In vitro study of probiotic Lactobacillus helveticus: Antibacterial effects on Porphyromonas gingivalis
Husna Hazirah Bakri, Syarifah Nur Syed Abdul Rahman, Adyani Azizah Abd Halim, Nor Adinar Baharuddin, Hasnah Begum Said Gulam Khan, Zamirah Zainal Abidin, Mohd Hafiz Arzmi, Elly Munadziroh, Wan Himratul Aznita Wan Harun

TL;DR
This study shows that the probiotic Lactobacillus helveticus can inhibit the growth of Porphyromonas gingivalis, a harmful bacteria linked to gum disease.
Contribution
The study demonstrates the antibacterial effect of Lactobacillus helveticus against Porphyromonas gingivalis using multiple experimental methods.
Findings
L. helveticus significantly reduced P. gingivalis growth in a dose-dependent manner.
Biofilm biomass of P. gingivalis was reduced by up to 85% at high L. helveticus concentrations.
Treated P. gingivalis cells showed decreased viability and morphological changes.
Abstract
Probiotics are gaining attention for their benefits as a supplement to improve oral health. This study aimed to evaluate the antibacterial effect of the probiotic Lactobacillus helveticus against Porphyromonas gingivalis, a significant pathogen in periodontal diseases. Antibacterial susceptibility was assessed using the well diffusion assay, with 0.12% chlorhexidine (CHX) served as the positive control. Biofilm biomass was evaluated using crystal violet staining. Cell viability in P. gingivalis treated with L. helveticus was determined using the LIVE/DEAD Baclight bacterial assay via fluorescence microscopy. Ultra-morphological alterations in these cells were further examined using Field Emission Scanning Electron Microscopy. The results indicated that L. helveticus significantly reduced the growth of P. gingivalis. The highest concentration of 109 cells/mL achieved the most substantial…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Fig 1
Fig 1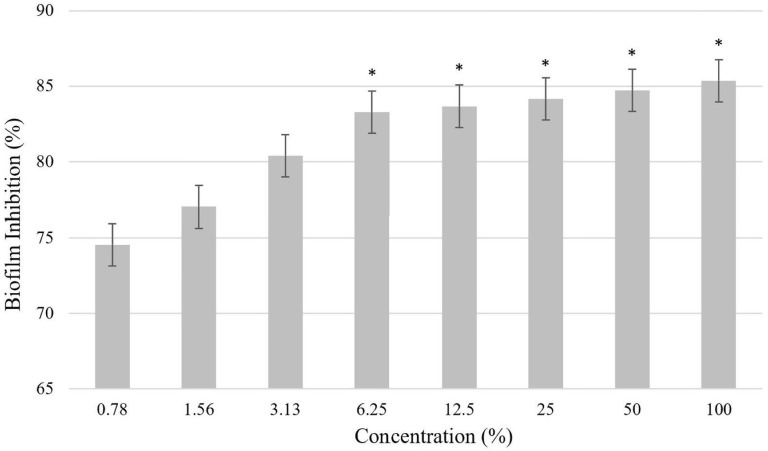 Fig 2
Fig 2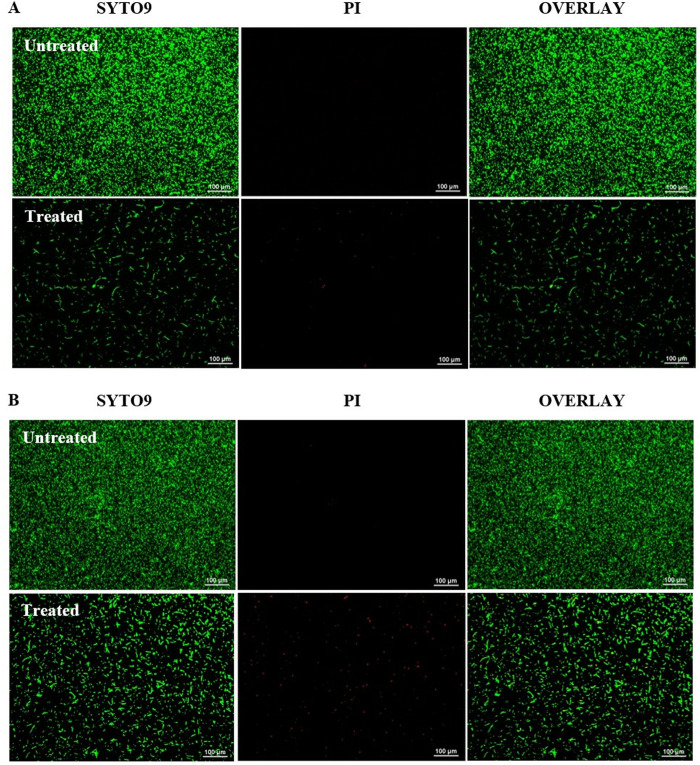 Fig 3
Fig 3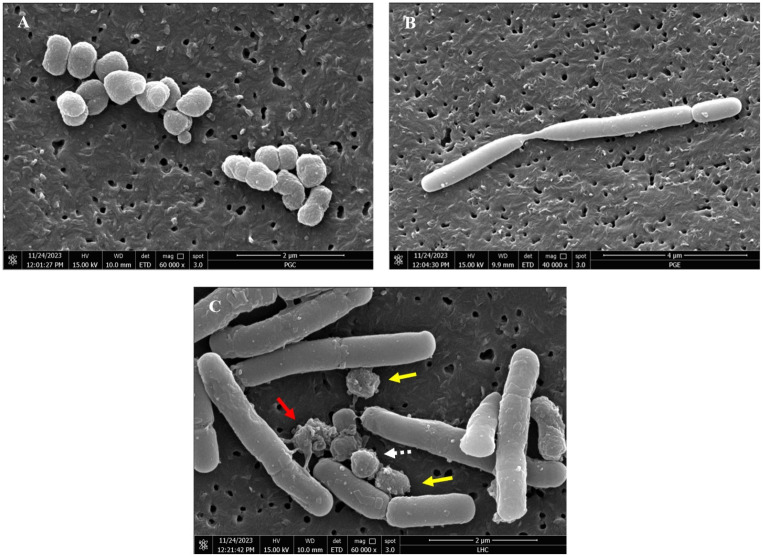 Fig 4
Fig 4- —http://dx.doi.org/10.13039/501100002385Ministry of Higher Education
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsOral microbiology and periodontitis research · Probiotics and Fermented Foods · Antimicrobial Peptides and Activities
1. Introduction
