Two new species of Phaeoclavulina (Gomphaceae, Gomphales) from North China based on morphological and phylogenetic analysis

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
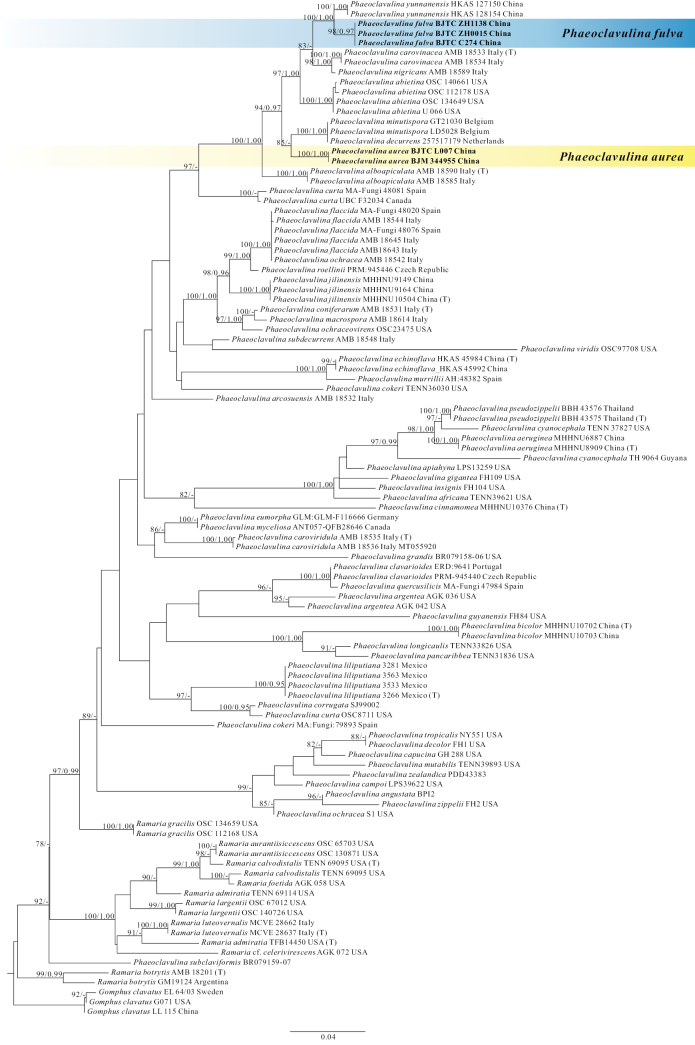 Figure 1
Figure 1 Figure 2
Figure 2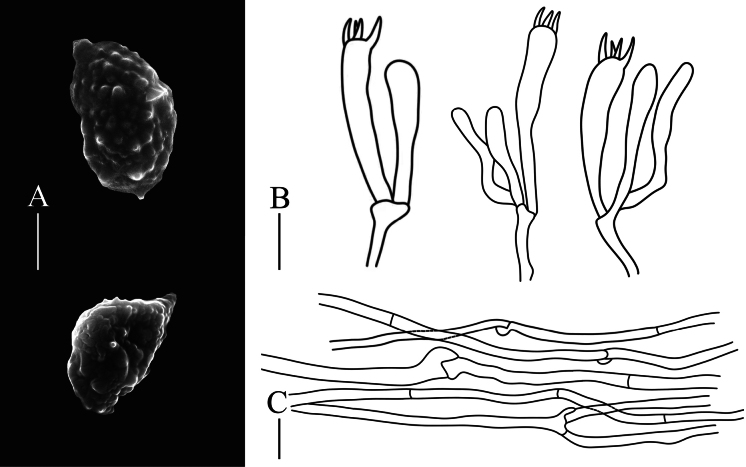 Figure 3
Figure 3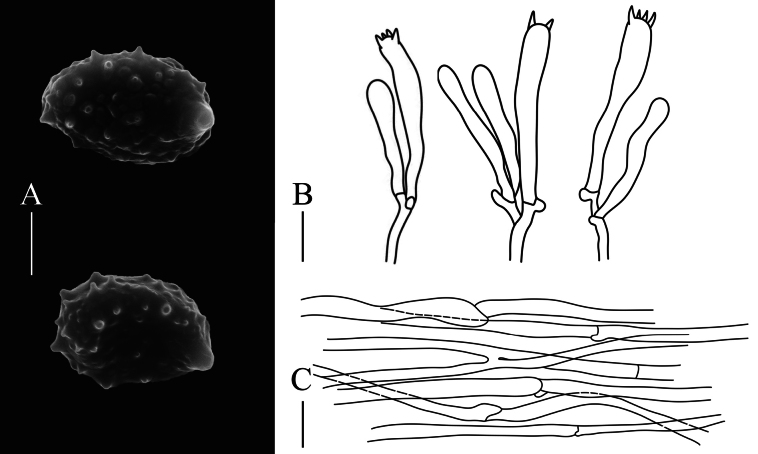 Figure 4
Figure 4| Taxonomy | Voucher/strain | Location | References | GenBank Number | |
|---|---|---|---|---|---|
| nrLSU | nrITS | ||||
|
| LL 115 | China |
| - |
|
|
| G 071 | USA | Unpublished |
| - |
|
| EL 64/03 | Sweden | Larsson et al. (2007) |
|
|
|
| U 066 | USA | Unpublished | - |
|
|
| OSC 134649 | USA | Unpublished |
|
|
|
| OSC 140661 | USA | Unpublished | - |
|
|
| OSC 112178 | USA | Unpublished |
| - |
|
| MHHNU6887 | China |
|
|
|
|
| MHHNU8909 (T) | China |
|
|
|
|
| TENN 39621 | USA |
|
| - |
|
| AMB 18590 (T) | Italy |
|
|
|
|
| AMB 18585 | Italy |
|
|
|
|
| BPI2 | - | Unpublished |
| - |
|
| LPS 13259 | USA | Unpublished |
| - |
|
| AMB 18532 | Italy |
|
|
|
|
| AGK 036 | USA | Unpublished | - |
|
|
| AGK 042 | USA | Unpublished | - |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| MHHNU10702 (T) | China |
|
|
|
|
| MHHNU10703 | China |
|
|
|
|
| LPS39622 | USA | Unpublished |
| - |
|
| GH 288 | USA | Unpublished |
| - |
|
| AMB 18533 (T) | Italy |
| - |
|
|
| AMB 18534 | Italy |
| - |
|
|
| AMB 18535 (T) | Italy |
|
|
|
|
| AMB 18536 | Italy |
|
|
|
|
| MHHNU10376 (T) | China |
|
|
|
|
| ERD-9641 | Portugal | Unpublished | - |
|
|
| PRM:945440 | Czech Republic | Kříž et al. (2019) |
|
|
|
| TENN 36030 | USA |
|
| - |
|
| MA:Fungi:79893 | Spain |
| - |
|
|
| AMB 18531 (T) | Italy |
|
|
|
|
| SJ99002 | - | Larsson et al. (2004) |
| - |
|
| MA-Fungi 48081 | Spain | Unpublished | - |
|
|
| UBC | Canada | Unpublished | - |
|
|
| OSC 8711 | USA |
|
| - |
|
| TH 9064 | Guyana | Unpublished |
|
|
|
| TENN 37827 | USA |
|
| - |
|
| FH 1 | USA | Unpublished |
| - |
|
| 257517179 | Netherlands | Unpublished | - |
|
|
| HKAS 45984 (T) | China |
|
|
|
|
| HKAS 45992 | China |
|
|
|
|
| GLM:GLM-F116666 | Germany | Unpublished | - |
|
|
| 48020 | Spain | Unpublished | - |
|
|
| 48076 | Spain | Unpublished | - |
|
|
| AMB 18544 | Italy |
|
|
|
|
| AMB 18645 | Italy |
|
|
|
|
| Italy |
|
|
| |
|
|
|
|
|
|
|
|
|
|
|
| - |
|
|
|
|
|
|
|
|
|
| FH 109 | USA |
|
| - |
|
| BR079158-06 | USA |
|
| - |
|
| FH84 | USA |
|
| - |
|
| FH104 | USA |
