Discovery of the first resinicolous fungus in Mycosphaerellales (Dothideomycetes): Resinomelaniacommunis from conifer resins in Poland

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
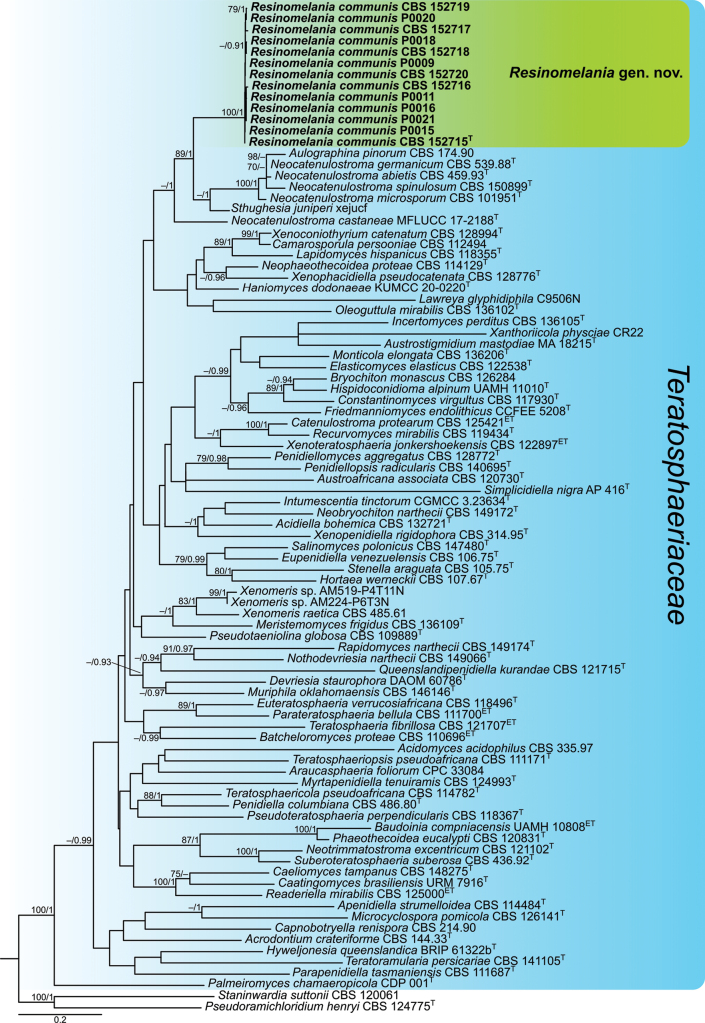 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3| Species | Strain | Isolation source | Location | GenBank accession numbers | ||||
|---|---|---|---|---|---|---|---|---|
|
|
|
|
|
| ||||
|
| CBS 132721T | Highly acidic soil | Czech Republic |
| – |
| – | – |
|
| CBS 335.97 | Acidophilic algae | Germany |
|
| – | – | – |
|
| CBS 144.33T | – | Netherlands |
|
| – | – | – |
|
| CBS 114484T | Russia |
|
|
| – | – | |
|
| CPC 33084 |
| Chile |
|
| – | – | – |
|
| CBS 174.90 | France |
| – |
| – | – | |
|
| CBS 120730T = CPC 13119 |
| Australia |
|
|
|
|
|
|
| MA 18215T |
| Antarctica |
| – | – | – | – |
|
| CBS 110696ET |
| South Africa |
|
|
| – | – |
|
| UAMH 10808ET | Tree branches near a distillery | France |
|
| – |
|
|
|
| CBS 126284 | Bryophyte | Finland |
|
| – | – | – |
|
| URM 7916T |
| Brazil |
|
| – |
| – |
|
| CBS 148275T = CPC 41060 | Office dust | USA |
|
| – |
|
|
|
| CBS 112494 = CPC 3350 | Australia |
|
|
| – | – | |
|
| CBS 214.90 |
| Japan |
|
| – | – | – |
|
| CBS 125421ET = CPC 15370 |
| South Africa |
|
|
| – | – |
|
| CBS 117930T = TRN79 | Rock, limestone | Spain |
|
|
| – |
|
|
| DAOM 60786T | United Kingdom | – |
| – | – | – | |
|
| CBS 122538T = CCFEE 5313 | Lichen thallus ( | Antarctica |
|
| – | – | – |
|
| CBS 106.75T | Venezuela |
|
|
|
|
| |
|
| CBS 118496T = CPC 11167 | Indonesia | – |
| – | – | – | |
|
| CCFEE 5208T | Rock | Antarctica | – |
| – | – | – |
|
| KUMCC 20-0220T | China |
|
|
|
| – | |
|
| UAMH 11010T |
| Canada |
|
| – | – | – |
|
| CBS 107.67T | Portugal |
|
| – |
| – | |
|
| BRIP 61322bT | Cocoon of an unidentified microlepidoptera | Australia |
|
| – | – | – |
|
| CBS 136105T = CCFEE 5385 | Rock | Italy |
|
|
| – |
|
|
| CGMCC 3.23634T |
| China |
|
|
|
| – |
|
| CBS 118355T = TRN500 | Rock | Spain |
|
| – | – |
|
|
| C9506N |
| USA |
| – | – | – | – |
|
| CBS 136109T = CCFEE 5508 | Rock | Argentina |
|
|
| – |
|
|
| CBS 126141T = CPC 16175 | Germany |
|
| – | – | – | |
|
| CBS 136206T = CCFEE 5394 | Rock | Italy |
|
|
| – |
|
|
| CBS 146146T = CCF 5751 | Outside wall of alcohol distillery | USA |
|
| – | – | – |
|
| CBS 124993T = CPC 13692 |
| Australia |
|
|
|
|
|
|
| CBS 149172T = CPC 41972 | Netherlands |
|
| – | – | – | |
|
| CBS 459.93T | Germany |
|
| – | – | – | |
|
| MFLUCC 17–2188T | Italy |
|
| – | – | – | |
|
| CBS 539.88T | Rock | Germany |
|
|
| – | – |
|
| CBS 101951T = CPC 1960 | South Africa |
|
|
|
|
| |
|
| CBS 150899T = FMR 18793 | PVC pipe | Spain |
|
| – | – | – |
|
| CBS 114129T = CPC 2831 |
| South Africa |
|
|
| – | – |
|
| CBS 121102T = CPC 13092 |
| Australia |
|
|
|
|
|
|
| CBS 149066T = CPC 42166 | Netherlads |
|
| – |
|
| |
|
| CBS 136102T = CCFEE 5523 | Rock | Antarctica |
|
| – | – | – |
|
| CDP 001T | Portugal |
|
| – | – | – | |
|
| CBS 111687T = CMW 14780 = CPC 1555 |
| Australia |
|
|
|
|
|
|
| CBS 111700ET = CPC 1821 |
| South Africa |
|
| – | – | – |
|
| CBS 486.80T |
| Colombia |
|
|
|
|
|
|
| CBS 128772T = CPC 18278 | South Africa |
|
| – | – | – | |
|
| CBS 140695T = UTHSC DI-13-256 | USA |
|
| – | – | – | |
|
| CBS 120831T = CPC 12918 |
