Detection of SXT/R391 integrative conjugative elements carrying tigecycline resistance genes in Shewanella spp. isolated from retail seafood
Wenhui Zhang, Kai Peng, Ming Liu, Xuesong Luo, Zhiqiang Wang, Ruichao Li

TL;DR
This study found tigecycline-resistant Shewanella bacteria in retail seafood, which could spread antibiotic resistance to other bacteria.
Contribution
First report of SXT/R391 ICEs carrying tigecycline resistance genes in Shewanella strains.
Findings
Shewanella was the most common tigecycline-resistant genus isolated from retail seafood.
Two Shewanella strains carried the tet(X4) gene, and one co-harbored tmexCD2-toprJ2 and blaNDM-1.
Tigecycline resistance genes were found on novel SXT/R391 ICEs, which can spread resistance among aquatic bacteria.
Abstract
Tigecycline is a last resort antibiotic that is used to treat serious infections; however, some bacteria have developed tigecycline resistance by producing a tigecycline-inactivating enzyme or tigecycline resistance efflux pump, encoded by tet(X) and tmexCD-toprJ genes, respectively. Tons of seafood are consumed annually in China; however, whether seafood harbors tigecycline-resistant bacteria is not known. In this study, we isolated various tigecycline-resistant bacteria from retail seafood; among these, Shewanella was the predominant tigecycline-resistant genus (33/76, 43.4%). Genomic sequencing revealed that two Shewanella strains carried the tet(X4) gene, while one Shewanella chilikensis strain co-harbored tmexCD2-toprJ2 and blaNDM-1 genes. The tet(X4) and tmexCD2-toprJ2 were found to be located on novel members of the SXT/R391 family of integrated conjugative elements (ICEs). As…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
