The regulatory effects of PD-1/PD-L1 inhibitors on bone metabolism: opportunities and challenges in osteoporosis management
Jia-Wen Wang, Mu-Wei Dai, Jia-Hui Liu

TL;DR
PD-1/PD-L1 inhibitors, used in cancer treatment, have mixed effects on bone health, offering both protection and causing fractures, which requires careful management.
Contribution
This paper provides a comprehensive review of the dual regulatory effects of PD-1/PD-L1 inhibitors on bone metabolism and proposes strategies for managing bone health during immunotherapy.
Findings
PD-1/PD-L1 inhibitors show bone-protective benefits but also cause bone-related adverse events in up to 69% of patients.
The contradictory effects may stem from clinical variables, microenvironmental factors, and interactions with pathways like Wnt/β-Catenin.
Osteoporosis and fragility fractures during therapy are suggested as a subclass of immune-related adverse events.
Abstract
Programmed death-1 (PD-1) and its ligand PD-L1 inhibitors have become pivotal agents in cancer immunotherapy, demonstrating significant efficacy across multiple malignancies. However, beyond regulating T cell activation, the PD-1/PD-L1 axis also exerts complex and critical effects on bone metabolism. Notably, both clinical observations and mechanistic studies have revealed a paradox: on one hand, PD-1/PD-L1 blockade appears to confer bone-protective benefits; on the other hand, it has been associated with bone-related adverse events (AEs) in up to 69% of patients, including pathological fractures and vertebral compression fractures. This review comprehensively explores the bidirectional regulatory effects of the PD-1/PD-L1 pathway on bone metabolism and investigates the underlying mechanisms contributing to these contradictory findings. The discrepancies may be attributed to a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
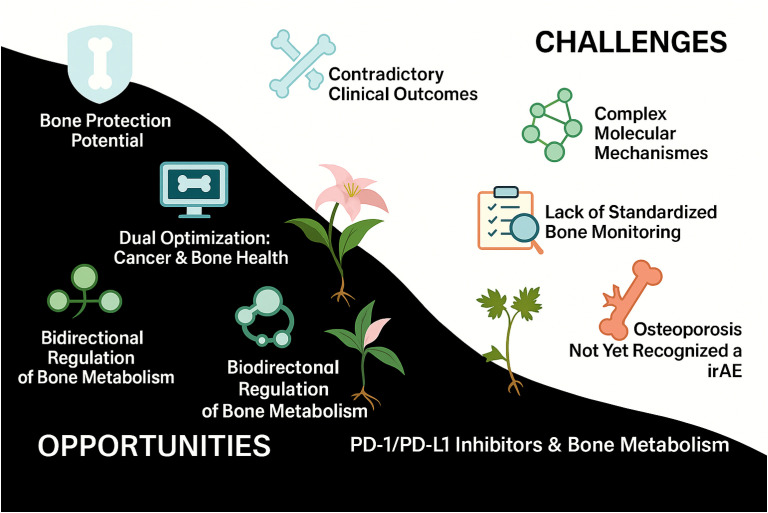 Figure 1
Figure 1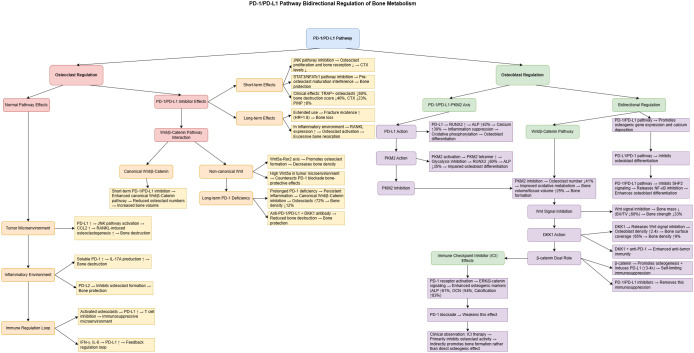 Figure 2
Figure 2| Year | Model / System | Intervention & Main Indicators | Osteoclast / Bone Metabolism Findings | Mechanistic Insights | Ref. |
|---|---|---|---|---|---|
| 2020 | Lewis lung carcinoma bone metastasis mouse model; RAW264.7 cells | Anti-PD-1 (nivolumab 10 mg/kg) or Pdcd1–/–; TRAP staining, μCT, serum CTX-I/PINP | TRAP+ osteoclasts ↓ ≈60%; serum CTX-I ↓ (Day 8); bone destruction score ↓ 40%; pain alleviation | Tumor-derived PD-L1 activates PD-1 on osteoclast precursors → JNK → CCL2 ↑ → promotes RANKL-driven osteoclastogenesis; PD-1 blockade reverses this process | ( |
| 2022 | PD-1–/– and PD-L1–/– mice; human/mouse BM cells | Gene knockout or neutralizing antibodies; RANKL/M-CSF induction | PD-1 KO: RANKL ↑, RANKL/OPG ratio ↑; TRAP+ osteoclasts ↑ 3–4-fold; BMD ↓ | PD-1 pathway suppresses RANKL and osteoclast activation in inflammatory settings; its loss leads to excessive bone resorption | ( |
| 2023 | Cohort of 2,532 cancer patients receiving ICIs | Observational cohort; endpoint: major fracture incidence | Fracture incidence 7.3 vs 3.6/100 person-years; adjusted HR ≈ 1.6 (95% CI: 1.2–2.2) | Enhanced T cell activation + corticosteroids may accelerate bone loss; highlights need for skeletal monitoring | ( |
| 2024 | 9 solid tumor patients on ICI monotherapy (no bone metastases) + ex vivo 3D bone model | PD-1/PD-L1 inhibitors (pembrolizumab, atezolizumab); dynamic bone turnover markers (CTX, PINP, OCN); PBMC–osteoblast/osteoclast coculture | CTX ↓ 23% at 1 month (p = 0.01); PINP ↑ 8% at 4 months (p = 0.02); ex vivo TRAP+ osteoclasts ↓ 50–70% dose-dependently | ICIs inhibit mature osteoclast differentiation, disrupting OC/OB coupling; net effect favors osteogenesis | ( |
| Year | Model / Population | Wnt/β-Catenin Regulation (Canonical vs Non-Canonical) + ICI | Osteoclast / Bone Findings | Mechanistic Insights | Ref. |
|---|---|---|---|---|---|
| 2012 | Wnt5a–/–, Ror2–/–, and WT mice | Non-canonical: Wnt5a–Ror2 axis | TRAP+ osteoclasts ↓ 45–60%; BMD ↑ 20–33% | Osteoblast-derived Wnt5a → Ror2+ osteoclast precursors → NFATc1 upregulation; tumor-bone microenvironmental Wnt5a may negate PD-1 blockade bone-protective effects | ( |