|
| - |
|
| MHHNU9149 | China |
|
|
|
|
| MHHNU9164 | China |
|
|
|
|
| MHHNU10504 (T) | China |
|
|
|
|
| 3281 | Mexico |
|
| - |
|
| 3533 | Mexico |
|
| - |
|
| 3266 (T) | Mexico |
|
| - |
|
| 3563 | Mexico |
|
| - |
|
| TENN 33826 | USA |
|
| - |
|
| AMB 18614 | Italy |
| - |
|
|
| GT21030 | Belgium | Unpublished |
|
|
|
| LD5028 | Belgium | Unpublished |
|
|
|
| AH:48382 | Spain |
| - |
|
|
| TENN 39893 | USA | Unpublished |
| - |
|
| ANT057- | Canada | Unpublished | - |
|
|
| AMB 18589 | Italy |
|
|
|
|
| S 1 | USA | Unpublished |
| - |
|
| AMB 18542 | Italy |
|
|
|
|
| OSC 23475 | USA |
|
| - |
|
| TENN31836 | USA |
|
| - |
|
| BBH 43576 | Thailand |
|
|
|
|
| BBH 43575 (T) | Thailand |
|
|
|
|
| MA-Fungi 47984 | Spain | Unpublished | - |
|
|
| PRM:945446 | Czech Republic | Kříž et al. (2019) | - |
|
|
| BR079159-07 | - |
|
| - |
|
| AMB 18548 | Italy |
|
|
|
|
| NY551 | USA | Unpublished |
| - |
|
| HKAS 127150 | China |
|
|
|
|
| HKAS 128154 | China |
|
|
|
|
| - | Unpublished |
| - | |
|
| FH2 | USA | Unpublished |
| - |
|
| TENN 69114 | USA |
|
| - |
|
| TFB 14450 (T) | USA |
| - |
|
|
| OSC 65703 | USA | Unpublished |
|
|
|
| OSC 130871 | USA | Unpublished |
|
|
|
| AMB 18201 (T) | Italy |
|
|
|
|
| GM19124 | Argentina | González et al. (2022) |
|
|
|
| TENN 69095 | USA |
|
| - |
|
| TENN 69095 (T) | USA |
| - |
|
|
| AGK 072 | USA | Unpublished | - |
|
|
| AGK 058 | USA | Unpublished |
| - |
|
| OSC 134659 | USA | Unpublished |
|
|
|
| OSC 112168 | USA | Unpublished |
|
|
|
| OSC 67012 | USA | Unpublished |
|
|
|
| OSC 140726 | USA | Unpublished |
| - |
|
| MCVE 28662 | Italy | Franchi et al. (2015) | - |
|
|
| MCVE 28637 (T) | Italy | Franchi et al. (2015) | - |
|
- —National Natural Science Foundation of China 501100001809 https://ror.org/01h0zpd94 http://doi.org/10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMycorrhizal Fungi and Plant Interactions · Plant Pathogens and Fungal Diseases · Plant and Fungal Species Descriptions
Introduction
PhaeoclavulinaBrinkmann (1897: 197) (Gomphaceae, Gomphales), with P.macrosporaBrinkmann (1897: 198) as the type species, was established in 1897 (Brinkmann 1897). It includes gomphoid and ramarioid forms characterized by ramarioid, unipileate, or merismatoid basidiomata. When pileate, basidiomata are glabrous or subtomentose and infundibuliform or flabelliform (González-Ávila et al. 2020). The genus exhibits a diverse palette of colors