementary Fig. 1).
Memory-updating of taste-immune associative learning boosts systemic immunosuppression but does not affect local CHS symptoms
In the second setup (Experiment 2), animals underwent three conditioning trials using CsA (80 mg/kg) as UCS and saccharin as CS before being immunized with DNFB. Similar to Experiment 1, retrieval began one day prior to DNFB challenge (Fig. 4A). However, conditioned animals in the CSlow group received saccharin paired with injections of 25% (20 mg/kg) of the full therapeutic CsA dose. Control groups received water together with injections of either 20 mg/kg CsA (USlow group*)* or 80 mg/kg CsA (US group), or vehicle (CS0, Fig. 4B). ANOVA revealed significant group (F(3,90) = 152.634; p < 0.001; η^2^=0.906) and time effects (F(5,90 = 3.78; p < 0.01; η^2^=0.174) at retrieval. The CSlow group exhibited a robust CTA over all six retrieval trials as reflected by significantly reduced fluid intake (p < 0.001; Fig. 4C). Systemic IL-2 cytokine levels were also affected (F(3,18) = 12.85; p < 0.001; η^2^=0.682) with significantly reduced IL-2 levels in the CSlow and US (80 mg/kg CsA) groups compared to CS0 animals *(*p < 0.05, p < 0.001; Fig. 4D). In line with Experiment 1, reduced ear swelling reduction was observed only in the US group but not in conditioned animals (ANOVA: F(3,18) = 4.02; p < 0.05; η^2^=0.395; Fig. 5A). Furthermore, local immune cell infiltration did not differ significantly between groups (Fig. 5B).
Fig. 5. Local CHS inflammation following taste-immune associative learning with reminder cues. (A) Ear thickness measurement revealed significant diminished swelling in US animals compared to the CS0 group (ANOVA followed by Bonferroni post hoc analysis, *p < 0.05). (B) Mast cell infiltration into the inflamed ear was analyzed and no differences were observed. Data are shown as mean ± SEM (n = 5–6/group).
Discussion
The present study investigated the potential of taste-immune associative learning with CsA in a rat model of CHS. Compared to controls, we demonstrated that conditioned immunopharmacological responses with CsA, reflected by suppressed level of the proinflammatory cytokine IL-2, were preserved when 25% of the drug dose (20 mg/kg CsA) was applied at retrieval alongside with the CS (saccharin). However, this conditioned immunosuppression was insufficient to prevent the emergence of local allergic ear swelling.
The chronic use of small-molecule immunosuppressants can lead to significant side effects^11^, highlighting the urgent need for alternative or supportive treatments options in immunological diseases. Based on the intense crosstalk between the central nervous system and the peripheral immune system ^34,35^, conditioning of immunopharmacological effects via associative learning has been suggested as means to reduce drug dosages and associated side effects, while maintaining treatment efficacy^15^. Numerous experiments in rodents document the potential clinical relevance of taste-immune associative learning protocols by reporting symptom alleviation in various animal disease models^16^. Against this background, we applied an established paradigm of conditioned immunosuppression with CsA in a DNFB-induced CHS rat model. One major difference from previous studies was, however, the necessity of using a high dose of CsA (80 mg/kg) to prevent DNFB-induced ear swelling, compared to the standard 20 mg/kg. This requirement aligns with other studies reporting the need for high doses in similar contexts ^31,36–39^. Consequently, the taste-immune associative learning paradigm was modified to incorporate high dose CsA as UCS. While earlier studies exclusively employing 20 mg/kg CsA for taste-immune associative learning, our findings showed that conditioned animals exhibited pronounced CTA and reduced IL-2 cytokine levels in ex vivo CD3-stimulated splenocytes. Nonetheless, analysis of the local reaction site revealed no significant impact of immune conditioning. Despite a trend towards higher mast cell infiltration, only the pharmacological group treated with 80 mg/kg CsA exhibited reduced ear thickness.
CsA is known to inhibit mast cells’ histamine release^40–42^, potentially accounting for the higher cell count observed in these groups, whereas the reduced numbers in controls might reflect a degranulated stage. Given that histamine release contributes to vasodilation-dependent oedema^43,44^its inhibition in CsA treated animals likely resulted in diminished ear thickness. However, in conditioned animals the trend towards higher of mast cell numbers as insufficient to reduce swelling, likely due to the complex immune responses involved in CHS, which comprise various cytokines and immune cells^1^. Analyses of these mediators showed reductions in CsA treated but not conditioned animals (data not shown). Interestingly, a similar study using a dinitrochlorobenzene (DNCB)-induced CHS model reported a conditioned reduction in ear swelling with only 20 mg/kg CsA^29^. This discrepancy underscores the allergen-dependent variability in allergic responses ^45^. Thus, DNFB-induced reactions may have been too severe to respond to conditioned immunosuppressive effects at the local site of allergic reaction.
Conditioned responses are known to weaken over time with repeated exposure to the CS without the UCS ^46^. To address this, previous studies introduced a non-invasive memory-updating approach where low or sub-effective doses of the drug used as UCS are paired with the CS during retrieval, preventing extinction of the learned immune memory^25,26^. This approach has demonstrated extended graft survival in transplantation^26^reduced rheumatoid arthritis symptoms ^27^ and attenuated glioblastoma disease progression^28^ in animals. In the present study, 25% of the initial drug dose was administered in conditioned animals together with the CS during retrieval, which sustained CTA as well as conditioned suppressed cytokine level over six retrieval trials. Nevertheless, allergic ear swelling and allergic histological markers remained unaffected. The lack of histological differences might reflect the timing of analysis, as the allergic reaction may have entered a resolution phase by day four post-challenge^1^. The unaltered symptoms could be attributed to the intensity of the DNFB-induced allergic response, which may have been too strong for conditioned immunosuppressive effects to overcome. Similar findings were observed in a murine model of autoimmune uveitis, where conditioned reductions in IL-2 cytokine levels did not translate to symptom improvement^47^ It was suggested that active sensitization might mask conditioned symptom reduction, and splenocyte transfer from conditioned to naïve animals could help reveal these effects. Such approaches may also clarify the results of the current study. It is important to note that CsA is not a first-line treatment for CHS, nor is pre-treatment a standard clinical approach. Thus, future studies should investigate the use of alternative drugs, such as dexamethasone, and explore adjusted treatment schedules. Additionally, subjective symptoms like itch, which better reflect patient burden, should be prioritized over tissue swelling. Previous studies have shown reduced itch despite unaffected wheal sizes ^48^.
Taken together, the present study confirms that conditioned immunosuppression can be preserved by using a memory-updating approach. However, despite a strengthened CS-UCS association CHS symptomatology and histology were unaffected in conditioned animals. These results highlight the need to further investigate the underlying mechanisms and clinical applicability of taste-immune associative learning approaches to optimize its responsiveness towards disease-specific symptoms and to enhance its translational potential.
Supplementary Information
Below is the link to the electronic supplementary material.
Supplementary Material 1
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Castanedo-Tardan, M. P. & Zug, K. A. in Lowell A. Goldsmith et al. (eds.) Fitzpatrick’s Dermatology in General Medicine. The Mc Graw-Hill Companies: UK (2012).