| 2020 | Lewis lung carcinoma femoral metastasis model; nivolumab i.v. | ICI monotherapy: anti-PD-1 | TRAP+ osteoclasts ↓ ≈60%; μCT BV/TV ↑ 29%; CTX-I ↓; pain relief | PD-1–SHP2 blockade → suppresses JNK–CCL2 cascade → inhibits osteoclast precursor differentiation; canonical Wnt activation may synergize | ( |
| 2021 | Lung cancer (CMT167/MC38) + bone marrow fibroblasts (BMF) | Canonical inhibition: Tankyrase inhibitor XAV-939 + anti-PD-L1 | TRAP activity ↓ ≈45% in co-culture; CD8+IFN-γ+ T cells ↑ 4.2-fold in combo group | BMF-induced β-catenin → PD-L1 ↑; XAV-939 inhibits β-catenin → PD-L1 ↓ → enhances anti-PD-L1 efficacy and suppresses osteoclasts | ( |
| 2022 | Gastric cancer xenograft + dual antibody therapy | Canonical antagonism: High DKK1 (β-catenin inhibitor) + anti-PD-1 | DKK1 overexpression → OC area ↑ 2.1-fold, OPG ↓; DKK1-mAb + anti-PD-1 → OC area ↓ 48%, bone metastases ↓ 40% | DKK1 suppresses β-catenin → M2-like TAM ↑, RANKL ↑; DKK1 neutralization restores β-catenin–OPG axis, suppresses osteoclasts and enhances ICI efficacy | ( |
| 2025 | Pdcd1–/– mice under physiological conditions | PD-1 deletion → low canonical Wnt activity | Sex-specific: Male mice TRAP+ osteoclasts ↑ 72%, BMD ↓ 12%; β-catenin target Axin2 ↓ | Chronic PD-1 loss → inflammation chemotaxis → inhibits β-catenin–OPG signaling → bone resorption imbalance; suggests prolonged/high-dose ICI or inflammatory context may reverse bone protection | ( |
| Year | Model / Intervention | Osteogenic Outcomes (ALP / Mineralization, etc.) | Mechanistic Insights | Ref. |
|---|---|---|---|---|
| 2020 | MC3T3-E1 + PKM2 activator DASA-58 | RUNX2 mRNA ↓ 60%; ALP ↓ 35%; impaired osteogenesis | PKM2 tetramer formation ↑ → glycolysis ↓ → suppressed osteoblast activity | ( |
| 2022 | hADMSC–osteoblast coculture ± PD-L1 | PD-L1 treatment → RUNX2 ↑; ALP activity ↑ 42%; mineralized nodules ↑ 30% | PD-1/PD-L1 alleviates IL-6/TNF-α inflammation; suggests metabolic shift toward OxPhos | ( |
| 2024 | Clinical ICI-treated cohort (n = 428) | Vertebral BMD ↑ 5.6% at 12 months; ALP/mineralization unchanged | ICIs mainly inhibit osteoclasts; limited direct osteoblast impact observed clinically | ( |
| 2025 | BMSCs: si-PKM2 or TEPP46 | PKM2-KD: BV/TV ↑ 25%, OC.N/BS ↓ 41%; TEPP46 showed similar effects | PKM2 inhibition promotes oxidative metabolism and mitochondrial restoration | ( |
| Year | Tumor Model / System | Wnt/β-Catenin Regulation + ICI Treatment | Osteoblast/Bone Metabolism Outcomes | Mechanistic Insights | Ref. |
|---|---|---|---|---|---|
| 2018 | C57BL/6 mice (♀, 10 w) | Porcupine inhibitor LGK974 (5 mg/kg/day, 4 weeks) ± anti-PD-1 combo (RxC004/NIVO, public data) | Trabecular BV/TV ↓ ≈60%; cortical thickness ↓ 22%; MAR/BFR ↓ 40–45%; 3-pt bending strength ↓ 33% | Global Wnt inhibition → β-catenin suppression → Runx2/OPG ↓, Sost ↑; osteoblast inhibition with secondary resorption; suggests need for bone protection in ICI combination strategies | ( |
| 2020 | U251 and GL261 glioma + ICG-001/anti-PD-1 | Wnt3a/EGF → β-catenin ↑ → PD-L1 ↑ 3–4×; β-catenin/AKT inhibition or anti-PD-1 reverses effects | β-catenin–OE BMSC coculture: ALP/OCN ↑ 1.7×; PD-L1 suppresses CD8+ T activation; ICG-001 + anti-PD-1 restores CD8 function and maintains Runx2/Col1a1 → osteogenesis preserved | Canonical β-catenin promotes osteogenesis but induces PD-L1 → “self-limiting” immunosuppression; PD-1 blockade releases immune brake without impairing bone formation | ( |
| 2007 + 2021 | SCID-rab myeloma model + B16F0/4T1 | DKK1-neutralizing mAb (BHQ880/DKN-01) ± anti-PD-1 | Myeloma transplant BMD ↑ 9% (vs −6%); OCN+ osteoblast density ↑ 2.4×; bone surface osteoblast coverage ↑ 65%; DKN-01 + anti-PD-1 maintained bone mass and inhibited tumors | DKK1 blockade → LRP5/6 reactivation → β-catenin–TCF–Runx2–OPG restored; osteoblast proliferation and anti-resorptive effects; also reduces MDSCs and activates NK/T cells → enhanced PD-1 efficacy | ( |
