DNA methylation differences stratified by normalized fetal/placental weight ratios suggest neurodevelopmental deficits in neonates with congenital heart disease
Marin Jacobwitz, Michael Xie, Jamie Catalano, Ingo Helbig, J. William Gaynor, Nancy Burnham, Rebecca L. Linn, Juliana Gebb, Mark W. Russell, Hakon Hakonarson, Barbara H. Chaiyachati, Ana G. Cristancho

TL;DR
This study explores DNA methylation differences in neonates with congenital heart disease, linking unbalanced fetal/placental weight ratios to potential neurodevelopmental issues.
Contribution
The study introduces a novel approach to linking DNA methylation patterns with fetal/placental weight ratios to predict neurodevelopmental outcomes in CHD neonates.
Findings
Differential methylation was observed in gene sets related to brain development and neurogenesis.
Methylation differences were found between unbalanced and balanced fetal/placental weight ratio groups.
Gene pathways associated with neurodevelopmental dysfunction were identified in CHD neonates.
Abstract
We lack early biomarkers for predicting neurodevelopment (ND) outcomes in children with congenital heart disease (CHD). Placentas of fetuses with CHD have abnormalities, including unbalanced fetal/placental weight ratios (F/P). Although DNA methylation profiles have revealed insights into the maternal-fetal environment (MFE), it is unknown if DNA methylation correlates to normalized F/P weight ratio groups and how these differences relate to ND outcomes. We prospectively recruited a cohort of pregnant women carrying a fetus with CHD. A subset of the cohort had DNA methylation performed on either umbilical cord blood or postnatal blood (45 full-term neonates). We calculated normalized F/P weight ratios, focusing on three normalized F/P ratio groups for analysis. We calculated differential methylation signals in eight ND disabilities-associated gene sets. Normalized F/P ratios were…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
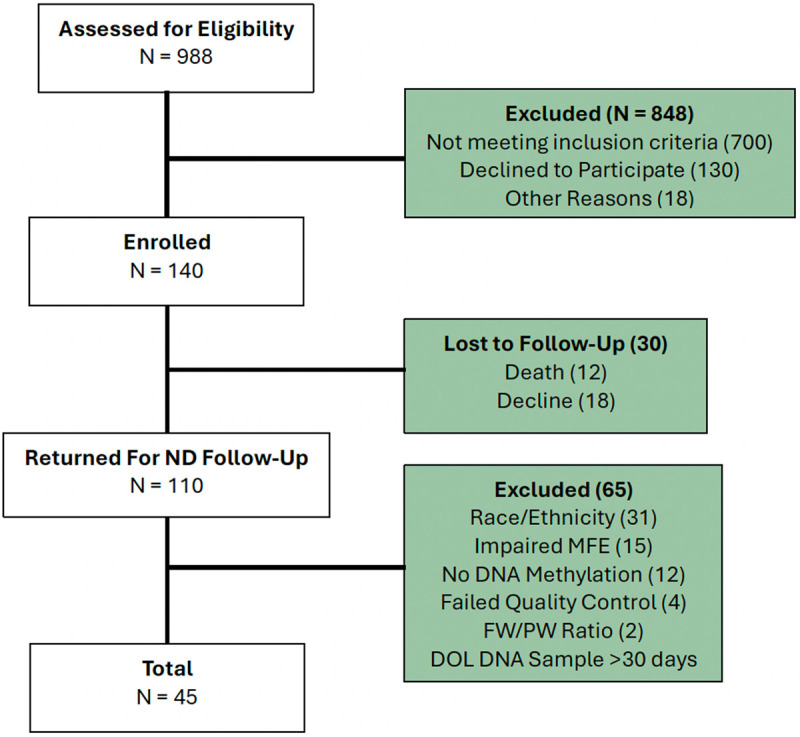 Fig 1
Fig 1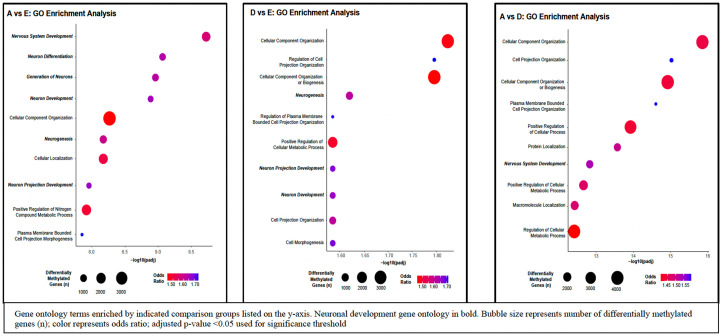 Fig 2
Fig 2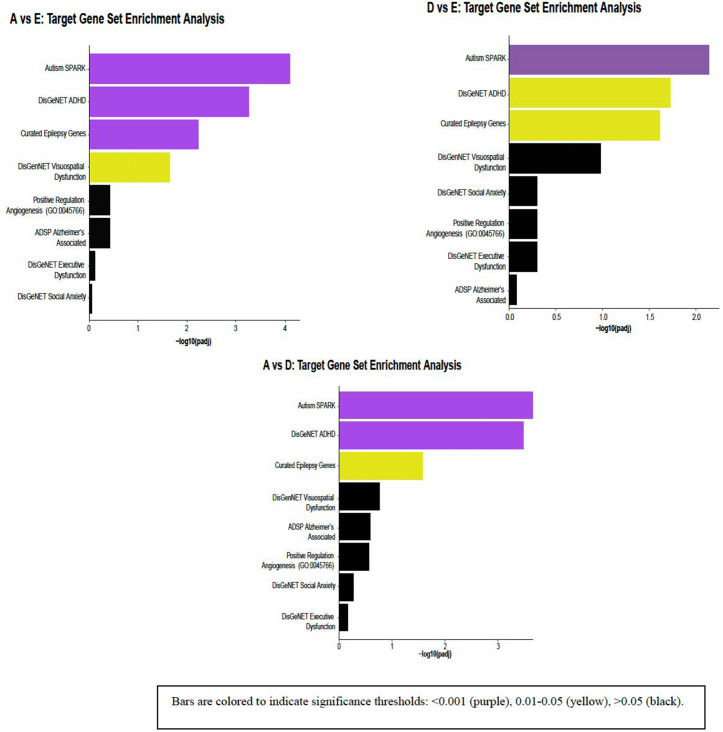 Fig 3
Fig 3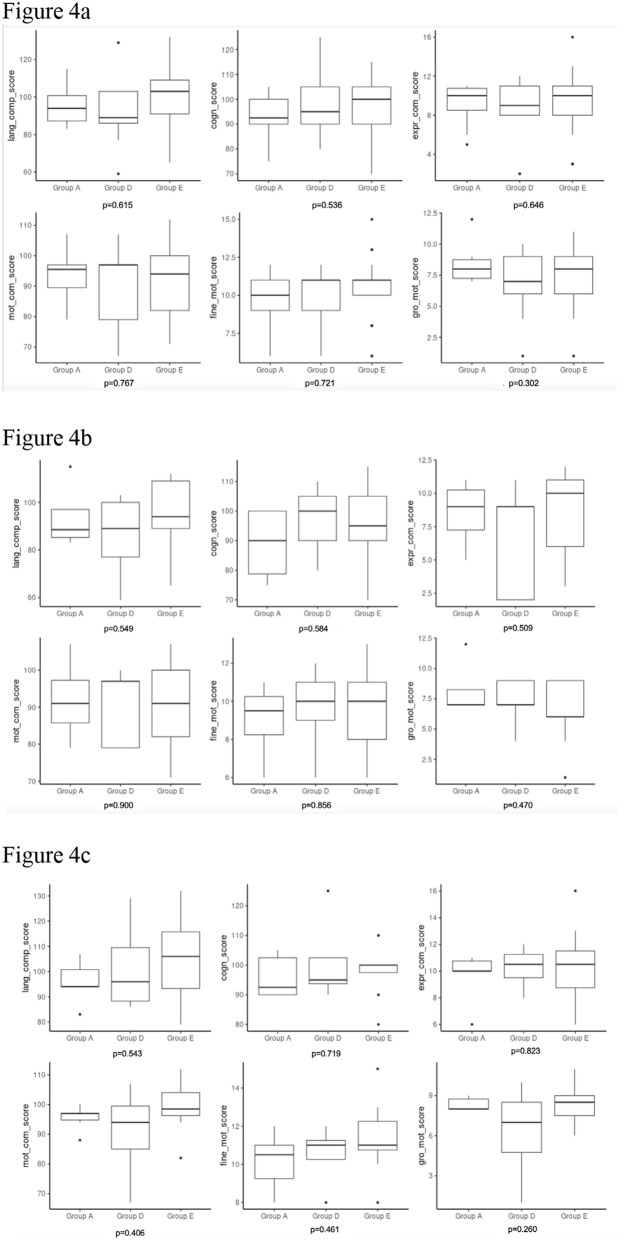 Fig 4
Fig 4- —http://dx.doi.org/10.13039/100001340W. W. Smith Charitable Trust
- —http://dx.doi.org/10.13039/100019274National Institute of Mental Health and Neurosciences