on the pileus and branch surfaces, varying from white, brown-green, pale to sordid olivaceous, violet, brown-yellow, and red cinnamon, through to gray, brick red, and pale to dark orange-yellow, even blue-green. Spores show the presence of clamps and are echinulate to verrucose, subreticulate, or reticulate (Giachini 2004; Giachini and Castellano 2011). Some Phaeoclavulina species are ectomycorrhizal. For example, P.abietina (Pers.) Giachini (2011: 189) is associated with Betula, Pinus, and Pseudotsuga (Herrera et al. 2002; Norvell and Exeter 2004); P.flaccida (Fr.)Giachini (2011: 192) with Pinus and Quercus (Kim et al. 2003); P.zippelii (Lév.) Overeem (1923: 262) with Acacia, Casuarina, and Eucalyptus (Sims et al. 1997); and P.cyanocephala (Berk. & M.A. Curtis) Giachini (2011: 191) with Abies (Estrada-Torres 1994).
Initially, Brinkmann (1897) placed Phaeoclavulina within the family Clavariaceae, along with other genera, viz. Clavaria Vaill. ex L. (1753: 1182), Clavariella P. Karst. (1881: 21), Clavulina J. Schröt. (1888: 442), and Typhula (Pers.) Fr. (1818: 296). Overeem (1923) recognized Phaeoclavulina as a distinct genus and assigned one species, P.zippelii, to it, which had previously been classified under Clavaria based on macromorphological characteristics. Despite this, the recognition of Phaeoclavulina did not gain widespread support among mycologists, and species of this genus were often accommodated with other ramarioid fungi having ochre spores in Ramaria Fr. ex Bonord. (1851: 166) (González-Ávila et al. 2020). Corner (1970) went further by placing the ramarioid species of Phaeoclavulina within Ramariasubgen.EchinoramariaCorner (1970: 238), a classification later adopted by other authors such as Marr and Stuntz (1973) and Petersen (1981). In recent research, Phaeoclavulina was once again recognized as a valid genus based on morphological, molecular, and phylogenetic data (Giachini 2004; Giachini et al. 2010; Giachini and Castellano 2011). It now includes not only the original species but also some gomphoid species with spiny, verrucose, subreticulate, or reticulate spores and a terrestrial and/or lignicolous substrate affinity (González-Ávila et al. 2020). Currently, approximately 57 species of Phaeoclavulina have been described from temperate and tropical ecosystems; however, it is likely more abundant in the tropics and subtropics (González-Ávila et al. 2020; Liu et al. 2022).