Periodontitis is one of the most common inflammatory diseases affecting all age groups, but it has a higher prevalence in the elderly population [1]. Periodontitis has a prevalence rate of about 50% worldwide and most commonly occurs in adulthood. It is defined as a chronic inflammatory disease of the supporting tissues of the teeth of infectious origin [2]. The supportive tissues of the teeth that are affected by inflammation include the cementum, periodontal ligament, alveolar bone and gingiva surrounding the tooth [3,4]. Inflammation in these areas not only contributes to tooth loss but has also been associated with various systemic diseases, including diabetes, atherosclerosis, rheumatoid arthritis, Alzheimer’s disease, gastrointestinal disorders, and low birth weight in newborns [5]. Periodontitis is characterised by inflammation of the gingiva, which is associated to the buildup of dental plaque. This condition may progress to include inflammation of the bone and periodontal ligament, resulting in the formation of periodontal pockets, a key feature of the disease [6].
Periodontitis is characterised as a dysbiotic disease marked by a shift in the subgingival microbial communities within periodontal pockets, from predominantly Gram-positive aerobic bacteria to a dominance of Gram-negative anaerobes. Notably, the “red-complex” bacteria, which includes Porphyromonas gingivalis, Tannerella forsythia, and Treponema denticola, are significant contributors to this dysbiosis [7]. Among these, P. gingivalis is recognised as a primary causative agent closely associated with the development of periodontitis [8]. P. gingivalis is an obligate anaerobe that thrives in low-oxygen environments and has evolved various mechanisms that allow it to colonise beneath the gingiva, evade the host immune response, and cause damage to oral tissues. Its pathogenicity is enhanced by several virulence factors, including fimbriae, haemolysin, haemagglutinin, capsules, outer membrane vesicles, lipopolysaccharides, and gingipains. Furthermore, P. gingivalis has been shown to form biofilms, which can protect it from host immune responses and enhance its ability to persist within the oral environment [1]. Fimbriae are considered a critical factor in the onset of periodontitis. These surface structures consist of fimbrillin subunits [9], and are essential for various processes, including biofilm formation, auto-aggregation, co-aggregation with other oral bacteria, adhesion to host molecules, and invasion of host cells [10]. These invasion mechanisms contribute to the resorption of alveolar bone, the destruction of surrounding periodontal tissues, and an increased risk of developing systemic diseases associated with periodontitis [11].