| Australia |
|
|
|
|
|
|
| CBS 124775T = CPC 13121 |
| Australia |
|
|
|
|
|
|
| CBS 109889T | Rock, sandstone | Italy |
|
|
| – |
|
|
| CBS 118367T = CPC 10983 |
| Colombia |
|
|
|
|
|
|
| CBS 121715T = CPC 13333 | Unidentified tree, exudates of bleeding stem cankers | Australia |
|
|
|
|
|
|
| CBS 149174T = CPC 41974 | Netherlands |
|
| – | – | – | |
|
| CBS 125000ET = CPC 12370 |
| Australia |
|
|
|
|
|
|
| CBS 119434T = CCFEE 5264 | Rock, sandstone | Antarctica |
|
|
| – | – |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| – |
|
|
|
|
|
|
|
|
| – |
|
|
|
|
|
|
|
|
| – |
|
|
|
|
|
|
|
|
| – |
|
|
|
|
|
|
|
|
| – |
|
|
|
|
|
|
|
|
| – |
|
|
|
| CBS 147480T | Brine | Poland |
|
|
| – | – |
|
| AP 416T = CBMAI 1939 |
| Brazil |
|
| – |
|
|
|
| CBS 120061 |
| Australia |
|
|
|
|
|
|
| CBS 105.75T = ATCC 24788 = FMC 245 | Venezuela |
|
| – | – | – | |
|
| xejucf |
| USA |
|
| – | – | – |
|
| CBS 436.92T = CPC 515 |
| Brazil |
|
|
|
| – |
|
| CBS 141105T = CPC 11410 |
| South Korea |
|
|
|
| – |
|
| CBS 121707ET = CPC 13960 | South Africa |
|
| – |
|
| |
|
| CBS 114782T = CPC 1230 |
| Zambia |
|
|
|
|
|
|
| CBS 111171T = CPC 1261 | South Africa |
|
|
|
|
| |
|
| CR22 |
| United Kingdom |
| – | – | – | – |
|
| CBS 128994T = CMW 22113 | Twig litter of | South Africa |
|
| – | – | – |
|
| CBS 485.61 | Switzerland |
|
| – | – | – | |
| AM224-P6T3N | Pinyon tree, needle | USA | – |
| – | – | – | |
| AM519-P4T11N | Pinyon tree, needle | USA | – |
| – | – | – | |
|
| CBS 314.95T | Leaf litter of | Cuba |
|
|
| – | – |
|
| CBS 128776T = CPC 18472 |
| South Africa |
|
|
| – | – |
|
| CBS 122897ET = CPC 13984 | South Africa |
|
|
| – | – | |
| Partitions |
| MB |
|---|---|---|
|
| TrN+I+G4 | GTR+I+G4 |
|
| TIM1+I+G4 | SYM+I+G4 |
|
| TIM3ef+I+G4 | SYM+I+G4 |
|
| TPM2uf+G4 | HKY+G4 |
|
| TPM2+I+G4 | SYM+I+G4 |
|
| HKY+I+G4 | HKY+I+G4 |
|
| TrNef+I+G4 | JC+I+G4 |
|
| JC+I | JC+I |
|
| TrN+G4 | HKY+G4 |
|
| TPM3uf+G4 | HKY+G4 |
|
| JC+I | JC+I |
|
| TPM1uf+G4 | HKY+G4 |
|
| TPM3uf+G4 | K80+G4 |
|
| HKY+I+G4 | HKY+I+G4 |
- —Narodowe Centrum Nauki 501100004281 https://ror.org/03ha2q922 http://doi.org/10.13039/501100004281
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Mycorrhizal Fungi and Plant Interactions · Yeasts and Rust Fungi Studies
Introduction
Resins play a wide protective role in plant defense, as they constitute both a physical and chemical barrier against harmful organisms. Due to their physical properties, resins coat wounds and engulf potential threats. In turn, their chemical composition protects plants from pests, herbivores, and pathogenic microorganisms (Langenheim 1994, 2003; Cabrita 2018). This chemical protection is made possible by their composition. In general, resins are a mixture of volatile and nonvolatile terpenoid and/or phenolic compounds. Gymnosperms (conifers) produce only terpenoid resins, whereas angiosperms produce both terpenoid and phenolic resins (Langenheim 1994, 2003; Cabrita 2018; Seyfullah et al. 2018). Such a composition makes resins recalcitrant to microbial decomposition and grants them antimicrobial properties (Langenheim 2003). Despite this, some fungi can live on such substrates. These are referred to as resinicolous fungi (Rikkinen and Poinar 2000; Tuovila et al. 2013; Beimforde et al. 2017a, b, 2020; Mitchell 2021).