In China, 22 Phaeoclavulina species have been reported based on morphological features and phylogenetic analyses, viz. P.abietina, P.aeruginea P. Zhang (2022: 31), P.bicolor P. Zhang & W.H. Liu (2024: 6), P.campestris (K. Yokoy. & Sagara) Giachini (2011: 190), P.capucina (Pat.) Giachini (2011: 190), P.cinnamomea W.Q. Qin (2022: 32), P.cokeri (R.H. Petersen) Giachini (2011: 190), P.curta (Fr.) Giachini (2011: 190), P.cyanocephala, P.decolor (Berk. & M.A. Curtis) Giachini (2011: 191), P.echinoflava P. Zhang & W.H. Liu (2024: 7), P.eumorpha (P. Karst.) Giachini (2011: 191), P.flaccida, P.grandis (Corner) Giachini (2011: 193), P.jilinensis P. Zhang & W.H. Liu (2024: 9), P.longicaulis (Peck) Giachini (2011: 193), P.macrospora, P.mutabilis (Schild & R.H. Petersen) Giachini (2011: 194), P.sikkimia (S.S. Rattan & Khurana) Giachini (2011: 194), P.viridis (Pat.) Giachini (2011: 195), P.yunnanensis W.H. Lu, D.G. Zheng, Karun. & Tibpromma (2024: 113), and P.zippelii (Liu et al. 2022; Deng et al. 2024; Zheng et al. 2024). Despite numerous prior reports from China, there remains significant potential for the discovery of new species, particularly in North China.
Recently, several Phaeoclavulina-like samples were collected during an investigation of the Yanshan Mountains (39°40′–41°20′ N, 115°–119°47′ E) in North China, a warm temperate region. In this study, two new species are described and illustrated. The nuclear ribosomal internal transcribed spacer (nrITS) and the large subunit of nuclear ribosomal RNA (nrLSU) were sequenced from dried basidiomata of each species for phylogenetic analysis.
Materials and methods
Collecting and site description
The specimens were collected from Beijing and Tianjin, North China, between 2019 and 2023, and important collection data were recorded (Rathnayaka et al. 2024). These regions have a warm temperate continental monsoon climate, which supports a diverse assemblage of plant species (Zhou et al. 2022). Deciduous broad-leaved forest and mixed coniferous and broad-leaved forest are the dominant vegetation types in the area. Notable plant species include Abiesnephrolepis (Trautv.) Maxim., Betulaplatyphylla Suk., Pinustabuliformis Carr., Populustomentosa Carrière, and Quercusmongolica Fisch. ex Ledeb. (Wang et al. 2021; Zhou et al. 2022). The annual precipitation is approximately 700 mm (Zhou et al. 2022), and the elevation ranges from 200 to 2200 m. The collected specimens were dehydrated using an electric dryer (Dorrex) at 50 °C and then deposited in the Herbarium of the College of Life Sciences, Capital Normal University, Beijing, China (BJTC), and the Herbarium of the Natural History Museum of China, Beijing, China (NNHMC).
Morphological observation
Macroscopic characteristics of the specimens were recorded, including basidiomata color, size, branching pattern, stipe color, morphology, and dimensions. Microscopic features were analyzed by examining thin sections mounted in 3% potassium hydroxide (KOH) or sterilized water. The morphology and dimensions of microscopic structures were observed and recorded using a light microscope (Olympus DP71, Tokyo, Japan). In the description of basidiospores, the abbreviation n/m/p indicates that n basidiospores were measured from m basidiomata of p collections. Measurements and Q values are presented in the format (a)b–c(d), where “a” represents the minimum value, “b–c” the 10% to 90% range, and “d” the maximum value. Q represents the ratio of basidiospore length to width in side view (Liu et al. 2022), and Q_m_ represents the average Q value of all measured basidiospores ± the sample standard deviation (Liu et al. 2022). Nomenclatural details were submitted to MycoBank. Color terms followed the designations provided by the website ColorHexa (https://www.colorhexa.com).
DNA extraction, PCR amplification, and sequencing
DNA extraction was carried out using the M5 Plant Genomic DNA Kit (Mei5 Biotechnology Co., Ltd., China). The extracted DNA was dissolved in 1× TE buffer and stored at –20 °C for later use. PCRs were performed using a Bio-Rad S1000 thermal cycler (Bio-Rad Laboratories, Inc., USA). The primer set ITS5/ITS4 (Vilgalys and Hester 1990; White et al. 1990) was used to amplify the nrITS region, and LR0R/LR5 (Vilgalys and Hester 1990) was used for the nrLSU region. PCRs were conducted in a 25 μL reaction volume containing 2 μL of DNA template, 1 μL of each primer (10 μM), 12.5 μL of 2× Master Mix (Mei5 Biotechnology Co., Ltd., China), and 8 μL of ddH_2_O. PCR amplification conditions for nrITS followed Wannathes et al. (2018), Liu et al. (2022), and Gao et al. (2024), and the conditions for nrLSU followed Yan et al. (2020) and Sui et al. (2023). All DNA sequences were generated by Sangon Biotech (Shanghai) Co., Ltd.