Lactic acid bacteria (LAB) are non-spore-forming, Gram-positive, non-respiratory probiotics that generate lactic acid through the fermentation of carbohydrates. Lactobacillus species, widely recognised as probiotics and integral members of the human microflora across various sites, including the oral cavity, have gained increasing interest for their ability to inhibit the growth of numerous pathogens [12]. Recently, probiotics have garnered increasing attention for their potential benefits in improving oral health. Numerous studies have demonstrated the ability of probiotics to inhibit oral pathogens, either through live cultures or cell-free supernatants [13,14]. Among these, L. helveticus, one of the several species of lactic acid bacteria, has demonstrated several probiotic properties in previous in vitro studies, such as resilience in the gastrointestinal tract, adhesion to epithelial cells, and the capability to antagonize pathogenic microorganisms [15,16]. Apart from that, a previous study has reported that L. helveticus has been shown to reduce the increase of pro-inflammatory cytokine expression induced by P. gingivalis stimulation in gingival epithelial cells [17]. The antibacterial properties of probiotics are attributed to their production of organic acids, their ability to adhere to pathogens, and their capacity to decrease bacterial adherence. Furthermore, studies have shown that Lactobacilli produce various bactericidal compounds [18]. Evidences suggest that the antibacterial activity of Lactobacillus strains primarily stems from the production of organic acids and bacteriocins [19,20].
Conventional antibacterial treatments, such as cefazolin and ampicillin, face significant bacterial resistance [21]. Therefore, developing a safe and effective therapeutic agent for oral health is considered vital. In response, probiotics have emerged as a potential defence against antibiotic resistance in severe periodontitis [22]. Effective probiotics must meet certain criteria, including acid and bile resistance, surface adherence, and inhibitory activity against pathogens, all of which are crucial for managing of gingival and periodontal diseases [23]. While various studies have highlighted the inhibitory effects of probiotics on periodontal pathogens, the research specifically focusing on L. helveticus in relation to periodontal disease-associated pathogens remains limited. To address this gap, the objective of the present study is to investigate the potential of L. helveticus ATCC 15009 strain as a probiotic for promoting oral health, focusing specifically on its antibacterial activity against P. gingivalis, a main causative anaerobe bacteria implicated in periodontal disease. Antibacterial susceptibility was assessed using the well diffusion assay, while biofilm biomass was evaluated using crystal violet staining. Cell viability in P. gingivalis treated with L. helveticus was determined using the LIVE/DEAD assay, employing fluorescence microscopy analysis. Furthermore, ultra-morphological alterations in the treated bacterial cells were examined using FESEM. By investigating this relationship through in vitro examination, the study aims to assess the probiotic potential of L. helveticus for preventing or mitigating periodontal disease, which ultimately contributes to improved oral health.
2. Materials and methods
2.1. Bacterial strains and culture conditions
L. helveticus ATCC 15009 from the working stock culture was sub-cultured on MRS agar (de Man, Rogosa, Sharpe) for 24 hours at 37°C [24]. Meanwhile, P. gingivalis ATCC 33277 was sub-cultured from the working stock culture on supplemented BHI-T blood agar plates (Tryptone Soya agar & Brain Heart Infusion broth), enriched with 5% defibrinated sheep blood, hemin (5.0 mg/mL), menadione (5.0 mg/mL), and cysteine (0.5 g/mL) for 48–72 hours at 37°C under anaerobic conditions, using an anaerobic jar with an Anaerogen gas pack (Thermo Scientific™ Oxoid AnaeroGen 3.5L) [25,26].
Following the initial growth, a loopful of colonies from the agar was transferred into 20 mL MRS broth (de Man, Rogosa, Sharpe) and BHI-T broth for L. helveticus and P. gingivalis, respectively. The inoculated broth was then incubated for another 24 hours and 48 hours at 37°C under anaerobic conditions, which promoted bacterial multiplication. Subsequently, the bacterial cells were harvested by centrifugation at 10,000 × g for 10 min at 4°C and washed three times with phosphate-buffered saline (PBS). The L. helveticus cells were then resuspended in MRS broth, and the bacterial concentration was adjusted spectrophotometrically, which is equivalent to cell densities of approximately 10^7^, 10^8^, and 10^9^ cells/mL based on OD_600_ readings [27–29]. The P. gingivalis cells were resuspended in BHI-T broth and was adjusted to optical density (OD_600_) of 0.1 using a UV-VIS spectrophotometer, which is equivalent to approximately 10^8^ colony-forming units per millilitre (cells/mL) [30]. Fresh P. gingivalis and L. helveticus cultures were Gram-stained for confirmation before being employed in all relevant studies.