| 2022 | hADMSC-derived osteoblasts ± BMS-202 | Exogenous PD-L1 / exosomes → PD-1 activation; BMS-202 inhibition | ALP ↑ 61%, OCN ↑ 54%, Ca²+ crystal area ↑ 83%; BMS-202 normalized osteogenesis index to control | PD-1 on osteoblasts → p-ERK/β-catenin ↑ → COL1A1–RUNX2 signaling ↑; low-inflammation microenvironment; co-culture with osteoclasts maintains bone homeostasis | ( |
| Year | Tumor Type / Cohort (n) | Key Clinical Features Assessed | Bone Metabolic Effects & Core Findings | Mechanistic Implications | Ref. |
|---|---|---|---|---|---|
| 2020 | Breast cancer bone metastasis (mouse model) | Tumor burden (bone metastasis vs. non-metastatic) | PD-1 blockade ↓ osteoclasts by 46%, suppressed bone resorption | PD-1 expressed on osteoclast precursors; blockade → NFATc1 inhibition → anti-resorptive effect | ( |
| 2023 | Various solid tumors (claims database, n = 1,873) | Age (≥65 vs <65), sex, mono- vs. dual-ICI | Major fracture risk ↑ 2.5-fold within 1 year post-ICI; highest risk in elderly females and dual-ICI users | T cell activation → RANKL upregulation → increased bone resorption | ( |
| 2024 | Melanoma (claims database, n = 3,137) | Age (mean 68), sex (36% female), dual ICI, prior fractures | MOF risk HR ≈ 1.8 during years 1–2 post-ICI; higher risk in older age, females, and combination therapy | Systemic immune activation → hyperactive osteoclastogenesis | ( |
| 2024 | Multiple solid tumors (prospective cohort, n = 57) | Mean age 59 ± 13; 57% prior chemotherapy; 21% prior radiotherapy | P1NP ↑ 34%, CTX ↓ 18% at 12 weeks → bone formation favored | PD-1/PD-L1 blockade ↑ CD14+ osteoprogenitors, promoting osteogenesis | ( |
| 2024 | NSCLC (n = 229) | Baseline BMD (QCT) and TNM stage III–IV | Low BMD group: PFS HR 1.72, OS HR 1.88 → bone loss correlated with poor ICI outcomes | Bone mass reflects systemic inflammation/nutrition; bone-immune interplay | ( |
| 2025 | Mouse model + patient serum | Age, sex | Young female mice: trabecular bone volume ↓ 33%; males unaffected; aged mice of both sexes showed BMD decline | CD3+ T cell–mediated RANKL/OPG imbalance; PD-1 blockade drives T cell–dependent bone loss | ( |
| 2025 | Melanoma (opportunistic QCT, n = 98) | Baseline vBMD, high-dose corticosteroids | vBMD ↓ 6.9 mg/cm³ at 12 months in non-ICI group; stable in ICI group; patients with low baseline vBMD still declined | ICI may preserve bone by suppressing inflammation-driven resorption; corticosteroids counteract benefit | ( |
| Year | Study Design / Cohort (n) | Baseline BMD Evaluation Rate | Baseline Use of Bone-Modifying Agents | Bone Events or Biomarker Data | Identified Monitoring / Intervention Gaps | Ref. |
|---|---|---|---|---|---|---|
| 2018 | Case series of 6 “bone irAEs” | 0% (no DXA performed before fractures) | 0% (ZA or denosumab started post-fracture) | 3 new vertebral fractures; 3 focal osteolytic lesions | No baseline BMD assessment or preventive treatment; multiple early skeletal events | ( |
| 2021 | Case series (n = 4) + FAERS pharmacovigilance (n = 650) | 0% (no DXA in cases; FAERS lacked data) | <10% reported concurrent use of BMA | FAERS: pathological fracture ROR 3.17; 3/4 case series with multiple vertebral fractures | Large-scale reports lack bone baselines; bone-protective treatment highly underused | ( |
| 2022 | Prospective observational; NSCLC/RCC (n = 44) | 0% (DXA/QCT not performed; only CTX-I, P1NP tested) | 0% (excluded prior BMA users) | CTX-I ↑, P1NP ↓ within 3 months → increased bone resorption | No imaging-based BMD at baseline; no protective intervention | ( |
| 2023 | Real-world index cohort (Canada; n = 1,600 ICI users) | NR (DXA not recorded in database) | 8.8% received anti-osteoporotic drugs before ICI | MOF incidence 27.3/1000 person-years post-ICI; IRR 2.43 | >90% without bone protection; no risk stratification applied | ( |
| 2024 | Prospective cohort; ICI monotherapy in various cancers (n = 9) | 0% (osteoporotic patients excluded; no DXA) | 0% (excluded prior ZA/denosumab use) | CTX ↓ at 1 month; PINP & OCN ↑ at 4–6 months → biphasic metabolic shift | Lack of baseline bone mass assessment limits clinical interpretation of metabolic changes | ( |
| Year | Key Pathway | Drug / Modality | Core Bone Metabolic Effects & Representative Data | Overlaps/ Key Differences | Ref. |
|---|---|---|---|---|---|