- —http://dx.doi.org/10.13039/100006279St. Jude Medical
- —http://dx.doi.org/10.13039/100000065National Institute of Neurological Disorders and Stroke
- —http://dx.doi.org/10.13039/100000065National Institute of Neurological Disorders and Stroke
- —http://dx.doi.org/10.13039/100000065National Institute of Neurological Disorders and Stroke
- —http://dx.doi.org/10.13039/100000867Robert Wood Johnson Foundation
- —http://dx.doi.org/10.13039/100000065National Institute of Neurological Disorders and Stroke
- —http://dx.doi.org/10.13039/100009633Eunice Kennedy Shriver National Institute of Child Health and Human Development
- —http://dx.doi.org/10.13039/100006279St. Jude Medical
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCongenital Heart Disease Studies · Congenital heart defects research · Congenital Diaphragmatic Hernia Studies
Introduction
Congenital heart disease (CHD) is the most common birth defect, impacting approximately nine per 1,000 live births, with three per 1,000 having critical CHD requiring catheter-based intervention or surgery in the neonatal period to ensure survival [1,2]. With improvements in surgical and medical outcomes, children with CHD are now surviving well into adulthood, which has allowed for longitudinal neurodevelopmental surveillance [2,3]. It has become apparent that many survivors of CHD have an array of neurodevelopmental disabilities impacting their quality of life and adult independence, including autism, visuospatial deficits, attention deficit-hyperactivity disorder (ADHD), and epilepsy [2–7]. Genetic predisposition may account for some, but not all, of the adverse neurodevelopmental outcomes, as only 25–40% of children with CHD have extracardiac anomalies or an identified genetic syndrome [8], and the currently known postnatal, medical, and surgical risk factors only account for approximately 30% of the variation in these long-term outcomes [9,10]. Although earlier theories focused on neonatal surgery as the primary culprit contributing to adverse neurodevelopmental outcomes, recent research has identified that an adverse in-utero environment, including abnormal placentology, may be the foundation of brain vulnerability with additive insults postnatally [11–14].