Resinicolous fungi have evolved across different lineages within the fungal kingdom. They are best known within Eurotiomycetes (Chaetothyriales, Mycocaliciales), Leotiomycetes (Helotiales, Leotiales), Orbiliomycetes (Orbiliales), and Xylonomycetes (syn. Sareomycetes) (Xylonales, syn. Sareales). However, some species also occur in various scattered phylogenetic lineages (Rikkinen and Poinar 2000; Tuovila et al. 2013; Beimforde et al. 2017a,b, 2020; Baral et al. 2020; Hashimoto et al. 2021; Mitchell 2021; Mitchell et al. 2021; Czachura and Janik 2025). Resinicolous fungi are uncommon in the class Dothideomycetes. Within this class, only scattered reports of fungi exhibiting a resinicolous lifestyle exist, and their identification has mostly relied on morphological characters. Thus, their exact phylogenetic placement remains unknown (Mitchell 2021). The best-known examples—supported by DNA sequence data and phylogenetic analyses—are members of the order Mytilinidiales (Mitchell 2021; Czachura and Janik 2024). However, some reports based on morphologically identified species (Hughes 1968; Mitchell 2021), metabarcoding or recently published studies (Gøtzsche et al. 2024; Czachura et al. 2025) suggest that resinicolous fungi may be more common in Dothideomycetes than currently recognized.
During a study of fungi inhabiting resin of four conifer species in Poland (Abiesalba, Larixdeciduassp.polonica, Piceaabies, and Pinussylvestris; all from the family Pinaceae), 13 strains of a dark, slow-growing fungus belonging to the class Dothideomycetes were isolated. This was one of the most commonly isolated fungi from the resins of all studied conifer trees. Initial identification based on ITS rDNA sequences indicated an affinity with members of the order Mycosphaerellales. Based on multi-locus phylogenetic analyses and morphological features, the isolated strains were identified as representing an undescribed species in a new genus within the family Teratosphaeriaceae.
Materials and methods
Fungal isolation
Resin samples were collected from three different areas in central and southern Poland: the Modrzyna Reserve in the Beskid Niski Mountains, the Świętokrzyski National Park, and the Tatra National Park. Resin outpourings were sterilely collected from four conifer species—Abiesalba, Larixdeciduassp.polonica, Piceaabies, and Pinussylvestris (Pinaceae). In the laboratory, to remove external contamination, samples were washed with a 0.1% solution of Tween 20 and physiological saline. Details of the washing procedure are described in Czachura and Janik (2025). After drying, the outer resin layer was scraped using razor blades. Resin particles were spread on Petri dishes with four different media: dichloran–18% glycerol agar (DG18), dichloran rose bengal chloramphenicol agar (DRBC), rose bengal chloramphenicol agar (RBC), and potato dextrose agar (PDA). The mycological media were prepared as described by Crous et al. (2019) and Corry et al. (1995). Cultures containing resin particles were incubated in the dark at 15 °C or 25 °C. Selected representatives of each morphotype were transferred to malt extract agar (MEA) for storage prior to further processing. Thirteen strains of a dark, slow-growing fungus from the class Dothideomycetes were analyzed in detail in this study. All examined strains of Resinomelaniacommunis are preserved as dried cultures (KRAM F) and cultures stored under paraffin oil in the fungal collection of the W. Szafer Institute of Botany, Polish Academy of Sciences, Kraków, Poland. Representative cultures are also deposited in the culture collection (CBS) of the Westerdijk Fungal Biodiversity Institute, Utrecht, the Netherlands.
Morphological analyses
Macroscopic features of the cultures were examined on 90 mm Petri dishes with four different media: malt extract agar (MEA), oatmeal agar (OA), potato dextrose agar (PDA), and synthetic nutrient-poor agar (SNA) after four weeks at 15 °C and 25 °C in darkness. Micromorphological features were studied on SNA. Micromorphology was observed on slides using a Nikon Eclipse 80i light microscope. Photographs were taken with the NIS‐Elements BR 3.0 imaging software. The figures showing morphological features of the analyzed strain were prepared using Inkscape 0.92.4.