2.2. Well diffusion assay
The antibacterial activity was tested using the well diffusion method. A sterile cotton swab was employed to evenly distribute 100 µL of P. gingivalis bacterial suspension (10^8^ cells/mL) across the surface of the supplemented BHI-T blood agar [31]. Then, the inoculated agar was punctured with a cork borer to create several wells, each with a diameter of 6.0 mm. For the treatment, each well was filled with 30 µL of various concentrations of the L. helveticus probiotic suspension (10^7^, 10^8^, and 10^9^ cells/mL). The MRS broth served as the negative control, while 0.12% chlorhexidine gluconate (CHX) was used as the positive control. Subsequently, the agar plates were incubated under anaerobic conditions at 37°C for 72 hours. Following this incubation, the presence or absence of a clear zone around the wells was observed. The diameters of the inhibition zones were measured in millimetres (mm) [32]. Larger inhibition zones indicate greater efficacy, reflecting the ability of the probiotic L. helveticus to inhibit P. gingivalis.
2.3. Assessment of biofilm activity
Biofilm biomass was determined using crystal violet staining, utilising the broth microdilution method with minor modifications [33]. In a 96-well plate, 100 µL of P. gingivalis suspension was added to each well, followed by treatment with 10^9^ cells/mL of probiotic L. helveticus suspension at two-fold serial dilutions ranging from 100% to 0.78%. The blank assay consisted of L. helveticus suspension at varying concentrations without any bacterial culture, while wells containing the mixture of BHI-T broth and P. gingivalis suspension served as negative controls. Additionally, a 0.12% CHX solution was utilised as a positive control. The plates were incubated for 48 hours under anaerobic conditions. Then, the wells were washed three times with sterile PBS. Adherent biofilms were subsequently treated with methanol for 15 minutes, stained with 0.04% (w/v) crystal violet for an additional 15 minutes, and rinsed with deionised water. Finally, 95% ethanol was added to each well, and the optical density (OD) values were measured at 550 nm.
The percentage of biofilm inhibition against P. gingivalis at different concentrations of L. helveticus suspension was calculated using the following formula [34]:
where AC indicates the absorbance of the well with treated P. gingivalis (with probiotic) and A0 indicates the absorbance of the control well with untreated P. gingivalis (without probiotic).
2.4. Live/dead bacterial cell viability assay
In this experiment, a probiotic suspension of L. helveticus (10^9^ cells/mL) was added to the wells containing a bacterial suspension of P. gingivalis (10^8^ cells/mL) in a 12-well plate, at a volume ratio of 1:1 (v/v) [35]. The samples were incubated at 37°C for two different time points, 24 and 48 hours under anaerobic conditions. Following the incubation period, the samples were centrifuged for 10 minutes at 10,000 × g to pellet the cells, which were then resuspended in supplemented BHI-T broth. Subsequently, 3 µL of the dye mixture (SYTO9 and PI) from the LIVE/DEAD® BacLight™ Kit was added to the bacterial suspension. The dye mixture was then incubated at room temperature in the dark for 15 minutes. Then, an aliquot of the stained cell suspension (5 µL) was dropped onto a glass microscope slide to analyse the morphological changes in both treated and untreated P. gingivalis biofilms using a fluorescence microscope (Nikon Eclipse Ti-E Inverted Microscope).
2.5. Analysis of morphological alterations in bacterial cells via Field Emission Scanning Electron Microscopy (FESEM)
Briefly, the bacterial suspension of P. gingivalis (10^8^ cells/mL) was exposed to the probiotics L. helveticus (10^9^ cells/mL) in a 12-well plate, maintaining a 1:1 (v/v) ratio and incubated under anaerobic conditions for 24 hours at 37°C [35]. Following incubation, the suspensions were fixed with 4% glutaraldehyde for 4 hours, rinsed twice in buffer for 10 mins each, and then fixed in 1% osmium tetraoxide for 1 hour. After another rinse with double-distilled water (DDH_2_O) for 10 mins, the samples were dehydrated in increasing concentrations of ethanol (30%, 50%, 70%, 80%, 90%, 95% and 100%) for 15 mins each. The samples were then dehydrated for 15 mins in three different ratios of ethanol to acetone: 3:1, 1:1, and 1:3. The samples were subsequently fixed in pure acetone twice for 20 mins each and dried using critical point drying (CPD) for 1–3 hours. Finally, the samples were mounted on stubs with carbon glue, sputter-coated with gold, and observed using a field emission scanning electron microscope (FESEM).