The placenta is a highly vascular transient organ that acts as an interface for oxygen and nutrient delivery from the mother to the fetus [15,16]. The placenta has been increasingly implicated in adverse neurodevelopmental outcomes in CHD, with many studies identifying high rates of vascular and structural abnormalities in postnatal placental pathology in CHD pregnancies [15,17–19]. The severity of placental abnormalities in postnatal pathology has been correlated with smaller neonatal brain volumes, echoing the role of the placenta in fetal brain growth [17]. Neonates with CHD and superimposed placental dysfunction from gestational hypertension, preeclampsia, growth restriction, and pre-term birth demonstrated increased length of hospital stay and higher mortality than neonates with CHD and healthy placentas [20]. One theory is that the placenta in CHD has a structurally abnormal phenotype, leading to the inadequate oxygenation of fetal blood, with subsequent lower oxygen saturation of blood from the placenta [21]. In the already vulnerable fetus with CHD, this theory results in lower cerebral oxygen delivery, poor brain development, and ultimately impaired neurodevelopment [21]. With ongoing evidence that many survivors of CHD have neurodevelopmental disabilities regardless of cardiac lesion type, investigations into the influence of the maternal-fetal environment (MFE) and placenta on longitudinal neurodevelopmental outcomes are increasingly being investigated [22,23].
Categorization of fetal to placental ratio (hereafter F/P) into either “balanced,” which is normal growth of the fetus and the placenta relative to one another, or “unbalanced,” describing discrepant growth of the fetus and placenta, can be used in lieu of merely evaluating the growth of the fetus or the placenta as separate entities. For example, using a 9-block categorization based on placental weight and F/P weight ratio z-scores to create normalized F/P weight ratios, previous research has identified that there was an increased risk of perinatal death with unbalanced F/P growth compared to balanced F/P growth [24]. Furthermore, using placental weight and F/P weight ratio z-scores to classify normalized F/P weight ratios into clinically relevant categories allows for a better understanding of how the fetus and the placenta are growing relative to one another and, therefore, the health of the pregnancy [24].