DNA extraction, amplification, and sequencing
Extraction of DNA was performed using the CTAB method as described in Czachura and Janik (2025). For phylogenetic analyses of isolated strains, five loci were amplified: the internal transcribed spacer 1 and 2 regions and intervening 5.8S rRNA gene (ITS), the partial 28S rRNA gene (LSU), the partial RNA polymerase II second largest subunit gene (rpb2), the partial translation elongation factor 1-α gene (tef1), and the partial β-tubulin gene (tub2). A polymerase chain reaction (PCR) mixture for each sample contained 17.25 μL cell culture grade water (Sigma Aldrich, USA), 2.5 μL of 10× PCR Buffer (Sigma Aldrich, USA), 2.5 μL of 25 mM MgCl_2_ (Sigma Aldrich, USA), 0.5 μL of 10 mM dNTPs (EURx, Poland), 0.5 μL of each 10 μM primer (Sigma Aldrich, USA), 0.25 μL of 5 U/μL Taq DNA Polymerase (Sigma Aldrich, USA), and 1 μL of isolated DNA. A fragment containing ITS and LSU was amplified using primers ITS1 and LR5 (White et al. 1990; Vilgalys and Hester 1990), whereas parts of protein-coding genes were amplified using primers fRPB2-5F and fRPB2-7cR for rpb2 (Liu et al. 1999), EF1-728F and EF-2 for tef1 (O’Donnell et al. 1998; Carbone and Kohn 1999), and T1 and β-Sandy-R for tub2 (O’Donnell and Cigelnik 1997; Stukenbrock et al. 2012). The amplification of the fragment containing ITS and LSU was set as follows: an initial denaturation at 94 °C for 3 min, followed by 35 cycles of amplification (denaturation at 94 °C for 45 s, annealing at 50 °C for 45 s, and elongation at 72 °C for 2 min), and a final elongation step at 72 °C for 10 min. The amplification of rpb2 was carried out under the following conditions: an initial denaturation at 94 °C for 3 min, followed by 35 cycles of amplification (denaturation at 94 °C for 60 s, annealing at 54 °C for 90 s, and elongation at 72 °C for 2 min), and a final elongation step at 72 °C for 10 min. Amplification conditions for tef1 were conducted in the following steps: an initial denaturation at 94 °C for 3 min, followed by 40 cycles of amplification (denaturation at 94 °C for 45 s; annealing at 52 °C for 30 s; elongation at 72 °C for 90 s), and a final elongation step at 72 °C for 6 min. Amplification conditions for tub2 were as follows: an initial denaturation at 94 °C for 3 min, followed by 40 cycles of amplification (denaturation at 94 °C for 45 s; annealing at 52 °C for 30 s; elongation at 72 °C for 90 s), and a final elongation step at 72 °C for 7 min. PCR products were visualized by agarose gel electrophoresis. Amplicons were enzymatically cleaned using an Exo-BAP Mix (EURx, Poland) according to the manufacturer’s instructions and submitted for bidirectional sequencing by Macrogen Europe B.V. (Amsterdam, The Netherlands). The ITS and LSU fragments were sequenced using primers ITS1 and ITS4 for ITS (White et al. 1990) and LSU1Fd and LR5 for LSU (Vilgalys and Hester 1990; Crous et al. 2009) to ensure high-quality sequences across the full amplicon lengths. Protein-coding loci were sequenced using the same primer pairs as those used for amplification.
Phylogenetic analyses
Sequence assembly and preparation of sequence alignments were carried out using Geneious Prime 2022.1.1. Newly generated forward and reverse sequences were assembled and trimmed. To determine the closest taxonomic affinities of the obtained sequences, a megablast search was performed in the NCBI GenBank nucleotide database (https://www.ncbi.nlm.nih.gov/genbank/) (Zhang et al. 2000). Phylogenetic analyses were conducted using combined LSU, ITS, rpb2, tef1, and tub2 sequences of representatives of the Teratosphaeriaceae. Sequences used in the phylogenetic analyses are listed in Table 1. Reference sequences were derived from ex-type strains (whenever possible) of the type species of all genera assigned to Teratosphaeriaceae (Hyde et al. 2024). Additionally, sequences of all Xenomeris strains available in GenBank were included in the analyses, as Xenomerisjuniperi (current name: Sthughesiajuniperi) was identified as one of the closest relatives of the analyzed strains in the megablast search. Each single-locus dataset was aligned separately using the MAFFT algorithm (Katoh et al. 2019). Maximum likelihood (ML) and Bayesian inference (BI) analyses were used to estimate phylogenetic relationships. ML analysis was conducted using RAxML-NG v. 1.2.2 (Kozlov et al. 2019), with 1,000 bootstrap replicates, and BI analysis was performed using MrBayes v. 3.2.6 (Ronquist et al. 2012). The most suitable substitution models for each locus were selected separately using ModelTest-NG v. 0.2.0 (Darriba et al. 2020), based on the Bayesian Information Criterion (BIC). Substitution models applied in ML and BI analyses are provided in Table 2. For the BI analysis, four simultaneous Markov Chain Monte Carlo (MCMC) chains were run for 50,000,000 generations with a sampling frequency of 100 generations. The first 25% of sampled trees were discarded as burn-in, and the remaining trees were used to reconstruct the consensus tree and calculate posterior probability values for branches. The ML and BI phylogenetic trees were visualized using FigTree v. 1.4.3. The final tree topology was based on the ML analysis, with maximum likelihood bootstrap (MLB) and Bayesian posterior probability (BPP) values indicated next to branches. The final phylogenetic tree was graphically edited in Inkscape 0.92.4.