| 2014 | IPP–γδ T cell activation axis | Zoledronic acid → γδ T cell | ZA inhibits FPP synthase → IPP accumulation → activates Vγ9Vδ2 T cell degranulation and cytotoxicity against monocyte–myeloid lineage | Unique “immune adjuvant” effect of bisphosphonates; may synergize with ICIs to boost antitumor immunity; primarily anti-resorptive | ( |
| 2020 | RANKL → RANK → NFATc1 (osteoclastogenesis axis) | PD-1 inhibitor (nivolumab) | In murine bone metastasis: anti-PD-1 ↓ osteoclasts by 46%, inhibited bone resorption, relieved pain; blocks PD-1–SHP2 → suppresses NFATc1 | Targets same axis as denosumab but upstream (PD-1 vs RANKL); adds immuno-oncologic benefit | ( |
| 2022 | Mevalonate–FPP synthase | Nitrogen-containing bisphosphonates (ZA) | FPP synthase inhibition → IPP/ApppI accumulation → osteoclast apoptosis; CTX ↓ 50–60%, annual BMD ↑ 4–6% | Downstream metabolic action; RANKL-independent; complements both ICI and denosumab | ( |
| 2023 | RANKL → RANK/TRAF6 | Denosumab (anti-RANKL mAb) | Phase III trials: delayed skeletal events by 18–23%; CTX ↓ up to 80% | Same ligand target as ICI (RANKL) but acts via direct neutralization; fast, reversible effect; limited immune activation | ( |
| 2024 | T cell activation–derived RANKL | PD-1/PD-L1 antibodies (prospective multicancer cohort) | P1NP ↑ 34% at 12 weeks; 1-year fracture risk ↑ 2.5× (elderly/female at highest risk) | ICI shares ligand (RANKL) with denosumab but shows bidirectional effects; bone protection strategies required | ( |
| 2024 | RANKL blockade + PD-1 blockade combination | Denosumab + ICI | NSCLC with bone metastasis: ORR ↑ from 37% → 54%; skeletal events ↓ 30%; no increase in irAEs | Highlights RANKL–bone and checkpoint–immune synergy; dual benefit in bone protection and | ( |
| Year | Design / Population | Key Bone Findings | Implications for Monitoring–Stratification–Prevention | Ref. |
|---|---|---|---|---|
| 2023 | Review | Summarized bidirectional bone effects of ICIs; proposed follow-up algorithm | Recommends “Bone-Oncology Joint Clinic”: baseline DXA + BTM → 6-month reassessment → annual review | ( |
| 2023 | Review | ICIs may worsen outcomes in bone metastatic cancer; benefit from bone-targeted co-treatment | Advocates for separate stratification of bone metastasis patients; ICI + anti-resorptive combo and enhanced bone pain surveillance | ( |
| 2023 | Provincial Canadian database; 2,285 ICI-naïve cancer patients | 1-year fracture HR 2.6 post-ICI; most involved spine/ribs | Suggests DXA + FRAX at ICI initiation; anti-resorptives for high-risk individuals | ( |
| 2024 | Retrospective–matched cohort; 479 NSCLC patients on ICIs | Low CT-BMD group had shorter OS (~3 months); no impact on PFS | Proposes CT-derived BMD as prescreening tool; integrate low-BMD patients into osteoporosis prevention pathway | ( |
| 2024 | Prospective cohort (n = 30) + 3D bone model validation | After 3 months: CTX ↓ 25%, OCN ↑ 18% | Emphasizes dual-track: serum bone turnover markers + imaging; timely Ca/Vit D supplementation | ( |
| 2024 | Multicenter melanoma retrospective cohort (n = 1,104) | MOF incidence 31.2/1000 person-years (vs. 14.8); shoulder–hip–spine predominant | Proposes “Mel-ICI Fracture Score”; patients scoring ≥4 should initiate bisphosphonates/denosumab | ( |
| 2025 | ML-aided QCT study; 132 ICI patients | Median L1 vBMD ↓ 5.4% at 6 months; AI enabled automated quantification | Embeds AI-QCT into routine imaging to enable longitudinal monitoring and personalized alerts | ( |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBone health and treatments · Cancer Immunotherapy and Biomarkers · Cancer Diagnosis and Treatment
Introduction
1
Inhibitors targeting programmed death-1 (PD-1) and its ligand PD-L1 have profoundly reshaped cancer therapy. Since the first PD-1 inhibitor received regulatory approval in 2014, these agents have significantly improved overall survival (OS) and progression-free survival (PFS) rates (1), becoming standard treatments for a wide range of malignancies, including non-small cell lung cancer (NSCLC), melanoma, head and neck squamous cell carcinoma, and renal cell carcinoma (2).