2.6. Statistical analysis
The tests were conducted in triplicate, and the mean ± standard deviation (SD) was calculated. Statistical analyses were carried out using SPSS software (IBM Corp., version 27.0, Armonk, NY, USA). The Shapiro-Wilk test indicated that the data for all experiments did not follow a normal distribution (p < 0.05). To compare data between groups, the non-parametric Kruskal-Wallis test was used, followed by Dunn’s post-hoc test. All analyses were considered statistically significant with p-values < 0.05.
3. Results
3.1. Effects of L. helveticus on the growth inhibition of P. gingivalis
According to the results presented in Table 1 and illustrated in (Fig 1), L. helveticus at concentrations of 10^7^ cells/mL, 10^8^ cells/mL and 10^9^ cells/mL effectively inhibited the growth of P. gingivalis in a dose-dependent manner. The 10^9^ cells/mL concentration exhibited the largest inhibition zone (14.69 ± 0.78 mm), followed by 10^8^ cells/mL (11.35 ± 0.45 mm) and 10^7^ cells/mL (9.23 ± 0.93 mm). The negative control showed no inhibition. These results indicate that the growth of P. gingivalis was most effectively inhibited at 10^9^ cells/mL concentration of L. helveticus, leading to its selection for further analysis.
Table 1: Measurement of the inhibition zone diameters of L. helveticus against P. gingivalis in the BHI-T blood agar well diffusion assay.
Effect of L. helveticus at various concentrations on P. gingivalis.A 0.12% chlorhexidine (CHX) solution served as the positive control, and MRS broth was employed as the negative control.
3.2. Quantification P. gingivalis biofilm biomass inhibition by L. helveticus using the crystal violet assay
The overall biomass of P. gingivalis biofilms was quantified using the crystal violet assay, employing two-fold serial dilutions of L. helveticus from a stock solution of 10^9^ cells/mL. The results of the biomass analysis are shown in (Fig 2). The L. helveticus suspension exhibited significant inhibitory activity against the formation of P. gingivalis biofilms. Notably, at the highest concentrations of L. helveticus, the treated P. gingivalis biofilm displayed a substantial biomass reduction of 85%, as compared to the control group. Hence, these findings demonstrate that L. helveticus possesses bacteriostatic properties.
*Inhibition of P. gingivalis biofilm biomass by L. helveticus.The untreated P. gingivalis biofilm served as the negative control in this study. Experiments were conducted in triplicate and values are represented as mean ± standard deviation. Statistical significance was assessed utilising the Kruskal-Wallis test (p-value < 0.05) along with Dunn’s post-hoc analysis, with a significance level set at p < 0.05 when compared to the control group.
3.3. Evaluation of live/dead cell viability in P. gingivalis biofilms via fluorescence microscopy
Fluorescence microscopy was used to analyse the live/dead cell viability of P. gingivalis biofilms after 24 and 48 hours of incubation with L. helveticus. Both untreated and treated biofilm samples were stained with the LIVE/DEAD Bacterial Viability Kit, which employs SYTO9 as a green-fluorescent nucleic acid stain and propidium iodide (PI) as a red-fluorescent nucleic acid stain. Cells with intact membranes were stained green (SYTO-9) and classified as viable, whereas those with compromised membranes were stained red (PI) and regarded as dead cells. The overlay images from the assay at 24 hours (Fig 3A), revealed that the untreated P. gingivalis biofilm exhibited a highly uniform distribution and dense structure in the absence of L. helveticus. Conversely, treatment with L. helveticus (10^9^ cells/mL) resulted in significantly dispersed and markedly sparse biofilm structures. The observation indicates that this concentration inhibited P. gingivalis biofilm formation, leading to a less dense biofilm.
LIVE/DEAD viability staining of untreated and treated P. gingivalis biofilm with 109 cells/mL of L. helveticus, as revealed under fluorescence microscopy.The biofilm images are viewed after (A) 24 hours and (B) 48 hours of incubation. Viable (live) bacteria cells are green, whereas non-viable (dead) cells are red (Magnification: 10x, scale bar: 100 μm).