Molecular phylogenetic analyses
The newly obtained sequences were submitted to NCBI (https://www.ncbi.nlm.nih.gov). The nrITS and nrLSU sequences were aligned with selected sequences from GenBank and previously published literature (Deng et al. 2024). All sequences used are listed in Table 1. The raw reads of the DNA sequences were processed to obtain consensus sequences using SeqMan 7.1.0 in the DNASTAR Lasergene Core Suite software (DNASTAR Inc., Madison, WI, USA). Sequence alignment was performed using MAFFT 6 (Katoh and Toh 2010), and manual trimming was carried out in MEGA 6 (Tamura et al. 2013). For phylogenetic analyses, newly obtained sequences and additional reference sequences of Phaeoclavulina and Ramaria species were included in the nrITS and nrLSU dataset (Table 1), with Gomphusclavatus (Pers.) Gray designated as the outgroup following Wannathes et al. (2018).
To estimate maximum likelihood (ML) gene trees, RAxML 7.4.2 Black Box software was used (Stamatakis 2006; Stamatakis et al. 2008; Zhou and Hou 2019; Zhou et al. 2021), employing a GTRGAMMAI site substitution model (Guindon et al. 2010). Branch support was assessed with 1000 bootstrap (BS) replicates (Hillis and Bull 1993). Bayesian Inference (BI) analysis was conducted using MrBayes 3.1.2 (Ronquist and Huelsenbeck 2003), with a Markov chain Monte Carlo (MCMC) algorithm (Rannala and Yang 1996). The best-fit substitution model was determined using MrModeltest 2.3 (Zhou and Hou 2019; Zhou et al. 2021, 2022). The selected models were SYM + G for nrITS and SYM + I + G for nrLSU. Two MCMC chains were run for 10,000,000 generations, terminating when the average standard deviation of split frequencies fell below 0.01. Trees were sampled every 1000 generations, with the first 25% discarded as burn-in. Significant Bayesian posterior probabilities (pp) were calculated for branches in the resulting majority-rule consensus trees. The analyses yielded relatively stable topologies, and clades with high pp values reflected the phylogenetic relationships among species (Posada and Crandall 1998).
Results
Molecular phylogeny
A total of nine sequences, including five for nrITS and four for nrLSU, were newly generated in this study. The nrITS and nrLSU datasets were compiled to investigate the phylogenetic position of the new species in Phaeoclavulina. The combined nrITS–nrLSU dataset included 146 sequences (69 for nrITS and 77 for nrLSU), representing 102 samples. The concatenated alignment contained 840 characters, including gaps. Maximum Likelihood (ML) and Bayesian Inference (BI) analyses yielded highly similar tree topologies; therefore, only the tree inferred from the ML analysis is shown (Fig. 1).
Phylogenetic tree generated from an ML analysis based on nrITS-nrLSU sequences. Numbers representing maximum likelihood bootstrap support (MLBS ≥ 75%, left) and significant Bayesian posterior probability (BPP ≥ 0.95, right) are indicated above the nodes. Novel sequences are printed in bold. Voucher specimens and localities where the specimens were collected are provided behind the species names.
The tree topology is consistent with that of Zheng et al. (2024). In the phylogenetic reconstruction, each of the two new species, P.aurea and P.fulva, formed a distinct monophyletic lineage within the Phaeoclavulina clade (Fig. 1). The specimens of P.aurea (BJM 344955 and BJTC L007) and P.fulva (BJTC C274 and BJTC ZH1138) were identified as the sister species of P.yunnanensis, with strong statistical support (MLB = 100%, BPP = 1.00). Phaeoclavulinaaurea (BJM 344955 and BJTC L007) formed a clade with P.minutispora and P.decurrens, although this relationship was supported by a relatively lower value (MLB = 85%, BPP = 0.83). In conclusion, the ML and BI analyses of the nrITS–nrLSU dataset support the recognition of two new species of Phaeoclavulina from China, viz. P.aurea and P.fulva.
Taxonomy
Phaeoclavulina
aurea
Taxon classificationFungiGomphalesGomphaceae
Y. Gao, X. Tong, & C.L. Hou sp. nov.
E6FF7311-A747-5C13-B2A0-355B5531A117
855628
Diagnosis.
Phaeoclavulinaaurea differs from the known species of Phaeoclavulina in its pale yellow to golden yellow basidiomata, tips brownish black when age, truncate spines basidiospores 4.7–6.3 × 3.0–3.7 μm, basidia 25–50 × 4.6–7.4 μm, clamp connections present.
Morphology of Basidiomata. A–C. Phaeoclavulinaaurea (A, B. BJM 344955 holotype; C. BJTC L007); D–F. Phaeoclavulinafulva (D. BJTC C274 holotype; E. BJTC ZH0015; F. BJTC ZH1138). Scale bars: 1 cm.
Etymology.
The epithet “aurea” refers to the yellow to golden yellow basidiomata.