Results
Phylogenetic analyses
The phylogenetic placement of the analyzed strains was determined using multi-locus phylogenetic analyses based on the combined LSU, ITS, rpb2, tef1, and tub2 dataset. ML and BI analyses yielded similar tree topologies. The tree obtained from the ML analysis is presented in Fig. 1. Maximum likelihood bootstrap (MLB) support values above 70% and Bayesian posterior probabilities (BPP) above 0.9 were considered well supported, and only those values are shown next to the branches.
Maximum likelihood consensus tree inferred from the combined LSU, ITS, rpb2, tef1, and tub2 multiple sequence alignment of representatives of the family Teratosphaeriaceae. The position of Resinomelaniacommunis is indicated in bold and marked by a colored block. Maximum likelihood bootstrap (MLB) support values ≥70% and Bayesian posterior probabilities (BPP) ≥0.9 are shown next to the branches (MLB/BPP). Ex-type and ex-epitype strains are indicated with T and ET, respectively. The scale bar represents the average number of substitutions per site.
The phylogenetic analyses inferred that all genera in the family Teratosphaeriaceae formed a well-supported clade (MLB/BPP = 100/1), although most internal branches were weakly supported. As a result, the relationships among genera within Teratosphaeriaceae were largely unresolved, with some exceptions, including the group analyzed in this study. Sequences of Xenomeris species formed two distinct lineages: one composed of X.raetica and Xenomeris sp., which was related to Meristemomycesfrigidus (MLB/BPP = 45/1); and another containing Xenomerisjuniperi, which was sister to Neocatenulostroma (MLB/BPP = 63/1). The latter species is also known under the combination Sthughesiajuniperi, the type species of the genus Sthughesia. Sequences of the analyzed strains from resin samples formed a fully supported, independent lineage (MLB/BPP = 100/1) that clustered as a sister group to most members of the genera Neocatenulostroma and Sthughesia, both of which also formed independent lineages. The only exception was Neocatenulostromacastaneae, which formed a distinct single-species lineage distant from other Neocatenulostroma species, including the type species, N.microsporum. The phylogenetic distance from Neocatenulostroma and Sthughesia species suggests that the strains analyzed from conifer resins represent an undescribed species belonging to a novel genus within the family Teratosphaeriaceae.
Taxonomy
Resinomelania
Taxon classificationFungiMycosphaerellalesTeratosphaeriaceae
Czachura & Piątek gen. nov.
A6BB7CF4-811D-5FB4-9F14-662D4841D21A
859781
Etymology.
The genus name refers to a resinicolous lifestyle and the dark, melanized mycelium of this fungus.
Description.
Mycelium composed of branched, septate, hyaline, subhyaline, pale brown to brown, smooth or verruculose, cylindrical and moniliform hyphae, with hyphal swellings. Chlamydospores subglobose or broadly ellipsoidal, pale brown to brown, smooth or slightly verruculose, aseptate or muriformly septate, intercalary or terminal, single. Arthroconidia broadly ellipsoidal, one-septate or rarely aseptate, mostly constricted at septa, pale brown to brown, smooth or verruculose, intercalary or terminal, in chains.
Type species.
Resinomelaniacommunis Czachura & Piątek.
Resinomelania
communis
Taxon classificationFungiMycosphaerellalesTeratosphaeriaceae
Czachura & Piątek sp. nov.
7331A166-5292-5BE3-98E5-2000BB47AA74
859782
Etymology.
The epithet is derived from the Latin word that means ‘‘common’’ and refers to the common occurrence of this species on conifer resins.
Morphology of cultures of Resinomelaniacommunis (CBS 152715) after 4 weeks at 25°C: A, B. Colony on MEA; C, D. Colony on OA; E, F. Colony on PDA; G, H. Colony on SNA.
Typus.
Poland • Świętokrzyskie Province, Kielce County, the Świętokrzyski National Park, Mt. Łysica, on resin of Piceaabies, 18 June 2021, leg. P. Czachura (holotype: KRAM F-60010; culture ex-type: CBS 152715 = P0008).
Morphology of Resinomelaniacommunis (CBS 152715): A. Cylindrical hyphae; B. Moniliform hyphae; C. Cylindrical and moniliform hyphae; D. Anastomosing hyphae; E–G. Aseptate hyphal swellings; H–K. Hyphal swellings in form of multicellular bodies; L–O. Chlamydospores; P–V. Arthroconidia. Scale bars: 10 μm.
Description.