To specifically understand the importance of normalized F/P weight ratios, here we focus on the relationship between normalized F/P weight ratios and the fetal and neonatal DNA methylation profile. The health of the placenta likely impacts the epigenetic landscape of the fetus and newborn infant [25,26]. Epigenetic regulation of gene expression is a well-regulated process necessary for normal development. Dysregulation of the epigenome can result in pathology, including neurodevelopmental disorders [27–29]. Specifically, DNA methylation has been identified as an important part of epigenetic regulation in the placenta, with dynamic changes throughout gestation that may influence fetal and placental growth and function [26,28–31]. The objective of this paper was to identify if there are differences in DNA methylation of fetal or infant blood between balanced and unbalanced normalized F/P weight ratio groups in a cohort of neonates with CHD. Gene pathways associated with common neurodevelopmental dysfunction identified in survivors of CHD were specifically examined to determine if there may be a clinical relevance to these normalized F/P weight ratios and relevant outcomes.
Methods
Subject cohort
The cohort was prospectively recruited for a study focusing on environmental exposure influence on neurodevelopmental outcomes in neonates with CHD and has been previously published (hereafter referred to as “parent study”) [9]. Written informed consent was obtained from all pregnant patients, and the study was approved by the Institutional Review Board at The Children’s Hospital of Philadelphia. As part of the parent study, umbilical cord blood was collected from the neonates at the time of birth. If umbilical cord blood was unable to be collected, a postnatal blood sample was obtained. A subset of the cohort had DNA methylation profiles performed using Infinium MethylationEPIC BeadChip V1 (850K) microarray [32] in the Center for Applied Genomics at The Children’s Hospital of Philadelphia following the standard protocol from Illumina. Neurodevelopmental testing was completed at 18 months of age using The Bayley Scales of Infant and Toddler Development-III (BSID-III), which provide composite scores for motor, language, and cognitive skills, with higher scores indicating better skills [33]. For statistical analysis purposes, the neurodevelopmental scores were categorized by sex assigned at birth. Data for this study was accessed on April 4, 2023, for research purposes. For this study, the authors did not have access to information that could identify individual participants during or after data collection.
Inclusion/exclusion criteria
Inclusion criteria for the parent study were infants with CHD and expected cardiopulmonary bypass surgery at age 44 weeks post-conception or younger [9]. Subjects with an identified genetic syndrome, major extracardiac anomaly, or primary language other than English were excluded [9]. A total of 988 pregnant women were screened for eligibility in the parent study, with 848 subjects excluded for not meeting parent study inclusion criteria, declined to participate in the parent study, and other reasons in the parent study [9]. 140 pregnant women were enrolled in the parent study, with 110 infants (79%) returning at 18 months for neurodevelopmental (ND) follow-up. Additional inclusion and exclusion criteria were applied for this study based on principal component and methylation age analyses. Reported race and ethnicity have been shown to be associated with differential DNA methylation patterns, including differences in methylation in cord blood [34], consistent with our principal component analysis (S1 Fig). We therefore excluded subjects that did not identify as non-Hispanic white (N = 31) as we were underpowered for downstream analysis. Of the remaining 79 subjects, 12 subjects did not have DNA methylation profiles and were therefore excluded. Given the known impact of maternal co-morbidities [35,36] and cigarette smoking [37] on DNA methylation, all subjects with an impaired maternal-fetal environment (MFE), defined as pre-existing maternal comorbidities, gestational comorbidities, and cigarette use, were excluded (N = 15). All subjects with samples collected >30 days postnatally were excluded (N = 1). Of the remaining 51 subjects, 2 subjects were not within the three F/P weight ratio groups of interest, and 4 subjects failed quality control. Our final cohort consisted of 45 subjects (Fig 1). A comparison of umbilical cord blood and postnatal neonatal blood DNA samples of the 45 subjects revealed no significant differences in cell type composition (S2 Fig); therefore, we included subjects that had either umbilical cord blood or postnatal DNA samples. Samples with a mismatch between sex determined by their methylation array data and self-reported sex were also excluded.^33–36^
Cohort flow diagram.
Fetal/placental weight ratio categorization
The normalized fetal/placental (F/P) weight ratios were calculated for all subjects using the method previously reported by Matsuda et al. (2018) [24]. Matsuda et al. (2018) utilized placental weight and F/P weight ratio z-scores to establish a 9-block categorization of normalized F/P ratios to better understand F/P growth and perinatal outcomes [24]. Subjects with normalized F/P weight ratios that were one of the following three categories were of interest and therefore included in this study: inappropriately light placenta with a relatively heavy infant (Group A), light placenta with relatively balanced growth of infant (Group D), and balanced growth of both placenta and infant (Group E) [24]. All other F/P categories were excluded due to sample size (Group F, N = 1; Group B, N = 1).