Microscopic characteristics of Phaeoclavulinaaurea: A. Basidiospores; B. Basidia; C. Tramal hyphae. Scale bars: 20 µm (A); 10 µm (B, C).
Type.
China • Beijing, Yanqing District, Songshan, 40°30'28"N, 115°54'E, elev. 869 m, 16 Aug. 2023, H. Zhou, Y. Gao & X. Tong (BJM 344955). GenBank nrITS: PQ287856, nrLSU: PQ287860.
Description.
Basidiomata coralloid, solitary or scattered; individual basidiomata 3–8 cm tall, 2–5.5 cm wide across branches. Stipe 0.4–0.9 cm tall, 0.3–0.5 cm wide, subclavate to flattened, snow-white rhizomorphic strands near the ground, branches di- or tri-dichotomous, generally 3–4 times, cylindrical, pale yellow (#ffdd9a) to golden yellow (#ffc44d); as maturity deepens, tips blunt and short with brownish black (#473826), stipe surface white (#ffffff) to pale yellow (#ffdd9a). No color change when bruised. Odor and taste not recorded.
Basidiospores [70/2/2] (4.4–)4.7–6.3(− 6.7) ×3.0–3.7(− 4.2) μm, Q = 1.4–1.9(− 2.1), Q_m_ = 1.66 ± 0.16, ellipsoid, pale yellow to golden yellow in KOH, with thick wall and cyanophilic ornamentation in cotton blue; when seen with SEM, conical, truncate spines up to 0.6 µm high; with oleiferous guttule contents. Hilar appendixes acuminate (up to 1.1 µm in length). Basidia 25–50 × 4.6–7.4 µm, clavate, 3–4 sterigmata occur per basidium, 3.6–6.0 μm long, and cornute, clamped. Basidioles abundant, subclavate to subcylindrical. Tramal hyphae in the stipe smooth, thin-walled, hyaline in KOH, 2.3–5.8 μm wide; tramal hyphae in branch with hyaline and thin-walled, 1.8–4.0 μm wide; clamp connections abundant, H-connections present. Hyphal system monomitic. Cystidia absent.
Habit, habitat, and distribution.
Solitary or gregarious caespitose in humus layers on soils in mixed coniferous and broad-leaved forests, associated with Pinus L. Basidiomata generally occur in August; currently known only from Beijing, China.
Additional specimens examined.
China • Beijing, Huairou District, Hongluo Temple, 40°22'26"N, 116°37'26"E, elev. 153 m, 13 Aug. 2019, J.Q. Li, X.Y. Shen & R.T. Zhang (BJTC L007).
Notes.
Phaeoclavulinaaurea is morphologically similar to P.cinnamomea, which was originally described from China. However, P.cinnamomea produced cinnamon-to-salmon-orange basidiomata and relatively large basidiospores (12–15 × 5–7 µm) and basidia (40–60 × 8–12 µm) compared P.aurea (Liu et al. 2022). Phaeoclavulinaangustata (Lév.) Giachini (2011: 189) also has crowded branches, but the color of that is pale pink, and all parts soon turned sordid vinaceous on bruising, and the cyanophilic spines of basidiospores 1–3.5 µm tall (Giachini 2004). Phaeoclavulinaglaucoaromatica (R.H. Petersen) Giachini (2011: 193) has similar basidiomata with P.aurea, which doesn’t have molecular data. But P.glaucoaromatica differs from P.aurea by its branches becoming watery where handled and then changing color to olive-green; basidiospores are larger (8.2–11.1 × 4.4–5.5 µm) (Petersen 1981). Phaeoclavulinavinaceipes (Schild) Giachini (2011: 195) has the same branch color as P.aurea, which also doesn’t have molecular data. But P.vinaceipes’s stipe context bruising violet when exposed or handled, per basidium occur 4 sterigmata, and basidiospores are wider (4.8–8 × 2.7–3.8 µm) (Schild 1990). Phaeoclavulinaaurea is phylogenetically close to P.abietina, P.alboapiculata Franchi & M. Marchetti (2020: 1), and P.minutispora Franchi & M. Marchetti (2020: 3) (Fig. 1), but the stipes of P.abietina quickly turned deep blue-green when handled or confined (González-Ávila et al. 2020); the branches of P.alboapiculata are with white tips when young, and the white context gradually turns to wine when exposed to the air, and it has smaller basidiospores (4.1–5.5 × 2.5–3.4 µm) and basidia (22–30 × 5.0–6.5 µm) than P.aurea (Index Fungorum 2024); P.minutispora can be distinguished by its basidiomata surface, which gradually shows dark brown pigmentation when touched, white flesh turns maroon rapidly when exposed to air, and producing relatively small basidiospores (3.7–5.0 × 2.5–3.6 µm) and basidia (22–30 × 5–6.5 µm) (Index Fungorum 2024).