At 48 hours, a similar trend was observed in the control group, where the biofilm maintained a highly uniform distribution and a dense structure, suggesting increased bacterial growth in the untreated P. gingivalis (Fig 3B). Conversely, treated P. gingivalis biofilm cells showed an increase in cell numbers during the extended incubation period. Consequently, the structure of the treated P. gingivalis biofilm appeared moderately scattered and diffuse. Furthermore, a higher number of dead cells were observed in the treated samples, highlighting the detrimental impact of L. helveticus on P. gingivalis viability. This evidence collectively demonstrates the significant effects of L. helveticus on the morphology and viability of P. gingivalis biofilms over time.
3.4. The ultra-morphological effects of L. helveticus on P. gingivalis via FESEM
The morphological changes in treated and untreated P. gingivalis with L. helveticus were assessed using FESEM. The untreated P. gingivalis cells were spherical and regular in shape, with smooth surfaces and intact membranes (Fig 4A). In contrast, the viable (negative control) L. helveticus cells had smooth surfaces and were rod-shaped with intact structures (Fig 4B). However, after treatment with 10^9^ cells/mL of L. helveticus, the P. gingivalis cells showed disrupted structures, forming clumps of damaged cells. This observation indicates that P. gingivalis exhibited membrane disruption and irregular cell morphology compared to the untreated cells (Fig 4C).
FESEM images of (A) non-treated P. gingivalis, (B) non-treated L. helveticus and (C) treated P. gingivalis with 109 cells/mL concentration of L. helveticus.Red arrows: Distortion of cell walls. Yellow arrows: Blisters formation. White dotted arrow: Unaffected P. gingivalis cells. Magnification: 60kx (left, below), 40kx (right).
Discussion
The diseases of periodontitis, gingivitis, and dental caries are caused by microorganisms in the oral cavity. Among these, periodontitis, is a prevalent condition, predominantly associated with Gram-negative bacteria, such as P. gingivalis, Aggregatibacter actinomycetemcomitans (A. actinomycetemcomitans), T. denticola, and T. forsythia [36]. Notably, P. gingivalis is recognised as one of the most critical oral pathogens involved in the development of periodontal disease, owing to its various virulence factors that facilitate biofilm formation and contribute to persistent infections in the gingival region [37]. While a substantial literature addresses bacterial biofilms and their effects on periodontal health, further research is needed to fully understand their interactions and explore potential therapeutic strategies. Probiotics, particularly L. helveticus, have garnered interest for their oral health benefits. This study investigates the in vitro antibacterial effects of probiotic L. helveticus ATCC 15009 strain against P. gingivalis, providing a preliminary assessment that could guide future therapeutics.
Our study findings revealed a significant inhibitory effect of L. helveticus against P. gingivalis. The antibacterial activity of P. gingivalis was assessed using blood agar, which serves as both a differential and enriched medium. The inclusion of blood in this medium provides essential nutrients for the cultivation of fastidious organisms like P. gingivalis. Treatment with the highest concentration of L. helveticus, 10^9^ cells/mL demonstrated the largest inhibition zone diameters, indicating that L. helveticus displays antibacterial activity towards P. gingivalis. Similar to our findings, previous study has reported this strain was among the Lactobacillus sp. that has shown inhibitory activity against Gram-negative bacteria in the gut and intestine [38,39]. The data suggested the presence of quorum sensing-promoting molecules (oligopeptides) produced by L. helveticus ATCC 15009 are responsible for promoting bacterial communication and therefore enhance the antagonism of Lactobacillus sp. against Gram-negative bacteria. Moreover, several preceding studies has found that the inhibitory properties of oral lactobacilli can be attributed to the secretion of various antimicrobial substances or metabolites such as organic acids, hydrogen peroxide, diacetyl, bacteriocins, peptides and other bioactive molecules [40,41]. Specifically, organic acids generated during glucose fermentation, comprising lactic and acetic acid, hinder the growth of strains that are less acid-tolerant [42]. As a result, this may demonstrate the inhibitory activity between L. helveticus and P. gingivalis.
Once the antibacterial efficiency of the L. helveticus strain had been determined, the next stage was to assess its impact on biofilm biomass activity. In natural environments, bacteria primarily exist in structured communities attached to surfaces and surrounded by a self-produced extracellular matrix. When bacterial cells develop biofilm, they produce extracellular polymeric substance (EPS), which protects the biofilm and makes it tough to break down. This provides bacterial cells in biofilm mode with a significant ability to withstand environmental challenges, principally through the protective EPS [43]. These findings suggest that biofilm formation serves as a protective mechanism, redirecting metabolites towards EPS synthesis to provide enhanced protection against stressful conditions by manipulating metabolic processes. The biofilm network creates an anaerobic environment for bacteria embedded within it, allowing them to thrive and secure more resources compared to planktonic cells [44]. Therefore, investigating L. helveticus capacity to interrupt biofilm development at various doses is crucial in preventing P. gingivalis recolonisation [23].