Infinium MethylationEpic BeadChip data processing and quality control
Minfi R package (Version 3.18, ref [13], R version 4.3) was the primary data analysis pipeline. Raw idat files were imported and digitalized using “read.metharray.exp” function with all default parameters. We removed any poor-quality probes (detection p-value ≥ 0.01) and any sample with poor-quality probe rate (more than 5% of total probes). We removed cross-reactive probes [38]. Sex chromosomes were removed from the analysis to reduce sex-related variation given the small sample size. Quantile normalization method (“preprocessQuantile” function) was then applied to minimize unwanted variation between samples and reduce potential batch effects. The beta value was calculated using “getBeta” function and then logit transformed to M value using the “getM” function. M values for 683083 CpG probes were retained for follow-up statistical analysis.
DNA methylation differential analysis
After initial quality control and export of M values after normalization, the association between the three normalized F/P weight ratio categories, as described above, and probe methylation were evaluated. Linear models were fit for each probe (“lmFit” function in limma, version 3.54) with “sex” (“M” or “F”) and biospecimen source (“umbilical cord blood” or “whole blood DNA”) as covariates. All test statistics were modified by empirical Bayes procedure to protect against test statistic inflation [39]. Each contrast was studied and reported separately.
Gene set enrichment analysis
Based on results from differential analysis, a list of methylation sites with p-value < 0.05 was generated. The functional enrichment characteristics among those sites were evaluated by Gene Ontology annotation within Illumina Epic Array (“gometh” function in missMethyl, version 1.24). Additionally, eight gene sets were targeted based on commonly identified neurodevelopmental disabilities previously reported in the CHD population [2,4,5,7,40–42]: “Alzheimer’s,” [43] “autism,” [44] “epilepsy,” [45] “Attention Deficit Hyperactive Disorder (ADHD),” [46] “executive dysfunction,” [46] “social anxiety,” [46] “visuospatial dysfunction,” [46] and “positive regulation of angiogenesis” [10,47] gene sets. We utilized “methylgometh” function (“methylGSA” package, version 1.8) to measure gene set enrichment among our methylation sites. “methylgometh” function adjusts the number of CpGs for each gene by weighted resampling and Wallenius non-central hypergeometric approximation. We report p-value and Benjamini-Hochberg false discovery rate (FDR) adjusted p-values for each targeted gene set we curated for this study.
Placental groups and BSID-III
The cohort was assessed as a full cohort and then stratified by sex, as neurodevelopmental outcomes demonstrate sex differences [48]. We then examined the differences between the normalized F/P weight ratio groups and the BSID-III for language, motor, and cognition. ^50^A one-way ANOVA test (aov function, R stats version 3.6.2) was employed to test the significance of the association. Each comparison was tested and reported separately. For the association between methylation rate and neurodevelopment, we used a mean of 100 and SD of 15 [49], and categorized the cohort into three groups: within 1 SD of the mean (85–115) is normal, 0.5 SD below 85 is mild impairment, and ≤2 SD below 85 is severe impairment.
Results
Demographic and clinical characteristics of the cohort are in Table 1. Forty-nine percent (22/45) of the cohort was male (Group A 4/11, 36%; Group D 5/11, 45%; Group E 13/23, 57%). The most common cardiac lesion was transposition of the great arteries (TGA) in 38% (17/45) of subjects, followed by 33% (15/45) with hypoplastic left heart syndrome (HLHS). The average gestational age (GA) at birth was 39.23 weeks (SD 0.75 weeks). Forty percent (18/45) of the cohort had methylation profiles on umbilical cord blood sample; the remaining 60% had methylation microarrays performed on postnatal peripheral blood sample. Average day of life collection of blood samples was 5.7 days (SD 5.3 days; Table 1). The number of CpG sites detected for each normalized F/P ratio group using a significance threshold cutoff of 0.05 is detailed in Table 1. Data files with entire ranked CpG lists are provided as S1–S3 Appendices files to this manuscript, labeled by the group comparison. In addition, gene ontology analysis using genes normalizing for numbers of probes per genes at significant CpG sites were identified for further pathway analysis (number of genes listed in Table 1).
Table 1: Demographics and clinical characteristics.