Beyond their immunomodulatory effects on T cells, the PD-1/PD-L1 axis plays a multifaceted role in bone metabolism. Murine models with PD-1 or PD-L1 gene knockout exhibit pronounced osteoporotic phenotypes, including decreased trabecular bone volume, disrupted microarchitecture, elevated osteoclastogenesis, and increased RANKL/OPG ratios (3). These outcomes are mediated via multiple signaling cascades that finely regulate bone-resorbing and bone-forming cells (4–6). However, the complexity of these mechanisms has led to conflicting results. While some clinical studies suggest that PD-1/PD-L1 inhibitors exert bone-protective effects (4), others report the opposite, documenting bone-related adverse events in patients receiving immune checkpoint inhibitors (ICIs) (7, 8), even as some individuals maintain stable bone mineral density (9).
At the molecular level, similarly inconsistent findings are observed. Some investigations describe a pro-osteogenic role for PD-1/PD-L1 blockade (10), while others note inhibitory effects on osteoblast differentiation (6). These contradictions are likely influenced by a convergence of factors, including patient-specific clinical features, the immune microenvironment, cell-type-specific responses, soluble PD-1/PD-L2 activity (11), and the bidirectional nature of the Wnt/β-Catenin pathway (5, 12) and PD-L1–PKM2 metabolic signaling (13).
Furthermore, clinical evidence links PD-1/PD-L1 inhibitors to a heightened risk of pathological fractures, vertebral compression fractures, and femoral neck fractures (14), contributing to a cumulative incidence of bone-related adverse events as high as 69% (15). Despite their prevalence and clinical significance, conditions such as osteoporosis and fragility fractures are not formally recognized as immune-related adverse events (irAEs) (16), underscoring a gap in clinical classification and management.
Figure 1 outlines the opportunities and challenges associated with PD-1/PD-L1 blockade in bone metabolism, emphasizing its potential protective effects alongside its regulatory complexity and the under-recognition of skeletal irAEs such as osteoporosis.
Opportunities and challenges of PD-1/PD-L1 inhibitors in bone metabolism. This schematic illustrates the dual impact of PD-1/PD-L1 inhibitors on bone metabolism. The left panel highlights the potential benefits, including bone-protective effects, bidirectional immuno-skeletal regulation, and dual optimization of tumor control and bone health. In contrast, the right panel outlines key challenges, such as inconsistent clinical outcomes, complex molecular mechanisms, the absence of standardized bone surveillance protocols, and the underrecognition of osteoporosis as an immune-related adverse event (irAE).
To elucidate the dual role of PD-1/PD-L1 inhibitors in bone metabolism and explore their implications in osteoporosis prevention, this review addresses the following key questions:
What accounts for the contradictory findings regarding the effects of PD-1/PD-L1 inhibitors on bone metabolism? Why do some studies report bone-protective outcomes (e.g., reduced resorption markers and preserved BMD), while others observe increased bone-related AEs such as vertebral fractures? What clinical factors (e.g., baseline patient characteristics, cancer type, treatment regimens) may underlie these discrepancies?How does the PD-1/PD-L1 pathway achieve bidirectional regulation of bone metabolism? Through which signaling networks does it affect osteoclast differentiation/function and osteoblast activity/mineralization? What roles do the Wnt/β-Catenin pathway and the PD-L1–PKM2 axis play in this context, and how do these mechanisms reconcile clinical inconsistencies?How can the dual regulatory features of PD-1/PD-L1 blockade be translated into osteoporosis prevention strategies? What are the evidence-based approaches for integrating bone health monitoring and stratified prevention during ICI therapy? What justifies the inclusion of osteoporosis and fragility fractures as irAEs, and how can this reclassification enhance both cancer and skeletal outcomes?
Effects of PD-1/PD-L1 inhibitors on bone metabolism
2
PD-1/PD-L1 immune checkpoint inhibitors have become a cornerstone of modern cancer immunotherapy. However, their effects on bone metabolism remain underrecognized and exhibit contradictory findings across studies. Gassner et al. reported that in cancer patients without bone metastases, treatment with PD-1/PD-L1 inhibitors led to a significant early reduction in the bone resorption marker CTX (from a baseline mean of 0.51 ng/ml to 0.42 ng/ml at week 3), while bone formation markers such as PINP and osteocalcin (OCN) increased after 4 months of treatment, suggesting a bone-protective effect (4). Conversely, Pantano et al. observed a marked increase in CTX-I levels and a downward trend in PINP levels after 3 months of immune checkpoint inhibitor (ICI) therapy, which correlated with poor treatment response and decreased survival (7).
Moreover, some studies have documented adverse skeletal events during ICI treatment, including vertebral compression fractures and osteolytic lesions (8). In contrast, others have reported relatively stable bone mineral density (BMD) in ICI-treated patients compared to non-ICI counterparts, suggesting a long-term bone-preserving effect (9). These conflicting outcomes underscore the complex and multifactorial nature of PD-1/PD-L1 blockade on bone metabolism, likely influenced by clinical heterogeneity such as age, sex, tumor type, treatment regimen (monotherapy vs. combination therapy), and baseline skeletal health (17).