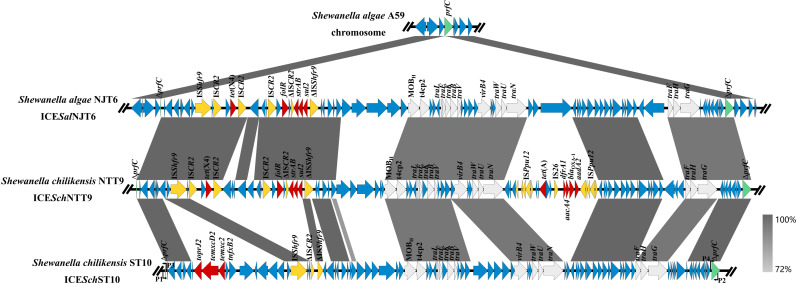 Fig 1
Fig 1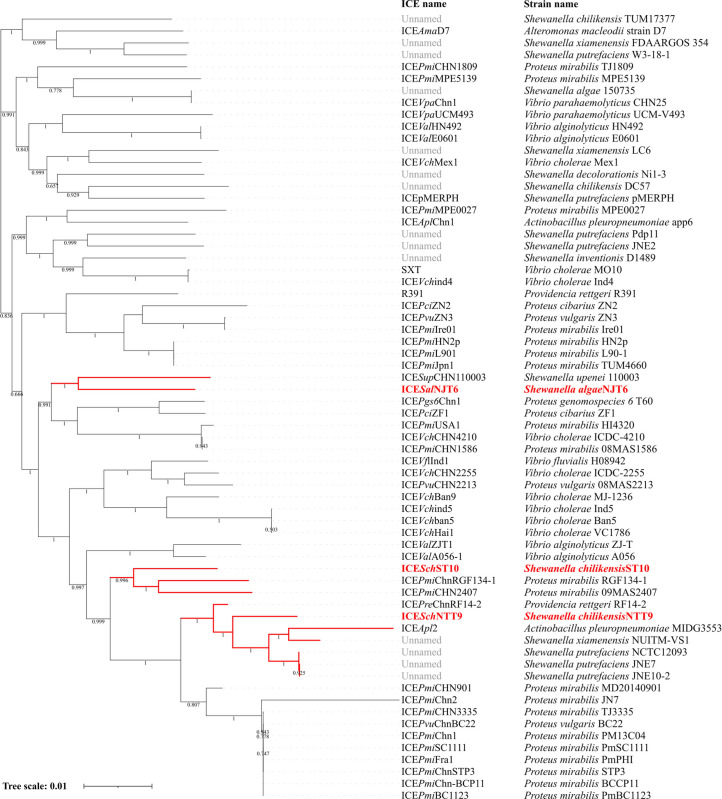 Fig 2
Fig 2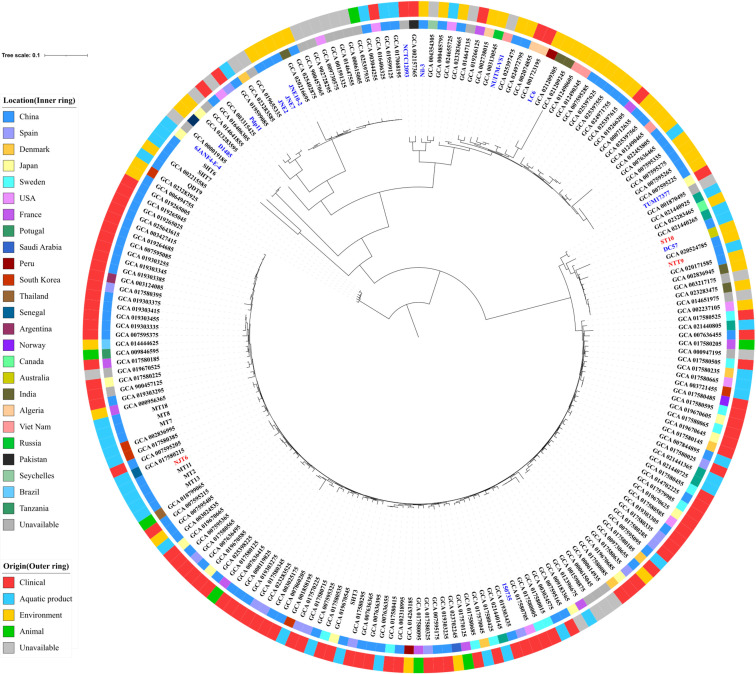 Fig 3
Fig 3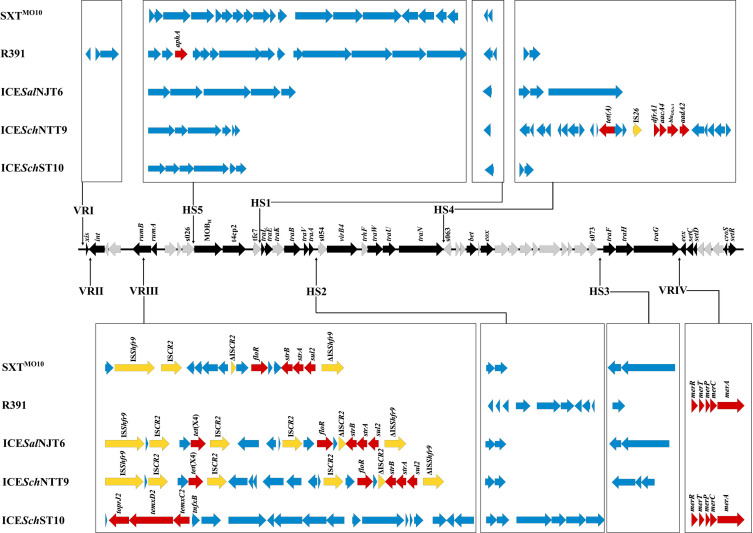 Fig 4
Fig 4| ICE | Strain | Species | Host | Year and location of isolation | Resistance genes in bacterial isolates | Resistance genes in ICE |
|---|---|---|---|---|---|---|
| ICE | NJT6 |
|
| Nanjing city, Jiangsu Province, China, | ||
| ICE | NTT9 |
|
| Nantong city, Jiangsu Province, China, | ||
| ICE | ST10 |
|
| Nanjing city, Jiangsu Province, China, |
|
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
- —National Natural Science Foundation of Chinahttp://dx.doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAntibiotic Resistance in Bacteria · Aquaculture disease management and microbiota · Bacteriophages and microbial interactions
INTRODUCTION
Tigecycline belongs to the glycylcycline class of broad-spectrum antimicrobials. It is considered a last-resort antibiotic for the treatment of severe infections caused by multidrug-resistant (MDR) pathogens, especially carbapenem-resistant pathogenic bacteria (1). However, tigecycline resistance genes, such as tet(X) and tmexCD-toprJ, as well as their variants, have emerged and disseminated among Gram-negative bacteria. To understand the mechanism by which tigecycline resistance genes evolved and disseminated in bacteria, previous studies characterized tigecycline-resistant bacteria, isolated from diverse environments, such as soil, freshwater, freshwater fishes, livestock and poultry farms, retail meats, and hospitals and hospital wastewater (2–5).