Mycelium composed of branched, septate, hyaline to subhyaline (when young) or pale brown to brown (when mature), smooth or verruculose, guttulate, sometimes anastomosing, cylindrical (2.0–5.0 μm wide) and moniliform hyphae (3.5–9.5 μm wide). Hyphae with numerous aseptate, one-septate, or multiseptate hyphal swellings, 5.0–9.5 μm wide; multiseptate hyphal swellings in form of multicellular bodies, which are formed by meristematic development of hyphae. Chlamydospores subglobose or broadly ellipsoidal, pale brown to brown, smooth or slightly verruculose, aseptate or muriformly septate, 6.5–12.5 × 6.0–10.0 μm, intercalary or terminal, single, guttulate. Arthroconidia formed by arthric disarticulation, broadly ellipsoidal, one-septate or rarely aseptate, mostly constricted at septa, pale brown to brown, smooth or verruculose, 6.0–12.5 × 4.5–7.0 μm, intercalary or terminal, in chains, guttulate.
Culture characteristics.
Colonies on MEA and PDA convex, with folded surface, grey olivaceous to smoke grey, margin slightly lobate to slightly crenate; on MEA reaching 14 mm diam. after 4 weeks at 15 °C and 16 mm diam. after 4 weeks at 25 °C, reverse grayish black; on PDA reaching 12 mm diam. after 4 weeks at 15 °C and 16 mm diam. after 4 weeks at 25 °C, reverse grayish black. Colonies on OA and SNA flat with protrusion at center, fuscous black, margin nearly entire to slightly undulate; on OA reaching 10 mm diam. after 4 weeks at 15 °C and 12 mm diam. after 4 weeks at 25 °C, reverse grayish black; on SNA reaching 6 mm diam. after 4 weeks at 15 °C and 12 mm diam. after 4 weeks at 25 °C, reverse grayish black.
Additional specimens examined.
Poland • Małopolskie Province, Tatra County, the Tatra National Park, Dolina Strążyska, on resin of Piceaabies, 23 Sept. 2020, leg. P. Czachura & M. Piątek (KRAM F-60012; culture: CBS 152716 = P0010); • Podkarpackie Province, Krosno County, the Modrzyna Reserve, on resin of Pinussylvestris, 22 Oct. 2020, leg. P. Czachura (KRAM F-60014; culture: CBS 152717 = P0012); • Świętokrzyskie Province, Kielce County, the Świętokrzyski National Park, the strict protection area Mokry Bór, on resin of Pinussylvestris, 18 June 2021, leg. P. Czachura (KRAM F-60016; culture: CBS 152718 = P0014); • Świętokrzyskie Province, Kielce County, the Świętokrzyski National Park, Psarska Góra, on resin of Abiesalba, 15 Oct. 2020, leg. P. Czachura (KRAM F-60019; culture: CBS 152719 = P0017); • Podkarpackie Province, Krosno County, the Modrzyna Reserve, on resin of Larixdeciduassp.polonica, 22 June 2021, leg. P. Czachura (KRAM F-60021; culture: CBS 152720 = P0019); • Świętokrzyskie Province, Kielce County, the Świętokrzyski National Park, the strict protection area Psarski Dół, on resin of Piceaabies, 16 Oct. 2020, leg. P. Czachura (KRAM F-60011; culture: P0009); • Małopolskie Province, Tatra County, the Tatra National Park, Dolina Filipka, on resin of Piceaabies, 15 July 2021, leg. P. Czachura & M. Piątek (KRAM F-60013; culture: P0011); • Świętokrzyskie Province, Kielce County, the Świętokrzyski National Park, Dolina Czarnej Wody, on resin of Pinussylvestris, 15 Oct. 2020, leg. P. Czachura (KRAM F-60017; culture: P0015); • Podkarpackie Province, Krosno County, the Modrzyna Reserve, on resin of Abiesalba, 21 June 2021, leg. P. Czachura (KRAM F-60018; culture: P0016); • Małopolskie Province, Tatra County, the Tatra National Park, Łysa Skałka, on resin of Abiesalba, 08 Oct. 2020, leg. P. Czachura & M. Piątek (KRAM F-60020; culture: P0018); • Świętokrzyskie Province, Kielce County, the Świętokrzyski National Park, Chełmowa Góra, on resin of Larixdeciduassp.polonica, 16 Oct. 2020, leg. P. Czachura (KRAM F-60022; culture: P0020); • Świętokrzyskie Province, Kielce County, the Świętokrzyski National Park, Chełmowa Góra, on resin of Larixdeciduassp.polonica, 19 June 2021, leg. P. Czachura (KRAM F-60023; culture: P0021).
Notes.
The genus Resinomelania forms a distinct phylogenetic lineage close to the genera Neocatenulostroma and Sthughesia. Resinomelania differs from Neocatenulostroma by having one-septate or aseptate arthroconidia formed by disarticulation of hyphae. Moreover, Resinomelania frequently forms hyphal swellings and chlamydospores. In contrast, members of the genus Neocatenulostroma produce variously shaped, multiseptate conidia (= arthroconidia) with transverse, longitudinal, or oblique septa that arise from conidiophores, which may be aggregated into sporodochia or are formed by disarticulation of hyphae. Additionally, conidia in Neocatenulostroma are characterized by having rounded, pale cells at both sides (apex and base), while this feature is absent in Resinomelania (Butin et al. 1996; Ruibal et al. 2005; Crous et al. 2007a; Quaedvlieg et al. 2014). The morphological comparison of Resinomelania with Sthughesia is not possible since Sthughesiajuniperi is described from sexual morph (ascomata with asci and ascospores), although with information that hyphae are moniliform (Barr 1987a). Additionally, phialides were observed in this fungus (Barr 1987b). The morphology of Resinomelaniacommunis, with hyphae that develop into arthroconidia and chlamydospores, resembles the morphology of different genera and species of so-called “black yeasts”, which are phylogenetically distinct (Isola et al. 2016; Humphries et al. 2017; Trovão et al. 2019; Muggia et al. 2021; Piątek et al. 2024).