Placental groups and gene ontology enrichment analysis
An unbiased gene ontology enrichment analysis of differential methylation regions revealed enrichment for pathways related to brain development and neuronal function. Normalized F/P weight ratio Group A (inappropriately light placenta with relatively heavy fetus) and normalized weight ratio Group E (normal) were enriched for differential methylation signals in the following pathways: nervous system development (GO:0007399), neuron differentiation (GO: 0030182), generation of neurons (GO:0048699), neuron development (GO:0048666), synapse (GO:0007399), and neuron projection (GO:0043005; Fig 2); normalized F/P weight ratio Group D (light placenta with relatively balanced growth of fetus) and normalized F/P weight ratio Group E (normal) were enriched for differential methylation signals in the following pathways: neurogenesis (GO:0022008), regulation of neuron projection development (GO:0010975), neuron projection development (GO:0031175), neuron development (GO:0048666), and generation of neurons (GO:0048699; Fig 2); normalized F/P weight ratio Group A (inappropriately light placenta with relatively heavy fetus) and normalized ratio Group D (light placenta with relatively balanced growth of fetus) were enriched for differential methylation signals in the nervous system development (GO:0007399) and neurogenesis (GO:0022008; Fig 2) pathways.
Placental Groups and Gene Ontology Enrichment Analysis, Gene ontology terms enriched by indicated comparison groups listed on the y-axis.Neuronal development gene ontology in bold. Bubble size represents the number of differentially methylated genes (n); color represents odds ratio. We report p-value and Benjamini-Hochberg false discovery rate (FDR) adjusted p-values for each targeted gene set.
Placental groups and ND-associated gene pathways
When comparing normalized F/P weight ratio Group A (inappropriately light placenta with relatively heavy fetus) to normalized F/P weight ratio Group E (normal), there were significant differences in methylation in the following gene pathways: autism (adjusted p < 0.0001), ADHD (adjusted p < 0.001), epilepsy (adjusted p < 0.01), and visuospatial dysfunction (adjusted p < 0.05; Fig 3).
ND-associated Gene Pathways by Placental Ratio Groups, color-coded based on significance.We report p-value and Benjamini-Hochberg false discovery rate (FDR) adjusted p-values for each targeted gene set. Bars are colored to indicate significance thresholds: < 0.001 (purple), 0.01-0.05 (yellow), > 0.05 (black).
Comparison of normalized F/P weight ratio Group D (light placenta with relatively balanced growth of fetus) to normalized F/P weight ratio Group E (normal) similarly identified significant methylation differences in autism (adjusted p < 0.01), ADHD (adjusted p < 0.05), and epilepsy (adjusted p < 0.05) gene pathway sets (Fig 3).
Significant methylation differences were also seen in autism (adjusted p < 0.0001), ADHD (adjusted p < 0.0001), and epilepsy (adjusted p < 0.05) gene pathways when comparing normalized F/P weight ratio Group A (inappropriately light placenta with relatively heavy fetus) to normalized F/P weight ratio Group D (light placenta with relatively balanced growth of fetus; Fig 3). Significant genes driving enrichment in gene pathways for normalized F/P weight ratio groups are detailed in S1 Table.
Placental groups and BSID-III
For the full cohort, there were no significant differences between the normalized F/P weight ratio groups in the BSID-III scales of neurodevelopment for language (p = 0.615), motor (p = 0.767), or cognition (p = 0.536; Fig 4a). Stratified by sex assigned at birth, for males, there were no significant differences between the normalized F/P weight ratio groups in the BSID-III scales of neurodevelopment for language (p = 0.549), motor (p = 0.900), or cognition (p = 0.584; Fig 4b). For females, there were also no significant differences between the normalized F/P weight ratio groups in the BSID-III scales of neurodevelopment for language (p = 0.543), motor (p = 0.406), or cognition (p = 0.719; Fig 4c).
Box plot showing correlation of the Bayley Scales of Infant Developed (BSID-III) with F/P weight ratios (4a full cohort, 4b male, 4c female).
Stratified by CHD lesion type, there was no significant difference in BSID-III scales of neurodevelopment for language (p = 0.128) or cognition (p = 0.122), but there was a significant difference for motor scores (p = 0.005; S3 Fig). There were no significant differences in the total methylation rate and the BSID-III scores categorized by normal, mild impairment, and severe impairment when evaluated as a full cohort, as well as when stratified by sex (S4 Fig).
Discussion
Infants with CHD demonstrated differentially methylated patterns of genes involved in nervous system development between normalized F/P weight ratio groups in support of F/P growth dynamics being relevant to neurodevelopmental outcomes in CHD.^16,55,56^ Furthermore, our data did not show significant methylation differences between normalized F/P weight ratios and BSID-III, but did identify differences in DNA methylation between normalized F/P weight ratio groups in gene pathways specific to common neurodevelopmental dysfunction in survivors of CHD. These results are in line with previous work demonstrating that abnormal placental pathology is related to perinatal, neonatal, and neurodevelopmental outcomes in both healthy pregnancies and pregnancies with CHD [15,50,51].