Importantly, there remains a lack of standardized bone health monitoring protocols for patients receiving ICIs, potentially delaying the detection and intervention of subclinical bone metabolic disorders (18). This gap may contribute to skeletal-related adverse events (SREs), ultimately impairing therapeutic efficacy and quality of life. Therefore, the impact of PD-1/PD-L1 inhibition on bone health warrants greater clinical attention and systematic evaluation.
Regulatory effects of PD-1/PD-L1 pathway and inhibitors on osteoclasts
3
While the PD-1/PD-L1 axis plays a pivotal role in immune regulation, it also significantly influences bone metabolism, particularly osteoclast differentiation and function. Genetic deletion of PD-1 or PD-L1 leads to osteoporotic phenotypes, implicating this pathway in the maintenance of bone homeostasis (3). Within the tumor microenvironment (TME), upregulation of PD-L1 and CCL2 facilitates osteoclastogenesis by activating the JNK pathway and enhancing CCL2-mediated RANKL signaling, thereby promoting bone resorption (5). Additionally, soluble PD-1 (sPD-1), which is elevated in inflammatory settings, stimulates IL-17A production—a key mediator of osteoclast activation—resulting in accelerated bone destruction. In contrast, PD-L2 expression under inflammatory conditions appears to suppress osteoclastogenesis and confer bone-protective effects (11).
Osteoclasts, particularly in their activated state, can upregulate PD-L1 expression to inhibit T cell proliferation and cytotoxicity, contributing to an immunosuppressive microenvironment. This PD-L1 upregulation is itself modulated by pro-inflammatory cytokines such as IFN-γ and IL-6, forming a feedback regulatory loop (19, 20).
PD-1/PD-L1 inhibitors interrupt this axis and exert both direct and indirect effects on osteoclast activity. By inhibiting the JNK pathway, these agents reduce osteoclast proliferation and resorptive capacity, leading to decreased CTX levels (4, 5). They can also interfere with the STAT3/NFATc1 signaling cascade, impeding pre-osteoclast maturation and reversing osteoclast-mediated immunosuppression (20). This dual mechanism results in a bidirectional modulation of bone remodeling (4), with short-term treatment reducing TRAP+ osteoclasts by approximately 60%, lowering bone destruction scores by 40%, and decreasing CTX by 23%, while increasing PINP levels by 8% at 4 months (p = 0.02) (4, 5). For a detailed summary of these findings across preclinical and clinical contexts, see Table 1.
However, prolonged ICI therapy has been associated with increased fracture risk, with adjusted hazard ratios nearing 1.6 (21), suggesting a shift toward net bone loss over time. Chronic inflammation during extended PD-1 blockade can enhance RANKL expression and osteoclast activation, further contributing to skeletal damage (3).
Wnt/β-Catenin signaling also plays a complex, bidirectional role in mediating the skeletal effects of PD-1/PD-L1 inhibition (Table 2). The canonical Wnt/β-Catenin pathway synergizes with short-term PD-1 blockade to suppress osteoclastogenesis and increase bone mass (5). In contrast, the non-canonical Wnt5a–Ror2 axis promotes osteoclast formation and reduces BMD, with Wnt5a overexpression in the TME antagonizing the bone-preserving effects of PD-1 inhibitors (12). Moreover, chronic PD-1 deficiency can suppress canonical Wnt signaling through sustained inflammation, leading to a 72% increase in osteoclast numbers and a 12% decrease in bone density (22).
To mitigate this deleterious effect, concurrent administration of anti-DKK1 antibodies—targeting inhibitors of canonical Wnt signaling—has shown promise in reducing skeletal damage and restoring bone homeostasis in the context of long-term ICI therapy (23).
Regulation of osteoblasts by the PD-1/PD-L1 pathway
4
Although the PD-1/PD-L1 pathway has been shown to promote osteogenic gene expression and calcium deposition (10), it can also inhibit osteogenic differentiation (6). This inhibition may occur through suppression of the SHP2 signaling pathway, which relieves its inhibitory effect on NF-κB activation, thereby enhancing osteoblast differentiation and bone formation (24). These seemingly contradictory effects may be related to the PD-1/PD-L1–PKM2 axis (13) (see Table 3). The balance within this axis plays a critical role: PD-L1 promotes osteogenic differentiation by upregulating RUNX2 expression, increasing ALP activity by 42%, enhancing mineralization by 30%, reducing inflammation, and promoting oxidative phosphorylation metabolism (10). In contrast, activation of PKM2 leads to increased tetramer formation, inhibition of glycolysis, a 60% reduction in RUNX2 expression, and a 35% decrease in ALP activity, collectively impairing osteoblast differentiation (25). Notably, PKM2 inhibition can reduce osteoclast numbers by 41%, improve oxidative metabolism and mitochondrial function, and enhance bone volume/tissue volume (BV/TV) ratio by 25%, thus promoting osteogenesis (26).