Discussion
In this study, 13 strains of a dark, slow-growing fungus belonging to the class Dothideomycetes were isolated from resin outpourings of four conifer species (Abiesalba, Larixdeciduassp.polonica, Piceaabies, and Pinussylvestris) in Poland. Based on multi-locus sequence analyses, the strains were identified and described as a new species, Resinomelaniacommunis, accommodated in the newly established genus Resinomelania, which resides in the family Teratosphaeriaceae within the order Mycosphaerellales. Teratosphaeriaceae is the second-largest family within Mycosphaerellales, following the most speciose family, Mycosphaerellaceae (Hyde et al. 2024; Pem et al. 2024). Members of Teratosphaeriaceae exhibit diverse lifestyles; they are known as plant pathogens, saprobes, human pathogens, or animal inhabitants (Quaedvlieg et al. 2014; Duarte et al. 2017). Some species also thrive in extreme environments, including rocks, honeydew deposits, acidic or saline soils and waters, or near alcohol distilleries (Hujslová et al. 2013; Egidi et al. 2014; Scott et al. 2016; Czachura et al. 2021; Kolařík et al. 2021; Crous et al. 2023). Resinomelaniacommunis, as a fungus inhabiting resin outpourings of conifers—substrates rich in terpenoids and characterized by low water availability—may undoubtedly be considered an extremophilic species. It is, however, the first resinicolous fungus reported in both the family Teratosphaeriaceae and the order Mycosphaerellales. In this context, it is worth noting that another member of this family, Queenslandipenidiellakurandae (syn. Penidiellakurandae), was described from exudates of unidentified trees in Australia caused by fungal infection (Crous et al. 2007b; Quaedvlieg et al. 2014). However, it remains unclear whether these exudates were resin outpourings; presumably not, though further investigation is needed. Phylogenetically, Resinomelaniacommunis is most closely related to members of the genera Neocatenulostroma and Sthughesia.
The genus Neocatenulostroma comprises six species names, including one based on a validly described species (Aulographinapinorum), although its combination into Neocatenulostroma as N.pinorum is invalid [Art. F.5.1, ICNafp (Shenzhen)] (Butin et al. 1996; Taylor and Crous 2000; Crous et al. 2007a; Quaedvlieg et al. 2014; Phukhamsakda et al. 2022; Sastoque et al. 2024). It should be noted that Aulographinapinorum is the type species of the genus Aulographina, an older name that would have priority over the younger name Neocatenulostroma. The strains CBS 174.90 and CBS 302.71 of Aulographinapinorum used in previous phylogenetic analyses (Crous et al. 2009; Schoch et al. 2009; Sastoque et al. 2024) do not originate from the type specimen, and Cheewangkoon et al. (2012) considered it unlikely that they are authentic representatives of that species. Therefore, the taxonomic status of Aulographinapinorum/Neocatenulostromapinorum and the genera Aulographina/Neocatenulostroma requires further investigation using new cultures clearly linked to A.pinorum. Based on the phylogenetic analyses conducted in this study, only four species belong to Neocatenulostroma s. str. (Fig. 1). Neocatenulostromacastaneae clusters outside the strongly supported clade comprising the genera Resinomelania, Neocatenulostroma s. str., and Sthughesia. This placement is consistent with the analyses conducted by Phukhamsakda et al. (2022), where N.castaneae was described and grouped—albeit with weak support—on a long branch adjacent to Neocatenulostroma species. However, Sthughesiajuniperi was not included in their analyses. Neocatenulostromacastaneae should be accommodated in a distinct genus within the family Teratosphaeriaceae, a conclusion supported by its phylogenetic position and certain morphological features observed in culture, particularly the production of appressorial pegs, chlamydospores, and chlamydospore-like structures—traits not reported in species of Neocatenulostroma s. str. (Butin et al. 1996; Taylor and Crous 2000; Crous et al. 2007a; Quaedvlieg et al. 2014; Markovskaja et al. 2016; Phukhamsakda et al. 2022; Sastoque et al. 2024).
The genus Sthughesia includes only one species, S.juniperi, previously known as Dimeriumjuniperi or Xenomerisjuniperi (Dearness 1926; Müller and von Arx 1962; Barr 1987a, 1987b). It was originally described based on morphological characters (Barr 1987a) but later confirmed by molecular data (Winton et al. 2007). However, due to limited sampling of sequences within Teratosphaeriaceae, S.juniperi grouped closely with Xenomerisraetica in the study by Winton et al. (2007), as well as in the study conducted by Zhang et al. (2011). In the present study, sequences from all available strains of Sthughesia and Xenomeris spp. were analyzed in a broader phylogenetic context that included all genera in the family Teratosphaeriaceae (Fig. 1). This was necessary because our initial megablast query indicated that Sthughesiajuniperi (syn. Xenomerisjuniperi) was one of the closest relatives of Resinomelaniacommunis. The phylogenetic analyses clearly separated S.juniperi from the remaining Xenomeris species (Fig. 1). Sthughesia represents an independent generic lineage closely related to Neocatenulostroma s. str. and R.communis. This study reveals its phylogenetic placement for the first time.