Phaeoclavulina
fulva
Taxon classificationFungiGomphalesGomphaceae
Y. Gao, X. Tong, & C.L. Hou sp. nov.
CEA9616D-989F-5274-ACBE-217BC04E3B65
855499
Diagnosis.
Phaeoclavulinafulva differs from the known species of Phaeoclavulina in its dirty orange basidiomata, stipe surface pale yellow, color lightens towards tips, rounded warts basidiospores 4.8–7.8 ×2.8–3.8 μm, basidia 22–43 × 4.3–7.3 μm.
Microscopic characteristics of Phaeoclavulinafulva: A. Basidiospores; B. Basidia; C. Tramal hyphae. Scale bars: 20 µm (A); 10 µm (B, C).
Etymology.
The epithet “fulvus” refers to the dirty orange to yellowish-brown basidiomata.
Type.
China • Tianjin, Jizhou District, Limutai Scenic Spot, 40°12'47"N, 117°2'7"E, elev. 658 m, 21 Aug. 2020, G.Q. Cheng, R.T. Zhang & C.L. Hou (BJTC C274). GenBank nrITS: PQ287852, nrLSU: PQ287857.
Description.
Basidiomata coralloid, solitary or scattered; individual basidiomata 4.2–12.5 cm tall, 4.5–10 cm wide across branches. Stipe 1.2–3.8 cm tall, 0.3–0.5 cm wide, subclavate to flattened, snow-white rhizomorphic strands near the ground, branches dichotomous, generally 3–4 times, subcylindrical, dirty orange (#d5995b) to yellowish-brown (#a27f16), tips slightly acute, stipe surface pale yellow (#fff7dc), color lightens towards tips. No color change when bruised. Odor and taste not recorded.
Basidiospores [90/4/3] (4.3–)4.8–7.8 × (2.5–)2.8–3.8(− 4.2) μm, Q = 1.5–2.2(− 2.37), Q_m_ = 1.89 ± 0.19, elongate-ellipsoid, pale yellow to golden yellow in KOH, with thick wall and cyanophilic ornamentation in cotton blue; when seen with SEM, conical, truncate warts up to 0.6 µm high; with oleiferous guttule contents. Hilar appendixes acuminate (up to 1.0 µm in length). Basidia 22–43 × 4.3–7.3 µm, clavate, 2–4 sterigmata occur per basidium, 1.8–7.0 μm long, and cornute, clamped. Basidioles abundant, subclavate to subcylindrical. Tramal hyphae in the stipe smooth, thin-walled, hyaline in KOH, 2.2–7.5 μm wide; tramal hyphae in branch with hyaline and thin-walled, 2.5–6.5 μm wide; clamp connections abundant, H-connections present. Hyphal system monomitic. Cystidia absent.
Habit, habitat, and distribution.
Solitary or gregarious caespitose in humus layers on soils in evergreen coniferous forests and deciduous broad-leaved forests, associated with Pinus L. and Populus L. Basidiomata generally occur from August; currently known only from Tianjin and Beijing, China.
Additional specimens examined.
China • Beijing, Huairou District, Hongluo Temple, 40°22'34"N, 116°37'28"E, elev. 109 m, 13 Aug. 2019, H. Zhou, G.Q. Cheng & C.L. Hou (BJTC ZH0015); China • Beijing, Huairou District, Erdaogou, 40°52'36"N, 116°31'23"E, elev. 780 m, 24 Aug. 2020, H. Zhou, X.Y. Shen & X.B. Huang (BJTC ZH1138).
Notes.
Phaeoclavulinafulva generally shares similar branching with P.sikkimia, P.cokeri, and P.articulotela (R.H. Petersen) Giachini (2011: 190). Phaeoclavulinasikkimia and P.articulotela lack molecular data (Petersen 1981). However, P.sikkimia differs from P.fulva in its relatively large basidiospores (6.5–9 × 4.5–6 µm) and basidia (60–70 × 7.5–9 µm) and in the presence of cyanophilic warts up to 2 µm tall (Giachini 2004). P.cokeri has cinnamon- to pink-yellow branches that become orange or dark red-brown, with pale orange tips; it also features strongly cyanophilic basidiospore spines 1–2 µm tall and relatively large basidiospores (9–16 × 4–7.5 µm) and basidia (45–80 × 7.5–12 µm) (Giachini 2004). P.articulotela has orange to dark orange basidiomata and relatively large basidiospores (6–9.5 × 3–5 µm) (Giachini 2004). Phylogenetically, P.fulva is related to P.abietina, P.carovinacea Franchi & M. Marchetti (2020: 2), P.nigricans E. Campo, Franchi & M. Marchetti (2020: 4), and P.yunnanensis in the analyses of the multilocus datasets, respectively (Fig. 1). However, P.abietina has a stipe that is olive-ochraceous to dull ocher upward, quickly turning deep blue-green when handled or confined; its branches are yellow-ocher to dull ocher when fresh or somewhat greenish-ocher (González-Ávila et al. 2020). Phaeoclavulinacarovinacea differs from P.fulva in its white flesh, which turns reddish brown when exposed to air (Index Fungorum 2024). Phaeoclavulinanigricans has basidiomata whose surface gradually develops a burgundy coloration when touched, and its light cream flesh turns black rapidly upon exposure to air (Index Fungorum 2024). Phaeoclavulinayunnanensis is characterized by its yellow basidiomata, with the tips showing a slight greenish-grey tint, and relatively small basidia (20–40 × 3–5 µm) (Zheng et al. 2024). The species also differs from the other new species described in this study, Phaeoclavulinaaurea, in both basidiomata color and basidiospore morphology. Phaeoclavulinaaurea has pale yellow to golden yellow basidiomata and basidiospores with truncate spines.