China produces over 60 million metric tons of seafood and consumes more than 50 million metric tons of seafood annually. Seafood is recognized as a significant reservoir of antibiotic-resistant bacteria. For example, a Citrobacter meridianamericanus strain and nine Escherichia coli strains, isolated from fishes, clams, and crabs, were found to harbor plasmids containing the tet(X4) gene (6). However, it is widely acknowledged that E. coli and C. meridianamericanus are not indigenous to aquatic environments (7). Therefore, the presence of tigecycline-resistant aquatic bacteria in retail seafood in China remains unknown.
The Shewanella genus comprises gram-negative, facultative anaerobic bacteria that primarily inhabit aquatic environments (8–10). In recent years, there has been a significant rise in Shewanella infections worldwide. Several Shewanella species, such as Shewanella algae, Shewanella putrefaciens, and Shewanella xiamenensis, have been found to be associated with humans (11, 12) and are considered to be opportunistic human pathogens. In addition, Shewanella species carry several antibiotic resistance genes, including carbapenem, beta-lactam (13, 14), and the variants of colistin (mcr) resistance genes (15). However, to date, only the S. xiamenensis strain, isolated from a drainage sample in Vietnam, has been reported to carry the tigecycline resistance gene tet(X4) (16).
SXT/R391 integrated conjugative elements (ICEs) shared a nearly identical integrase Int, which was considered to mediate their site-specific integration into the 5′ end of the prfC gene in the host chromosome. Previously, studies have revealed that SXT/R391 family ICEs contain 52 nearly conserved core genes. In addition, four variable regions (VRI–IV) and five hotspots (HS1–5) have been reported, which are comprised of variable genes conferring a variety of functions including antibiotic resistance. Several studies have investigated the presence of the SXT/R391 ICE family members in a variety of bacterial genera, including Vibrio (17), Proteus (18), Providencia (19), Marinomonas (20), and Shewanella (21). Among the SXT/R391 ICE members, identified in Proteus spp., Pseudomonas aeruginosa, and Providencia rettgeri, only a few were found to carry tigecycline resistance genes, like tet(X6) or tmexCD3-toprJ1b (22–27). Therefore, further research is required to identify the presence of SXT/R391 ICEs harboring tigecycline resistance genes in other bacterial genera.
To investigate the prevalence of tigecycline-resistant bacteria in seafood, we surveyed raw retail seafood sold in the supermarkets of seven cities, namely Wuhan, Baise, Qingdao, Nanjing, Dandong, Nantong, and Shanghai, in China, from 2021 to 2022. We isolated several tigecycline-resistant bacteria belonging to different genera, including Shewanella, Proteus, Morganella, Providencia, Pseudomonas, Pseudoalteromonas, Aeromonas, Citrobacter, Raoultella, Kluyvera, Enterobacter, Myroides, Janthinobacterium, and Serratia. Among these, the majority of tigecycline-resistant bacteria belonged to the genus Shewanella. We identified two Shewanella strains that carried the tet(X4) gene and one Shewanella strain that co-harbored the tmexCD2-toprJ2 and blaNDM-1 genes. Whole genome sequencing (WGS) revealed that the tet(X4) and tmexCD2-toprJ2 genes were located in the SXT/R391 ICE family. The tmexCD2-toprJ2-containing Shewanella species also possessed a novel Salmonella genomic island 1 (SGI1) variant carrying the blaNDM-1 gene. We also isolated eight Shewanella isolates that did not harbor the tet(X) or tmexCD-toprJ gene but were resistant to tigecycline; however, this phenotype was not transferable. Our results suggest that retail seafood carries diverse tigecycline-resistant bacteria and that Shewanella spp. are the predominant tigecycline-resistant bacteria in seafood.
MATERIALS AND METHODS
Isolation and identification of tigecycline-resistant bacterial species
For this analysis, we obtained raw seafood from supermarkets in Wuhan, Baise, Qingdao, Nanjing, Dandong, Nantong, and Shanghai, in China, during 2021 and 2022. Each sample was packaged individually in a sterile bag and placed in an ice box until further analysis. For pretreatment, the sample was treated with 9 mL of 2% NaCl-supplemented Luria-Bertani (LB) medium (LBS) containing 4 µg/mL tigecycline and homogenized in a tissue homogenizer for 3 minutes. Thereafter, 1 mL of the supernatant was transferred to a fresh tube along with LBS broth supplemented with 4 µg/mL tigecycline and cultured for 10 hours at 30°C. The culture was streaked on LBS agar plates containing 4 µg/mL tigecycline and incubated for 10–14 hours at 30°C. Different colonies obtained on the LBS agar plates were purified and subsequently screened for the presence of tet(X) and tmexCD-toprJ genes by polymerase chain reaction (PCR) using specific primers (Table S4). Matrix-assisted laser desorption/ionization-time of flight (Bruker) or 16S rDNA sequencing was used to identify the taxonomy of the bacterial isolates.