Additionally, the bidirectional modulation of the Wnt/β-Catenin pathway influences osteoblast function and bone metabolism in patients receiving immune checkpoint inhibitors (ICIs) (see Table 4). Suppression of Wnt signaling significantly reduces bone mass (BV/TV by approximately 60%) and bone strength (by 33%), suggesting the need for bone-protective interventions during ICI therapy (27). Conversely, DKK1 can relieve Wnt inhibition, increasing osteoblast density (2.4-fold), osteoblast surface coverage (by 65%), and bone mineral density (by 9%). When combined with anti-PD-1 therapy, it further enhances antitumor immunity (28, 29), as β-catenin not only promotes osteogenesis but also induces PD-L1 expression (3–4-fold increase), contributing to a “self-limiting” immunosuppressive feedback loop. PD-1/PD-L1 blockade can reverse this suppression (30). From an ICI-centered perspective, PD-1 receptor activation promotes osteogenic markers via the ERK/β-catenin signaling pathway (ALP ↑ 61%, OCN ↑ 54%, mineralization ↑ 83%), whereas PD-1 blockade may weaken this osteogenic effect (10). Therefore, in cancer patients receiving ICIs, bone metabolism assessment must take into account the net outcome of these bidirectional regulatory mechanisms.
Nevertheless, clinical evidence suggests that the primary bone-related effects of ICIs stem from inhibition of osteoclast activity and indirect promotion of bone formation, rather than direct osteoblast activation (4). These findings offer important implications for future osteoporosis management strategies.
Figure 2 provides an integrated overview of the bidirectional regulatory mechanisms of the PD-1/PD-L1 axis in bone metabolism and the effects of its inhibition. It illustrates how the PD-1/PD-L1 pathway interacts with multiple signaling axes to regulate both osteoclasts and osteoblasts, and highlights the temporal differences in short- vs. long-term blockade, the influence of the bone microenvironment, and crosstalk with the Wnt/β-Catenin pathway.
Bidirectional regulation of bone metabolism by the PD-1/PD-L1 pathway and the mechanisms of its inhibitors. This figure illustrates the regulatory network of the PD-1/PD-L1 pathway in bone metabolism. The left panel represents osteoclast regulation, while the right panel shows osteoblast regulation. Short-term administration of PD-1/PD-L1 inhibitors reduces osteoclast activity by approximately 60% via inhibition of the JNK signaling pathway, whereas long-term use is associated with an increased risk of fractures (hazard ratio ≈ 1.6). The PD-1/PD-L1–PKM2 axis plays a pivotal role in osteoblast differentiation: PD-L1 enhances osteogenesis by upregulating RUNX2 expression, while PKM2 activation suppresses osteoblast function. The Wnt signaling pathway exerts dual effects—its canonical branch inhibits osteoclastogenesis.
Challenges
5
Contradictory clinical findings and incomplete mechanistic understanding
5.1
Current clinical observations on the impact of PD-1/PD-L1 inhibition on bone metabolism yield conflicting results, and the underlying mechanisms remain poorly elucidated. Differences in clinical characteristics may partly explain these inconsistencies (see Table 5). Age and sex are major determinants: elderly female patients treated with ICIs have a significantly increased risk of fractures (21, 31), and baseline bone mineral density (BMD) has been identified as a critical predictor—patients with lower BMD derive less benefit from PD-1 blockade (32). Additionally, treatment regimens (monotherapy vs. combination therapy), tumor type, and tumor burden also influence skeletal outcomes, sometimes resulting in opposing bone metabolic responses (4, 5). These contradictions highlight the complexity of PD-1/PD-L1-mediated regulation of bone metabolism and underscore the urgent need for further research.
Lack of clinical monitoring and interventions targeting bone metabolism
5.2
Although ICIs are significantly associated with adverse skeletal events—including pathological fractures, vertebral compression fractures, and femoral neck fractures (14)—these complications are often under-recognized in clinical practice. Reports indicate that up to 69% of patients may experience bone-related adverse events, with some requiring extended treatment intervals or premature discontinuation of cancer therapy (15). These are not isolated cases but rather reflect a widespread oversight in clinical monitoring (see Table 6). Multiple studies (4, 7) reveal a lack of baseline BMD evaluation and exceedingly low usage of bone-protective agents—fewer than 10% of patients receive treatment for osteoporosis (14, 21). Such gaps in screening and intervention may directly contribute to the occurrence of multiple fractures and bone resorptive lesions (8). Nevertheless, osteoporosis and fragility fractures are not yet formally recognized as immune-related adverse events (irAEs) (16). Baseline and longitudinal assessments of skeletal health are essential and should be systematically implemented in patients undergoing ICI therapy (33).
Discussion
6
Both clinical and mechanistic studies have reported contradictory findings regarding the role of the PD-1/PD-L1 pathway in bone metabolism regulation (4, 6, 10, 18). These discrepancies highlight the complexity of this pathway and the bidirectional effects of its inhibitors on bone homeostasis, suggesting a potential bone-protective role for PD-1/PD-L1 blockade. Compared with traditional bone-protective agents (Table 7), PD-1/PD-L1 inhibitors act further upstream in the RANKL axis (5), exerting a dual regulatory effect—initial inhibition of osteoclastogenesis followed by promotion of osteogenesis (4). This complements the unidirectional effects of conventional agents such as bisphosphonates and denosumab (34, 35).
