New species of Vamsapriya (Vamsapriyaceae, Xylariales) and Neolinocarpon (Linocarpaceae, Chaetosphaeriales) from Yunnan, China

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
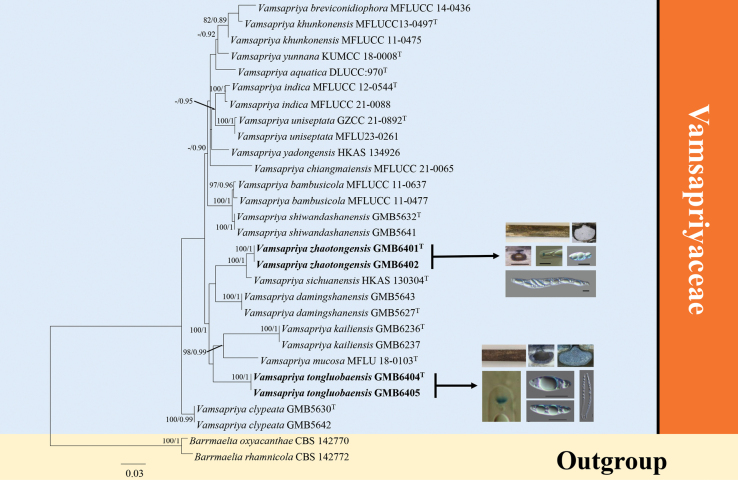 Figure 1
Figure 1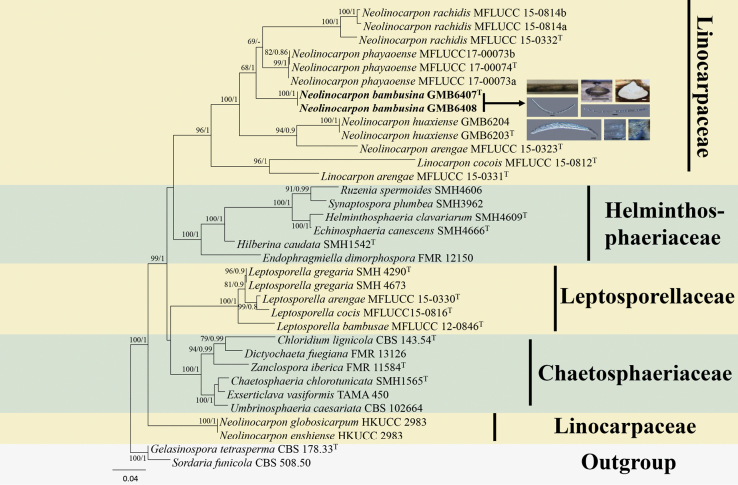 Figure 2
Figure 2 Figure 3