Antimicrobial susceptibility testing
The broth microdilution method was used to determine the susceptibility of Shewanella isolates and transconjugants to 16 antimicrobials, namely meropenem, imipenem, tigecycline, colistin, ceftiofur, ciprofloxacin, chloramphenicol, tetracycline, kanamycin, gentamicin, streptomycin, ceftriaxone, aztreonam, amikacin, ampicillin, and amoxicillin-clavulanic acid. The results were interpreted according to the guidelines provided in “non-Enterobacterales” in the Clinical and Laboratory Standards Institute (2020) (28). The breakpoints of tigecycline and colistin were interpreted according to the European Committee on Antimicrobial Susceptibility Testing v12.0 criteria (http://www.eucast.org/clinical_breakpoints/). The E. coli ATCC strain 25922 served as the quality control sample.
Extraction, WGS, and assembly of tigecycline-resistant Shewanella isolates
Genomic DNA of tigecycline-resistant Shewanella isolates was extracted using the FastPure Bacteria DNA Isolation Mini Kit (Vazyme, China). Briefly, the bacterial sample was centrifuged at 13,500 × g for 1 minute, and 2 mL of the bacterial culture was collected and treated with proteinase K and RNase. The genomic DNA was extracted using the column extraction method, and its quality and concentration were evaluated by gel electrophoresis and the Colibri LB spectrophotometer (Titertek-Berthold, Germany), respectively. The whole genomes of Shewanella isolates were sequenced by the Illumina Hiseq 2500 platform (Illumina) using two-paired libraries with 150 bp average length and 150 × coverage. The long-read Oxford Nanopore Technologies (ONT) MinION platform was employed to further sequence the genomes of Shewanella isolates harboring tet(X4) or tmexCD-toprJ genes. The Illumina read data were assembled using SPAdes v3.13.1 at default parameters (29). The hybrid de novo assembly of ONT reads was generated using Unicycler v0.4.8 (30).
Phylogenetic analysis of Shewanella isolates and SXT/R391 ICEs
The phylogenetic relationships of the isolated Shewanella strains were determined using the whole genome sequences of representative Shewanella strains available in the NCBI database (https://www.ncbi.nlm.nih.gov/genbank/). Information on these genome sequences is provided in Table S2. The draft genomes were annotated using Prokka v1.12, and the phylogenetic tree was built using Roary and FastTree based on single-nucleotide polymorphisms (SNPs) in the core genomes (31–33). The distances of SNPs in the core genomes were analyzed using snp-dists v0.7.0 (https://github.com/tseemann/snp-dists). Phylogenetic analysis of the isolated SXT/R391 ICEs and representative ICEs from the NCBI database was performed based on the SNPs in the conserved core genes. Relevant information on the representative ICEs is provided in Table S3. Phylogenetic trees were constructed using the MEGA 11.0 software via the maximum-likelihood method with 1000 replications of bootstrap values (34). The information on conserved core genes used in the phylogenetic trees is provided in Table S5.
Comparative analysis and genetic characterization of the novel SXT/R391 ICEs
The complete genome sequences of Shewanella isolates harboring SXT/R391 ICEs were annotated through the Rapid Annotation using the Subsystems Technology annotation server (http://rast.nmpdr.org/) (35). ResFinder v4.1, PlasmidFinder v2.1, and ISfinder v2.1 were used to identify ARGs, plasmid replicons, and insertion sequences, respectively. The novel SXT/R391 ICEs, identified in this study, were compared with the representative ICEs available in the public database (https://www.ncbi.nlm.nih.gov/genbank/). The conserved and variable regions in novel ICEs were compared with those in representative ICEs, and the genetic context of the tigecycline resistance genes was illustrated. Easyfig v2.2.3 was used to visualize the SXT/R391 ICEs (36).
Conjugation assays
The transferability of tigecycline resistance was determined through mating assays using rifampicin-resistant E. coli C600 as the recipient. Overnight cultures of the donor strain and E. coli C600 were mixed in a 1:1 ratio and incubated overnight at 37°C. The culture was then serially diluted by 10 folds and plated onto selective LB agar plates containing 300 µg/mL rifampicin and 4 µg/mL tigecycline, to count transconjugants. Simultaneously, 10-fold serially diluted cultures were plated onto LB agar plates containing 300 µg/mL rifampicin to count the recipient cells. The plates were incubated overnight at 37°C, and colony-forming units were recorded for three independent replicates. The transfer frequency of ICEs was determined by calculating the number of transconjugants per recipient cell.