In this study, the biofilm biomass of P. gingivalis exhibited a rapid decrease in the treated samples when compared to the control groups, indicating susceptibility to L. helveticus. This noteworthy finding suggests that L. helveticus may offer a promising approach for mitigating a significant proportion of P. gingivalis-related pathogenicity. These results align closely with various earlier findings which discovered that lactobacilli can disrupt pathogenic bacteria and fungus by fighting for nutrients, co-aggregation, and the generation of antimicrobials such as bacteriocin and hydrogen peroxide, organic acids and influencing the immune system [45]. Moreover, there are few studies showing that lactobacilli can integrate into target biofilms and compete with pathogens for attachment sites in the oral cavity. For example, Vestman et al. (2013) revealed that L. reuteri PTA5289 may be absorbed into the oral microbiota for a brief length of time during a 6-week intervention, resulting in a delay in the regrowth of S. mutans after oral disinfection with chlorhexidine [46]. Furthermore, James et al. (2016) discovered that a combination of three probiotic strains (L. helveticus CBS N116411, L. plantarum SD57870, and S. salivarius DSM14685) was efficient in both preventing and eliminating C. albicans biofilms [47].
The impact of L. helveticus were further examined by assessing its action against P. gingivalis using a LIVE/DEAD cells viability assay with fluorescence microscopic analysis. The LIVE/DEAD® BacLight^TM^ assay was used to differentiate live and dead cells based on membrane integrity, utilising a dual staining procedure of SYTO 9 and propidium iodide (PI) [48]. Both dyes intercalate with nucleic acids, resulting in an enhanced fluorescent signal. This kit employs SYTO 9 and PI dyes, which have different membrane permeability properties. SYTO 9 can enter all bacterial cell membranes, while PI can only enter cells with disrupted membranes. This allows for distinction between live and dead cells based on the relative green and red fluorescence from SYTO 9 and PI staining [49]. Detection of cell viability using the BacLight Kit offers numerous advantages over colony counting on agar plates, as it provides a rapid and direct quantification of cell death in real time. According to the images obtained, there was a higher level of bacterial cells and a dense biofilm structure of untreated P. gingivalis compared to the P. gingivalis biofilm treated with 10^9^ cells/mL L. helveticus. The exposure of L. helveticus to P. gingivalis biofilm revealed a clear sparsity and dispersion of the biofilm, indicating a significant reduction in viable cells. These findings support the biofilm assay results, where P. gingivalis biofilm formation was substantially inhibited by 10^9^ cells/mL of L. helveticus.
In this study, FESEM was utilised to further verify the ultra-morphological effects of L. helveticus against P. gingivalis. FESEM is an advanced technique used to capture high-resolution images of the microstructure of materials. The FESEM images of the current study revealed morphological changes in P. gingivalis cells after treatment with L. helveticus, compared to those without treatment, where cells were observed to have regular spherical shapes with smooth surfaces and intact cell membrane. However, not all bacterial cells were affected with L. helveticus. Some cells appeared distressed, revealing cell membranes, with blisters forming on the surface, as indicated by the yellow arrows in Fig 4C. Other P. gingivalis cells, marked by the white dotted arrow, illustrate that this cell was not affected by L. helveticus. In Fig 4C, the red arrow indicates a visible distortion of the cell wall in P. gingivalis. This observation suggests a disruption in the bacterium’s structural integrity. Such damage may involve disruption of the cytoplasmic membrane, leading to coagulation of the cell contents and impairing the membrane’s ability to act as a permeability barrier. The binding and antibacterial effect of L. helveticus on P. gingivalis were evident, warranting further investigation into the specific damage caused by L. helveticus-derived peptides. Genetic analysis reported that P. gingivalis primarily relies on amino acids for growth, with limited uptake and metabolism of carbohydrates [50]. A previous work on model membranes revealed that bacteriocins could interact with lipid membranes without a specified target antigenic determinant. The contact was most likely caused by an initial electrostatic attraction between the cationic bacteriocins and the anionic lipid membrane, which is a common mechanism found in many antimicrobial peptides [51]. P. gingivalis cells have been found to be highly negatively charged at pH values greater than 3 [52]. Matsuzaki et al. (1999) discovered that these positively charged antimicrobial peptides are drawn to the negatively charged components of P. gingivalis cells, which gravitate the initial encounter prior to pore development [53]. Similar mechanisms have been found in various cationic antimicrobial peptides, both natural and synthetic [54,55].