Gene ontology enrichment analysis revealed a potential link between the placenta and neurodevelopment, with significant differential methylation identified between the normalized F/P weight ratio groups in multiple genes involved in various stages of neurogenesis. Notably, we uncovered this difference in peripheral methylation patterns by unbiased analysis. It is well known the importance of adequate nutrition and perfusion from the placenta to the growing fetus to ensure optimal neurodevelopment [52]. Our data supports that abnormal placentation may influence pathways related to neurogenesis in the developing fetus. It is unclear how these peripheral changes in methylation correlate to potential disruption of the epigenome within the developing brain, though literature supports relevance between brain and peripheral signatures [53,54]. Taken together, this data suggests that the peripheral epigenetic profile may be providing insight into pathways that contribute to altered brain development in CHD.
Many survivors of CHD have a spectrum of neurodevelopmental disabilities that become more apparent with age, including but not limited to executive dysfunction, visuospatial dysfunction, ADHD, autistic-like phenotypes, and social anxiety [2,3]. As this population is now aging into adulthood, cognitive decline in early adulthood is being increasingly reported across all cardiac lesion phenotypes with features similar to Alzheimer’s disease [23,55–58]. Early neurodevelopmental evaluation, both clinically and for research, is now most commonly with the BSID-III [59,60]. Studies have suggested that the BSID-III may overestimate neurodevelopmental scores in survivors of CHD compared with prior versions of the neurodevelopmental scale [59,60]. Both as a full cohort and categorized by sex, our data did not reveal significant differences in BSID-III scores stratified by normalized F/P weight ratio groups. Given the above concerns about the BSID-III, the lack of differences in the BSID-III scores and the normalized F/P weight ratio groups in our cohort may be because this scale is insensitive for survivors of CHD, with poor correlation with long-term neurodevelopmental outcomes, as it is known that the majority of the CHD population has BSID scores that fall within 0.5–1 standard deviation of the normal population mean [60]. Furthermore, neurodevelopmental difficulties often manifest in later childhood in children with CHD, highlighting the importance of longitudinal neurodevelopmental surveillance [61–63]. Similar to prior studies [64–69], our data also reveal that neonates with HLHS have worse motor scores than the other cardiac lesions. Although our cohort did include neonates with different cardiac lesions, which may impact early neurodevelopmental outcomes [3], there was an overall equal distribution of these lesions across the F/P ratio groups. Therefore, we would expect the effect of CHD lesion type on the association between BSID-III and the F/P ratio groups to be mitigated. Additionally, our cohort was recruited from the same institution, ensuring general consistency in the preoperative, intraoperative, and postoperative management of subjects.
In both human and animal models, there is increasing evidence that there are shared developmental pathways between the brain, heart, and placenta, including but not limited to angiogenic pathways, vascular endothelial growth factor (VEGF), and folic acid-associated pathways [21]. However, there are still substantial gaps in understanding the relationship between the placenta, heart, and brain in CHD-affected pregnancies. Using previously reported gene set pathways for the above commonly reported neurologic dysfunction, significant differential methylation was identified, most notably, between the most unbalanced F/P weight ratio group and the balanced, normal F/P weight ratio group. This data suggests that there may be important epigenetic differences in the unbalanced fetal/placental interface when compared with balanced pregnancies in neonates with CHD.
This study is the first to report differences in DNA methylation using normalized F/P weight ratios in a cohort of neonates with CHD. Normalized F/P weight ratios have been used in other neonatal cohorts, but using these ratios to stratify groups for DNA methylation changes in CHD is novel. There are limitations that are important to acknowledge. First, recruitment for the parent study was not primarily for DNA methylation and required the exclusion of additional subjects to ensure appropriate methylation analyses. The small size of this cohort renders this study statistically underpowered. Our low statistical power and small sample size also required reduced statistical threshold stringency, increasing our risk of false positives. Second, this is a single-center study, and generalizability will be enhanced by data from other institutions or multi-center studies. Third, because of the small sample size of our cohort and the subsequent need to exclude subjects based on demographics, including race and ethnicity, we cannot generalize this to racially and ethnically diverse populations. Fourth, we did not have access to healthy controls for this study and, although there is normative DNA methylation data publicly available, this data does not incorporate the F/P ratio groups limiting its use for our study. Lastly, the other F/P ratio groups in the 9-block categorization system used were underrepresented in our cohort. This is likely, in part, due to the small sample size but also may reflect the incidence of these F/P ratios within the CHD population. This warrants further investigation in future prospective studies with a larger cohort. Thus, this study provides intriguing avenues for longitudinal multi-institutional collaboration to understand if normalized F/P weight ratios and DNA methylation are biomarkers for neurodevelopmental disability in CHD.