Pairwise growth competition assay
For this assay, overnight cultures of the NJT6 or NTT9 (SXT/R391 ICE-bearing E. coli C600) transconjugants and E. coli C600 were diluted to 0.5 McFarland standard and mixed at 1:1 ratio in 5 mL of LB broth. The mixtures were incubated at 37°C for 96 hours with shaking, and 5 µL of culture was reinoculated in 5 mL of fresh LB broth every 24 h. The bacterial cultures were serially diluted by 10-fold and inoculated on LB agar plates in the presence or absence of 4 µg/mL tigecycline. The relative fitness was calculated as follows: w = ln(NRt/NR0)/ln(NSt/NS0), where NR is the number of resistant clones and NS is the number of susceptible clones, with values < 1 indicating the fitness cost.
RESULTS AND DISCUSSION
Sample collection and bacterial identification
In this study, we isolated various bacterial species belonging to Proteus (n = 2), Morganella (n = 8), Providencia (n = 3), Pseudomonas (n = 4), Pseudoalteromonas (n = 4), Aeromonas (n = 7), Citrobacter (n = 6), Raoultella (n = 2), Kluyvera (n = 3), Enterobacter (n = 1), Myroides (n = 1), Janthinobacterium (n = 1), Serratia (n = 1), and Shewanella (n = 33) from retail seafood samples. All the bacterial isolates were screened to detect the presence of tet(X) and tmexCD-toprJ genes using PCR amplification. Except for S. algae NJT6 and Shewanella chilikensis NTT9 carrying tet(X) and S. chilikensis ST10 encoding tmexCD-toprJ, other isolates did not harbor tet(X) or tmexCD-toprJ gene. The strains NTT9 and NJT6 were isolated from Ruditapes philippinarum (manila clams) purchased from Nantong and Nanjing cities, respectively. The strain ST10 was isolated from Sinonovacula constricta (Chinese razor clam) purchased from Nanjing city. Notably, no E. coli strains were isolated from the seafood samples in our analysis, unlike a previous study (6). It is likely that the addition of NaCl in the LBS medium inhibited the growth of E. coli in our analysis.
Resistance phenotype of tigecycline-resistant Shewanella isolates
As Shewanella strains were predominant (33/76, 43.4%) among all the seafood bacterial isolates, we examined whether these strains were resistant to tigecycline. For this analysis, we randomly chose 10 Shewanella strains, in addition to strains NJT6, NTT9, and ST10. Antimicrobial susceptibility testing showed that all 13 Shewanella isolates were MDR strains (Table S1), with similar antibiotic resistance profiles. They exhibited high resistance to tigecycline (n = 13/13, 100%), tetracycline (n = 13/13, 100%), chloramphenicol (n = 13/13, 100%), streptomycin (n = 12/13, 92.3%), ampicillin (n = 12/13, 92.3%), ciprofloxacin (n = 11/13, 84.6%), and aztreonam (n = 8/13, 61.5%). Additionally, these isolates exhibited higher MIC values for imipenem compared to meropenem, but all the isolates were susceptible to amikacin and gentamicin.
We investigated the genomic structures of tet(X) or tmexCD-toprJ genes in the strains NJT6, NTT9, and ST10 by WGS. The sequencing results revealed that the tigecycline resistance genes in strains NJT6, NTT9, and ST10 were located in ICEs, designated as ICESalNJT6, ICESchNTT9, and ICESchST10, respectively (Fig. 1; Table 1). Upon comparison with the genome sequence of S. algae A59, we found that ICESalNJT6, ICESchNTT9, and ICESchST10 were integrated into the 5′ end of the prfC gene (which encodes peptide chain release factor three involved in translational regulation) in these Shewanella genomes.
Genomic map of the ICEs identified in this study. The red and yellow arrows represent the drug resistance genes and insertion sequences, respectively. T4SS-encoding genes were marked by white color, and the insertion site-prfC genes are highlighted in green. Primers P1, P2, P3, and P4 designed to identify the circular form of ICESchST10 were marked as black arrows.