A closer look at the ecology of the analyzed clade containing the genera Neocatenulostroma s. str., Resinomelania, and Sthughesia reveals several adaptations. Members of Neocatenulostroma have been reported as epiphytes or pathogens on living needles of Abiesalba and Pinus spp. (Butin et al. 1996; Markovskaja et al. 2016; Lazarević and Menkis 2020), on living leaves of Proteacynaroides (Taylor and Crous 2000; Quaedvlieg et al. 2014), on stones (Ruibal et al. 2005; Crous et al. 2007a; Quaedvlieg et al. 2014), on painted outdoor walls and PVC pipes of industrial warehouses (Quaedvlieg et al. 2014; Sastoque et al. 2024), as well as on skin lesions of human patients (Butin et al. 1996). Sthughesiajuniperi is a plant-associated species, isolated from a living branch of Juniperusoccidentalis (Dearness 1926). Resinomelaniacommunis is also a plant-associated fungus, but its resinicolous lifestyle distinguishes it from closely related genera and species. Its common occurrence on the resins of four different conifer species in the family Pinaceae indicates that it is a generalist in terms of host preference but a resinicolous species in terms of ecological strategy. Interestingly, only two ITS sequences available in GenBank were similar to those of R.communis. The first sequence, obtained from the bark beetle Hylesinuscrenatus on Fraxinusexcelsior in the Czech Republic (LR961841; named as Neocatenulostromamicrosporum; identities: 450/451, no gaps; M. Kolařík, unpubl.), differs from R.communis at only one nucleotide position, suggesting conspecificity. The second sequence (MH349092; named as Neocatenulostromamicrosporum; identities: 493/510, including 5 gaps), obtained from a lichen thallus on quartz in the extremely arid Atacama Desert in Peru (Díaz et al. 2023), is more divergent from R.communis, possibly representing a related but distinct species. However, the sequence is nearly identical along its entire length, with differences occurring only at the end of the read. Nevertheless, the widespread occurrence of R.communis on conifer resins and the rarity of related environmental sequences in GenBank from other substrates suggest that R.communis may exhibit a strong preference for resin outpourings.
In summary, this study provides new insights into the diversity of resinicolous fungi and the ecology of the order Mycosphaerellales, particularly the family Teratosphaeriaceae, by introducing the first genus and species exhibiting a resinicolous lifestyle in this fungal lineage. Plant resins represent a unique and chemically complex microhabitat in the natural environment. Fungi inhabiting such extreme and atypical environments often exhibit specialized metabolic pathways and adaptations. Therefore, microorganisms from these habitats hold high potential for numerous biotechnological applications (Tiquia-Arashiro and Grube 2019; Sahay 2022). Continued research on resinicolous fungi will contribute to a better understanding of this intriguing group within the fungal kingdom.
Supplementary Material
XML Treatment for Resinomelania
XML Treatment for Resinomelania communis
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baral HO Weber E Marson G (2020) Monograph of Orbiliomycetes (Ascomycota) Based on Vital Taxonomy. Part I + II.National Museum of Natural History, Luxembourg, 1752 pp.
- 2Barr ME (1987 a) New taxa and combinations in the Loculoascomycetes.Mycotaxon 29: 501–505.
- 3Barr ME (1987 b) Prodromus to Class Loculoascomycetes. Hamilton I. Newell, Inc. Amherst, Massachusetts.
- 4Beimforde C Seyfullah LJ Perrichot V Schmidt K Rikkinen J Schmidt AR (2017 a) Resin exudation and resinicolous communities on Araucariahumboldtensis in New Caledonia.Arthropod-Plant Interactions 11(4): 495–505. 10.1007/s 11829-016-9475-3 · doi ↗
- 5Beimforde C Tuovila H Schmidt AR Lee WG Gube M Rikkinen J (2017 b) Chaenothecopsisschefflerae (Ascomycota: Mycocaliciales): a widespread fungus on semi-hardened exudates of endemic New Zealand Araliaceae.New Zealand Journal of Botany 55(4): 387–406. 10.1080/0028825 X.2017.1360368 · doi ↗
- 6Beimforde C Mitchell JK Rikkinen J Schmidt AR (2020) Sareomycetes cl. nov.: A new proposal for placement of the resinicolous genus Sarea (Ascomycota, Pezizomycotina).Fungal Systematics and Evolution 6: 25–37. 10.3114/fuse.2020.06.0232904095 PMC 7451776 · doi ↗ · pubmed ↗
- 7Butin H Pehl Lde Hoog GS Wollenzien U (1996) Trimmatostromaabietis sp. nov. (hyphomycetes) and related species.Antonie van Leeuwenhoek 69: 203–209. 10.1007/BF 003996078740901 · doi ↗ · pubmed ↗
- 8Cabrita P (2018) Resin flow in conifers.Journal of Theoretical Biology 453: 48–57. 10.1016/j.jtbi.2018.05.02029782932 · doi ↗ · pubmed ↗