Discussion
The ornamentation of spores holds paramount significance in differentiating species within Phaeoclavulina. Previous studies have classified spore ornamentation in R.subg.Echinoramaria into five distinct types (Petersen 1981; Villegas et al. 2005). Volcanic spines, a distinctive form of spore ornamentation, are also evident in distinguishing Phaeoclavulina species, such as P.pancaribbea (R.H. Petersen) Giachini (2011: 194) (Petersen 1981; Giachini and Castellano 2011) and P.zealandica (R.H. Petersen) Giachini (2011: 195) (Petersen 1988; Giachini and Castellano 2011). The new species P.aurea and P.fulva described in this study also possess truncate warts or spines. However, it is noteworthy that both P.pancaribbea and P.zealandica have larger spores than the newly discovered P.aurea and P.fulva.
Some species within Phaeoclavulina exhibit host specificity. For example, P.yunnanensis has only been found under Quercus sp. (Zheng et al. 2024), P.cyanocephala is exclusively associated with Abies sp. (Estrada-Torres 2007), and the new species P.aurea discovered in this study has only been found under Pinus sp. This host specificity may serve as one of the diagnostic characteristics for species identification within Phaeoclavulina. Additionally, numerous species remain undiscovered globally, particularly in understudied forest ecosystems with favorable environmental conditions (Exeter et al. 2006), and many endemic taxa are still awaiting documentation. Therefore, further exploration of these habitats, combined with molecular and ecological studies, is essential to uncover the full diversity and host associations of Phaeoclavulina species.
Previous studies have indicated that clavarioid fungi, including Phaeoclavulina, are highly diverse and widely distributed across the globe (Corner 1950). However, to date, only 22 species of Phaeoclavulina have been described from China, despite the country’s vast territory, complex climate, diverse habitats, and rich species resources, all of which contribute to its exceptionally high fungal diversity (Miller et al. 1991). This gap may be attributed to the limited number of collections. In this study, two new species of Phaeoclavulina from North China were described using nrITS–nrLSU phylogenetic analyses (Fig. 1) and macrofungal morphological examination, thereby contributing to the current understanding of the species diversity within this genus. Moreover, specimen collection and field investigation were conducted only in August, suggesting that additional Phaeoclavulina species may yet be discovered in this study area.
Supplementary Material
XML Treatment for Phaeoclavulina aurea
XML Treatment for Phaeoclavulina fulva
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Brinkmann W (1897) Vorarbeiten zu einer Pilzflora Westfalen.Jahresbericht des Westfälischen Provinzial-Vereins für Wissenschaft und Kunst 25: 195–207.
- 2Corner EJH (1950) A Monograph of Clavaria and allied Genera.Oxford University Press, London, 740 pp.
- 3Corner EJH (1970) Supplement to “A monograph of Clavaria and allied genera”.Beihefte zur Nova Hedwigia 33: 1–299.
- 4Deng PT Liu WH Ge ZW Zhang P (2024) Three new ramarioid species of Phaeoclavulina (Gomphaceae, Gomphales) from China.Myco Keys 108: 1–14. 10.3897/mycokeys.108.12871639220354 PMC 11358615 · doi ↗ · pubmed ↗
- 5Estrada-Torres A (1994) La Familia Gomphaceae (Aphyllophorales: Fungi) en el Estado de Tlaxcala.Tesis doctorado (Doctorado en Ciencias (Biología) ENCB, IPN. México D. F., 186 pp.
- 6Estrada-Torres A (2007) Hongos ectomicorrizógenos y myxomycetes del Parque Nacional Lagunas de Montebello, Chiapas. Universidad Autónoma de Tlaxcala, Centro de Investigación en Ciencias Biológicas. Final report SNIB-CONABIO project BK 043. México, D. F.
- 7Exeter RL Norvell L Cazares E (2006) Ramaria of the Pacific Northwest.United States, Salem, Oregon, 157 pp.
- 8Franchi P Marchetti M (2022) I Funghi Clavarioidi in Italia (Clavarioid Fungi in Italy). Oviedo, Spain.