Transferability and fitness cost of SXT/R391 ICEs in tigecycline-resistant Shewanella isolates
A conjugation assay was performed to investigate the transferability of the three SXT/R391 ICE-harboring Shewanella isolates and the other 10 tigecycline-resistant Shewanella strains listed in Table S1. The tet(X4)-carrying ICEs exhibited a very low transfer frequency, ranging from 6.1 × 10^−7^ (strain NJT6) to 4.2 × 10^−7^ (strain NTT9) per recipient cell, indicating the possibility of tet(X4) gene dissemination via SXT/R391 ICEs. The transconjugants were also found to be resistant to antibiotics including chloramphenicol, tetracycline, and streptomycin (Table S1). However, tmexCD2-toprJ2-carrying ICEs (strain ST10) failed to transfer into E. coli C600 under test conditions. These results indicate that ICEs from different Shewanella strains show varying transferabilities. In addition, we found that the other tigecycline-resistant Shewanella strains, which did not harbor the tet(X) or tmexCD-toprJ gene, failed to transfer the tigecycline-resistant phenotype into E. coli C600 under test conditions. WGS revealed no tigecycline resistance genes in the genome sequences of these tigecycline-resistant strains. To investigate the potential role of efflux pumps in mediating tigecycline resistance, we assessed the impact of efflux pump inhibitors on the MICs of the eight non-transferable strains. Treatment with 1-(1-naphthylmethyl)-piperazine (NMP; 100 µg/mL) resulted in a remarkable 32-fold reduction in tigecycline MICs across all strains, while carbonyl cyanide m-chlorophenylhydrazone (100 µg/mL) treatment led to a more modest twofold decrease. These results strongly suggest that efflux pump activity plays a significant role in the observed tigecycline resistance. Notably, the pronounced effect of NMP—a known inhibitor of RND-type efflux pumps—particularly implicates this family of transporters in the resistance mechanism. To further elucidate the specific efflux systems involved, we are currently conducting RNA-seq analyses to identify differentially expressed efflux pump genes under tigecycline selection pressure (data not shown). Moreover, E. coli C600 showed a low competitive advantage over the transconjugants of NJT6 or NTT9 in the pairwise growth competition assay (Fig. S6). These results indicate that ICESalNJT6 and ICESchNTT9 exhibit undetectable fitness costs in host bacteria and could persist in bacteria of other species for a period of time, with the potential for further dissemination.
Phylogenetic relationships between SXT/R391 ICEs and Shewanella isolates
To investigate the phylogenetic relationship of ICESalNJT6, ICESchNTT9, and ICESchST10 with other known ICEs, phylogenetic analysis was performed (Fig. 2). ICESalNJT6 was classified into an independent cluster with ICESupCHN110003, present in Shewanella upenei strain 110003, which was isolated from a stool sample of a patient in Anhui province, China (37). Meanwhile, ICESchNTT9 was classified into a distinct branch adjacent to the cluster composed of ICEApl2 carried by Actinobacillus pleuropneumoniae and uncharacterized ICEs in S. putrefaciens and S. xiamenensis. It is likely that both ICESalNJT6 and ICESchNTT9 harboring tet(X4) have a close evolutionary relationship with ICEs of other Shewanella species. However, ICESchST10 harboring the tmexCD2-toprJ2 gene formed a unique branch along with ICEPmiChnRGF134-1 and ICEPmiCHN2407 carried by Proteus mirabilis. Notably, ICEPmiChnRGF134-1 also carried the tmexCD-toprJ gene cluster. This indicated that ICESchST10 may share the same ancestor with these Proteus ICEs. To the best of our knowledge, this is the first report on the presence of tet(X4)- and tmexCD2-toprJ2-carrying SXT/R391 ICEs in Shewanella species. Altogether, our results indicate that ICESalNJT6, ICESchNTT9, and ICESchST10 are novel members of the SXT/R391 family of ICEs and that SXT/R391 ICEs could be transferred horizontally in different bacterial species.
Phylogenetic analysis of the identified and representative SXT/R391 ICEs. The phylogenetic tree was constructed based on the SNPs in the conserved core genes of the ICEs via the maximum-likelihood method. ICEs identified in this study and the corresponding branches are highlighted in red.
We retrieved genome sequences of Shewanella strains deposited in the NCBI database and found that several uncharacterized SXT/R391 ICEs were encoded by S. algae, S. chilikensis, Shewanella decolorationis, Shewanella hafniensis, Shewanella inventionis, and S. xiamenensis. Further bioinformatics analysis showed that ICEs are not distributed widely among Shewanella species. In addition, SNP analysis revealed that the strains NJT6, NTT9, and ST10 are not phylogenetically close to other Shewanella strains (Fig. 3). Since SXT/R391 ICE elements were also reported in S. upenei (37), Shewanella halifaxensis (21), S. putrefaciens (38), and Shewanella haliotis (39), it is likely that Shewanella species are the reservoirs of SXT/R391 ICEs.