Conclusion
Gene ontology enrichment analysis revealed significant methylation differences in normalized F/P weight ratio groups, specifically in genes involved with neurogenesis. Furthermore, our data suggest that there may be notable DNA methylation differences between normalized F/P weight ratio groups in gene pathways implicated in neurodevelopmental disabilities that have been identified in aging survivors of CHD. Taken together, our data highlight the importance of further investigation into how DNA methylation changes in the placenta of fetuses with CHD impact postnatal neurodevelopmental outcomes, including if there is a direct epigenetic link between the placenta and neurogenesis in-utero.
Supporting information
S1 TableSignificant genes driving enrichment for each gene pathway.(PDF)
S1 FigPrincipal component analysis of race.(PDF)
S2 FigCell type composition in umbilical cord blood and postnatal DNA blood sample.(PDF)
S3 FigBSID-III stratified by cardiac lesion type.(PDF)
S4 FigAssociation between total methylation rate and BSID-III (A full cohort, B female, C male) categorized as normal (85–115), mild impairment (0.5 SD below 85), severe impairment (≤2 SD below 85).(PDF)
S1 AppendixRanked CpG lists, labeled by the group comparison.Group A vs Group D CpG rank site list.(CSV)
S2 AppendixRanked CpG lists, labeled by the group comparison.Group A vs Group E CpG rank site list.(CSV)
S3 AppendixRanked CpG lists, labeled by the group comparison.Group D vs Group E CpG site rank list.(CSV)
S1 FileData sharing statement PLOS 5.9.25.(DOCX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Panigrahy A, Schmithorst VJ, Wisnowski JL, Watson CG, Bellinger DC, Newburger JW, et al. Relationship of white matter network topology and cognitive outcome in adolescents with d-transposition of the great arteries. Neuroimage Clin. 2015.10.1016/j.nicl.2015.01.013PMC 431887425685710 · doi ↗ · pubmed ↗
- 2Marino BS, Lipkin PH, Newburger JW, Peacock G, Gerdes M, Gaynor JW, et al. Neurodeve lopmental outcomes in children with congenital heart disease: evaluation and management: a scientific statement from the American Heart Association. Circulation. 2012;126(9):1143–72. doi: 10.1161/CIR.0b 013e 318265 ee 8a 22851541 · doi ↗ · pubmed ↗
- 3Wernovsky G, Licht DJ. Neurodevelopmental Outcomes in Children with Congenital Heart Disease-What Can We Impact? Pediatric Crit Care Med. 2016:1–24.10.1097/PCC.0000000000000800 PMC 497548027490605 · doi ↗ · pubmed ↗
- 4Bellinger DC, Jonas RA, Rappaport LA. Developmental and neurologic status of children after heart surgery with hypothermic circulatory arrest or low-flow cardiopulmonary arrest. J Neurosurg Anesthesiol. 1995;7(3):196.10.1056/NEJM 1995030233209017838188 · doi ↗ · pubmed ↗
- 5Bellinger DC, Wypij D, Du Plessis AJ, Rappaport LA, Jonas RA, Wernovsky G, et al. Neurodevelopmental status at eight years in children with dextro-transposition of the great arteries: The Boston Circulatory Arrest Trial. J Thoracic Cardiovasc Surg. 2003.10.1016/s 0022-5223(03)00711-614666010 · doi ↗ · pubmed ↗
- 6Wypij D, Newburger JW, Rappaport LA, Du Plessis AJ, Jonas RA, Wernovsky G, et al. The effect of duration of deep hypothermic circulatory arrest in infant heart surgery on late neurodevelopment: The Boston Circulatory Arrest Trial. J Thoracic Cardiovasc Surg. 2003.10.1016/s 0022-5223(03)00940-114666011 · doi ↗ · pubmed ↗
- 7Bellinger DC, Wypij D, Rivkin MJ, De Maso DR, Robertson RL Jr, Dunbar-Masterson C, et al. Adolescents with d-transposition of the great arteries corrected with the arterial switch procedure: neuropsychological assessment and structural brain imaging. Circulation. 2011;124(12):1361–9. doi: 10.1161/CIRCULATIONAHA.111.026963 21875911 PMC 3217719 · doi ↗ · pubmed ↗
- 8Richards AA, Garg V. Genetics of congenital heart disease. Curr Cardiol Rev. 2010;6(2):91–7. doi: 10.2174/157340310791162703 21532774 PMC 2892081 · doi ↗ · pubmed ↗