In summary, this study highlights important insights into the antibacterial effects of L. helveticus against P. gingivalis. However, to enhance understanding and address the limitations of these findings, several approaches should be considered. Further studies should focus on elucidating the mechanisms of action of L. helveticus and employing models that replicate the oral environment, such as co-culture systems with other oral bacteria. Understanding the interactions among various microbial species, host immune responses, and environmental factors is crucial for evaluating the therapeutic potential of L. helveticus. Furthermore, the use of advanced in vitro techniques, such as whole genome sequencing and proteomics can provide more in-depth information about how L. helveticus influences cell viability and metabolic activity in harmful biofilms. In addition, ex vivo and in vivo studies should be conducted to validate and strengthen these findings. Addressing these aspects will enhance the development of effective probiotic treatments for improving oral health.
Conclusion
Succinctly, this study demonstrates that L. helveticus ATCC 15009 displayed profound inhibitory effects on the oral pathogen P. gingivalis, leading to reduced cell growth and morphological changes in bacteria. Thus, these findings suggest that probiotic therapy could be beneficial in managing periodontal diseases. Moreover, the potent antibacterial effect of L. helveticus against P. gingivalis emphasizes the importance of further research to identify optimal probiotic strategies that could enhance periodontal health. Understanding how probiotic bacteria can inhibit the growth of pathogenic strains may be crucial in developing preventive measures for periodontitis and optimizing treatment outcomes.
Supporting information
S1 FileThe inhibition zone diameters of L. helveticus against P. gingivalis.(XLSX)
S2 FileEffect of L. helveticus against P. gingivalis biofilm.(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Xu W, Zhou W, Wang H, Liang S. Roles of Porphyromonas gingivalis and its virulence factors in periodontitis. Adv Protein Chem Struct Biol. 2020;120:45–84. doi: 10.1016/bs.apcsb.2019.12.001 32085888 PMC 8204362 · doi ↗ · pubmed ↗
- 2Könönen E, Gursoy M, Gursoy UK. Periodontitis: A Multifaceted Disease of Tooth-Supporting Tissues. J Clin Med. 2019;8(8):1135. doi: 10.3390/jcm 8081135 31370168 PMC 6723779 · doi ↗ · pubmed ↗
- 3Kinane DF, Stathopoulou PG, Papapanou PN. Periodontal diseases. Nat Rev Dis Primers. 2017;3:17038. doi: 10.1038/nrdp.2017.38 28805207 · doi ↗ · pubmed ↗
- 4Li X, Liang H, Huang Y, Hu Q, Liang L, He J, et al. Near-infrared light-responsive copper-cerium bimetallic oxide nanozyme with antibacterial and antioxidant abilities for periodontitis therapy. Colloids Surf B Biointerfaces. 2025;252:114685. doi: 10.1016/j.colsurfb.2025.114685 40233479 · doi ↗ · pubmed ↗
- 5Isola G, Santonocito S, Lupi SM, Polizzi A, Sclafani R, Patini R, et al. Periodontal Health and Disease in the Context of Systemic Diseases. Mediators Inflamm. 2023;2023:9720947. doi: 10.1155/2023/9720947 37214190 PMC 10199803 · doi ↗ · pubmed ↗
- 6AleksijevićLH, AleksijevićM, Škrlec I, Šram M, Šram M, Talapko J. Porphyromonas gingivalis Virulence Factors and Clinical Significance in Periodontal Disease and Coronary Artery Diseases. Pathogens. 2022;11(10):1173. doi: 10.3390/pathogens 11101173 36297228 PMC 9609396 · doi ↗ · pubmed ↗
- 7Murugaiyan V, Utreja S, Hovey KM, Sun Y, La Monte MJ, Wactawski-Wende J, et al. Defining Porphyromonas gingivalis strains associated with periodontal disease. Sci Rep. 2024;14(1):6222. doi: 10.1038/s 41598-024-56849-x 38485747 PMC 10940620 · doi ↗ · pubmed ↗
- 8Zhang Z, Liu D, Liu S, Zhang S, Pan Y. The Role of Porphyromonas gingivalis Outer Membrane Vesicles in Periodontal Disease and Related Systemic Diseases. Front Cell Infect Microbiol. 2021;10:585917. doi: 10.3389/fcimb.2020.585917 33585266 PMC 7877337 · doi ↗ · pubmed ↗