Phylogenetic analysis of the isolated and representative Shewanella strains. The phylogenetic tree was constructed based on the SNPs in the core genes of the Shewanella strains. Strains carrying ICEs in this study are highlighted in red, and other strains carrying ICEs are labeled in blue.
Genomic context and variable regions in SXT/R391 ICEs
ICESalNJT6 was 104,326 bp long, with 47.2% guanine-cytosine (GC) content, while ICESchNTT9 was 112,127 bp in length, with 48.4% GC content. Additionally, ICESalNJT6 and ICESchNTT9 contained 86 and 111 predictive coding genes, respectively. BLASTn analysis showed that ICESalNJT6 exhibited high homologies with unnamed ICEs in Proteus columbae T60 and S. putrefaciens JNE7 with 96.53 and 96.21% identity, respectively, at 72% coverage (Fig. S1). Resistance genes floR, strA, strB, sul2, and insertion sequence ISCR2 were also found in these two ICEs, but tet(X4) was not detected. Meanwhile, ICESchNTT9 exhibited the highest homology with an unnamed ICE in an S. putrefaciens strain JNE7, with 98.25% identity at 80% coverage (Fig. S2). In addition, ICESchST10 carried by strain ST10 was 101,311 bp in length with 48.6% GC content and 96 predictive coding genes. Through BLASTn analysis, we found that ICESchST10 was the most similar to ICEPmiChnRGF134-1 carried by the P. mirabilis strain RGF134-1 with 98.43% identity at 75% coverage (Fig. S3). The novel tmexCD3-toprJ3 gene cluster and resistance genes, sul2, strA, strB, floR, were also found in ICEPmiChnRGF134-1. Notably, ICEPmiChnRGF134-1 also failed to transfer into E. coli. BLASTn against the NCBI database indicated that the tmexCD-toprJ-like gene cluster harbored by ICESchST10 shared 99.83% nucleotide sequence identity with tnfxB2-tmexC2D2.2-toprJ2 reported in Klebsiella oxytoca (40), indicating that ICESchST10 carries the tmexCD2-toprJ2 gene cluster.
The backbones of ICESchNTT9 and ICESchST10 were highly conserved, and both of them exhibited 96% similarity with the core backbone of the reference ICE SXT^MO10^. A total of HS1–5 and VRI–IV have been identified in these ICEs (Fig. 4). Notably, the VRIII in ICESalNJT6 and ICESchST10 contained the same antibiotic resistance genes, including strA, strB, sul2, floR, sul2, and tet(X4). The tet(X4) gene was flanked by ISCR2, forming the ISCR2-aph-tet(X4)-ISCR2 region, indicating that ISCR2 may mediate the insertion of tet(X4) into these two ICEs (Fig. S4). In addition, ICESchST10 carried an MDR region that was integrated into HS4, while the other four HS4 of ICEs did not carry any drug resistance genes. The resistance genes in the HS4 of ICESchST10 contained tet(A), dfrA1, aacA4, aadA2, blaOXA-1, and czcD genes. Through BLASTn analysis, we found that a similar MDR region (query coverage >90%) was widely distributed in P. mirabilis strains. These resistance genes in the HS4 of ICESchST10 were flanked by two copies of ISPpu12, indicating that ISPpu12 may mediate the insertion of this MDR region into ICESchST10. Further analysis showed that a region of 17,365 bp containing tmexCD2-toprJ2 was inserted into the VRIII of ICESchST10 (Fig. 4). Although the conjugation assay of ICESchST10 failed under test conditions, the circular formation of ICESchST10 was confirmed using two pairs of primers P1P2 and P3P4 listed in Table S4, indicating that the circular form of ICESchST10 carrying tmexC2D2-toprJ2 could be excised from the chromosome as the archetypical transfer process of ICE (Fig. S7).
Genomic composition of the ICEs identified in this study. The middle line shows the backbone of the SXT/R391 ICEs with conserved core genes, and the black arrows represent the core genes used for the construction of the phylogenetic tree. The red and yellow arrows represent the drug resistance genes and insertion sequences, respectively. HS1–5 represent five hotspots, and VRI–IV represent four variable regions.