Notably, combining PD-1 inhibitors with denosumab has been shown to increase the objective response rate in tumors (from 37% to 54%) and reduce the incidence of skeletal-related events by approximately 30% (36). Similarly, their combination with zoledronic acid can synergistically enhance anti-tumor immunity and anti-osteoporotic effects through the activation of γδ T cells (37). These findings suggest that PD-1/PD-L1 inhibitors may offer a novel therapeutic avenue for osteoporosis prevention and the management of skeletal-related adverse events (SREs) (14, 15).
As evidence continues to accumulate, it is anticipated that within the next five years, osteoporosis and fragility fractures will be increasingly recognized as part of the spectrum of immune-related adverse events (irAEs) associated with immune checkpoint inhibitors (ICIs) (16). Looking further ahead, over the next decade, the integration of bone-targeted strategies with ICI-specific bone-protective protocols may emerge (33), aiming to balance anti-tumor efficacy with bone health preservation and reduce the risk of SREs.
In the coming 5–10 years, an evidence-based framework for comprehensive skeletal health monitoring and stratified prevention is expected to be developed and refined. As summarized in Table 8, several emerging findings support this trend: baseline CT-derived BMD values have been shown to correlate with survival outcomes following ICI therapy (32), suggesting that CT-BMD should be incorporated into pre-treatment screening. The newly developed “Mel-ICI Fracture Score” can help identify high-risk individuals, with scores ≥4 warranting the initiation of bone-protective agents (31). Given that fracture risk increases 2.6-fold within the first year of ICI treatment, DXA assessments are recommended at therapy initiation (21). Moreover, AI-assisted QCT techniques, coupled with dual-track monitoring of BMD changes and serum bone turnover markers, can enable precise and timely surveillance of bone health (4, 9).
The establishment of a “Bone-Oncology Joint Clinic” model is also proposed, facilitating a closed-loop management approach—from baseline evaluation to regular follow-up and comprehensive intervention (38), as well as stratified care for patients with bone metastases (39). These innovations aim to address bone loss and provide continuity of care for patients who develop skeletal complications during ICI treatment (15), ultimately mitigating poor prognostic outcomes (7). Collectively, these strategies may enable dual optimization of tumor control and skeletal health during ICI therapy and significantly improve the quality of life and long-term health outcomes of cancer survivors.
whereas the non-canonical branch promotes osteoclast formation. Clinical observations suggest that immune checkpoint inhibitors primarily promote bone formation indirectly by suppressing osteoclast activity rather than by directly enhancing osteoblast differentiation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Wang BC Cao RB Li PD Fu C. The effects and safety of PD-1/PD-L 1 inhibitors on head and neck cancer: A systematic review and meta-analysis. Cancer Med. (2019) 8:5969–78. doi: 10.1002/cam 4.2510, PMID: 31436392 PMC 6792498 · doi ↗ · pubmed ↗
- 2Abaza A Sid Idris F Anis Shaikh H Vahora I Moparthi KP Al Rushaidi MT. Programmed cell death protein 1 (PD-1) and programmed cell death ligand 1 (PD-L 1) immunotherapy: A promising breakthrough in cancer therapeutics. Cureus. (2023) 15:e 44582. doi: 10.7759/cureus.44582, PMID: 37667784 PMC 10475160 · doi ↗ · pubmed ↗
- 3Greisen SR Kragstrup TW Thomsen JSHørslev–Pedersen K Lund Hetland M Stengaard–Pedersen K. The programmed death-1 pathway counter-regulates inflammation-induced osteoclast activity in clinical and experimental settings. Front Immunol. (2022) 13:773946. doi: 10.3389/fimmu.2022.773946, PMID: 35356000 PMC 8959817 · doi ↗ · pubmed ↗
- 4Gassner T Chittilappilly C Pirich T Neuditschko B Hackner K Lind J. Favorable impact of PD 1/PD-L 1 antagonists on bone remodeling: an exploratory prospective clinical study and ex vivo validation. J Immunother Cancer. (2024) 12:e 008669. doi: 10.1136/jitc-2023-008669, PMID: 38702145 PMC 11086513 · doi ↗ · pubmed ↗
- 5Wang K Gu Y Liao Y Bang S Donnelly CR Chen O. PD-1 blockade inhibits osteoclast formation and murine bone cancer pain. J Clin Invest. (2020) 130:3603–20. doi: 10.1172/JCI 133334, PMID: 32484460 PMC 7324182 · doi ↗ · pubmed ↗
- 6Jeon SM Lim JS Park SH Kim HJ Kim HR Lee JH. Blockade of PD-L 1/PD-1 signaling promotes osteo-/odontogenic differentiation through Ras activation. Int J Oral Sci. (2022) 14:18. doi: 10.1038/s 41368-022-00168-2, PMID: 35365595 PMC 8976080 · doi ↗ · pubmed ↗
- 7Pantano F Tramontana F Iuliani M Leanza G Simonetti S Piccoli A. Changes in bone turnover markers in patients without bone metastases receiving immune checkpoint inhibitors: An exploratory analysis. J Bone Oncol. (2022) 37:100459. doi: 10.1016/j.jbo.2022.100459, PMID: 36338920 PMC 9633734 · doi ↗ · pubmed ↗
- 8Moseley KF Naidoo J Bingham CO Carducci MA Forde PM Gibney GT. Immune-related adverse events with immune checkpoint inhibitors affecting the skeleton: a seminal case series. J Immunother Cancer. (2018) 6:104. doi: 10.1186/s 40425-018-0417-8, PMID: 30305172 PMC 6180387 · doi ↗ · pubmed ↗