Notably, blaNDM-1 was identified in strain ST10. We found that the blaNDM-1 gene was located in a genomic island, which was designated as SGI-ScNDM-1. SGI-ScNDM-1 was 87,737 bp in length with 52.7% GC content and 91 predictive coding genes. BLASTn analysis showed that the backbone of SGI-ScNDM-1 had high similarities with SGI1 members of other bacterial species. However, the variable region of SGI-ScNDM-1 was different from the known sequences in the NCBI database and contained tet(A), sul1, mph(A), and blaNDM-1 (Fig. S5). Several insertion sequences were also found in this variable region, such as IS5075, ISKpn26, and IS1326. However, the functions of other genes encoded in this region were unknown.
Conclusion
In conclusion, this is the first study to report on the emergence of novel SXT/R391 ICEs carrying tet(X4) and tmexCD2-toprJ2 genes in Shewanella species of seafood origin. The identified SXT/R391 ICEs could mediate the dissemination of tigecycline resistance genes among aquatic bacteria. Tigecycline-resistant bacterial species are prevalent in retail seafood, and Shewanella spp. are likely the potential reservoirs of tigecycline-resistance genes in aquatic products. Contact between seafood and consumers may lead to the dissemination of tigecycline-resistance genes in humans. Therefore, it is imperative to increase our awareness of the emergence of tigecycline-resistant aquatic bacteria.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Peterson LR. 2008. A review of tigecycline--the first glycylcycline. Int J Antimicrob Agents 32 Suppl 4:S 215–S 222. doi:10.1016/S 0924-8579(09)70005-619134522 · doi ↗ · pubmed ↗
- 2Dong N, Zeng Y, Cai C, Sun C, Lu J, Liu C, Zhou H, Sun Q, Shu L, Wang H, Wang Y, Wang S, Wu C, Chan EW-C, Chen G, Shen Z, Chen S, Zhang R. 2022. Prevalence, transmission, and molecular epidemiology of tet(X)-positive bacteria among humans, animals, and environmental niches in China: an epidemiological, and genomic-based study. Sci Total Environ 818:151767. doi:10.1016/j.scitotenv.2021.15176734801490 · doi ↗ · pubmed ↗
- 3Li Y, Sun X, Xiao X, Wang Z, Li R. 2023. Global distribution and genomic characteristics of tet(X)-positive Escherichia coli among humans, animals, and the environment. Sci Total Environ 887:164148. doi:10.1016/j.scitotenv.2023.16414837187393 · doi ↗ · pubmed ↗
- 4Dong N., Zeng Y, Wang Y, Liu C, Lu J, Cai C, Liu X, Chen Y, Wu Y, Fang Y, Fu Y, Hu Y, Zhou H, Cai J, Hu F, Wang S, Wang Y, Wu Y, Chen G, Shen Z, Chen S, Zhang R. 2022. Distribution and spread of the mobilised RND efflux pump gene cluster tmex CD-topr J in clinical Gram-negative bacteria: a molecular epidemiological study. Lancet Microbe 3:e 846–e 856. doi:10.1016/S 2666-5247(22)00221-X 36202114 · doi ↗ · pubmed ↗
- 5Mohsin M, Hassan B, Martins WMBS, Li R, Abdullah S, Sands K, Walsh TR. 2021. Emergence of plasmid-mediated tigecycline resistance tet(X 4) gene in Escherichia coli isolated from poultry, food and the environment in South Asia. Sci Total Environ 787:147613. doi:10.1016/j.scitotenv.2021.14761333992939 · doi ↗ · pubmed ↗
- 6Liu YY, Lu L, Yue C, Gao X, Chen J, Gao G, Li K, Deng H, Liu JH. 2024. Emergence of plasmid-mediated high-level tigecycline resistance gene tet(X 4) in Enterobacterales from retail aquatic products. Food Res Int 178:113952. doi:10.1016/j.foodres.2024.11395238309872 · doi ↗ · pubmed ↗
- 7Crona B, Wassénius E, Troell M, Barclay K, Mallory T, Fabinyi M, Zhang W, Lam VWY, Cao L, Henriksson PJG, Eriksson H. 2020. China at a crossroads: an analysis of China's changing seafood production and consumption. One Earth 3:32–44. doi:10.1016/j.oneear.2020.06.013 · doi ↗
- 8Yousfi K, Bekal S, Usongo V, Touati A. 2017. Current trends of human infections and antibiotic resistance of the genus Shewanella. Eur J Clin Microbiol Infect Dis 36:1353–1362. doi:10.1007/s 10096-017-2962-328299457 · doi ↗ · pubmed ↗