Figure 3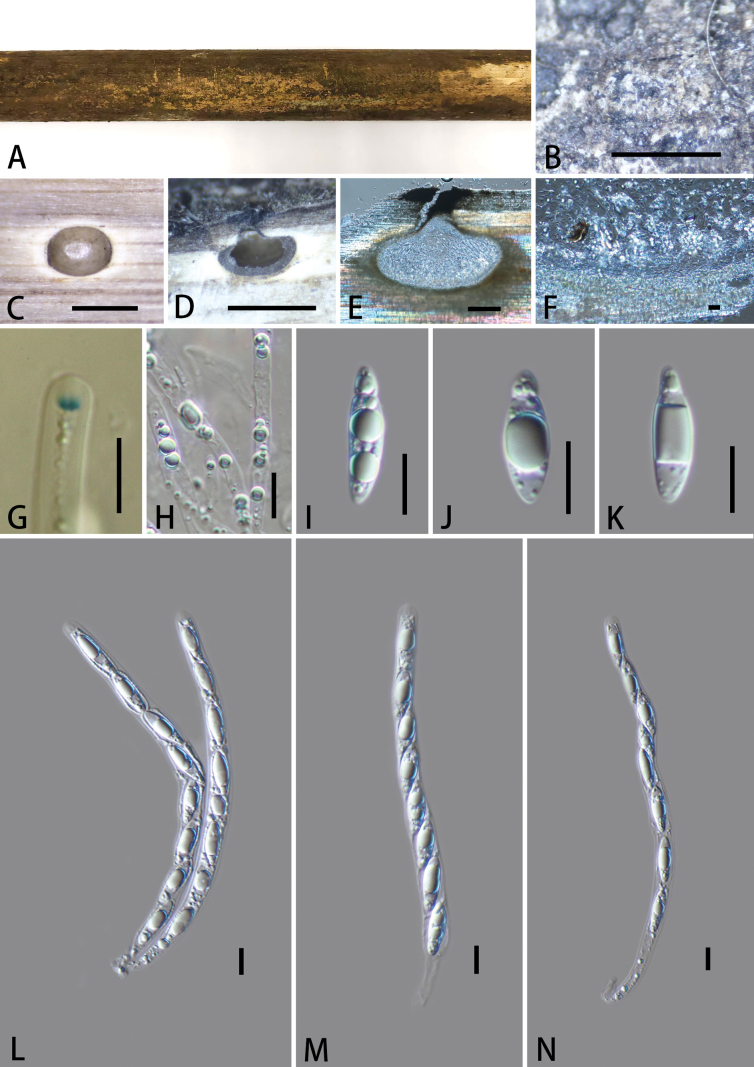 Figure 4
Figure 4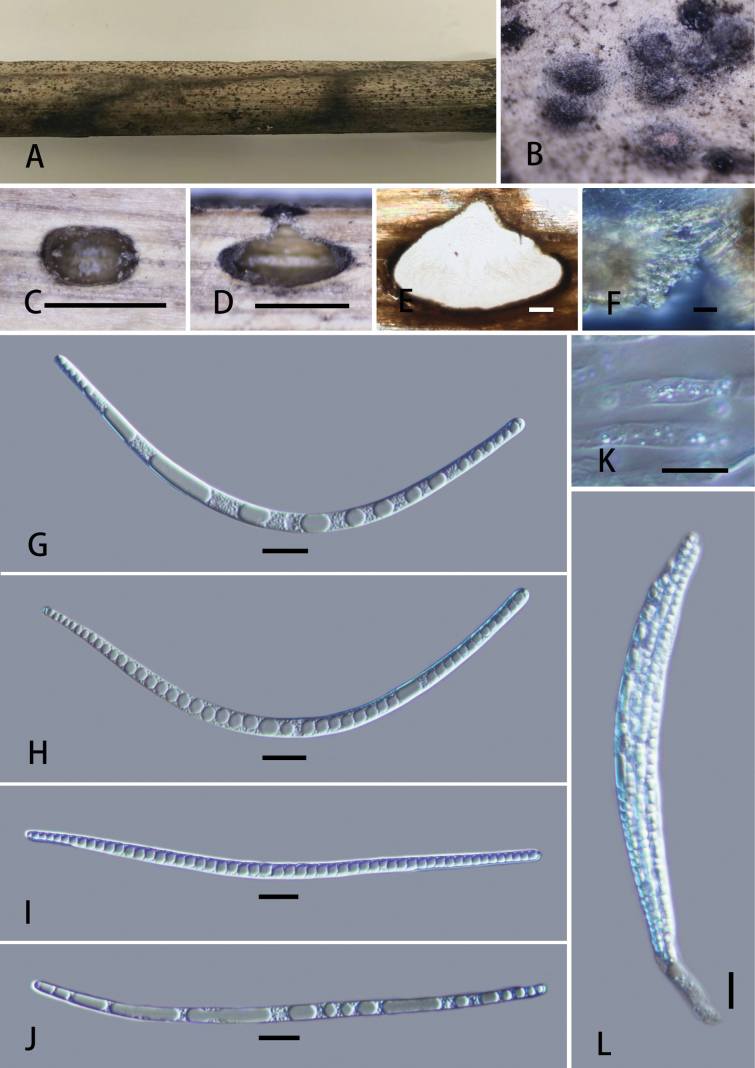 Figure 5
Figure 5| Species | Voucher / information | Host / Substrate | GenBank Accession Number | Reference | |||
|---|---|---|---|---|---|---|---|
|
|
|
|
| ||||
|
| CBS 142770 |
|
| NA |
|
|
|
|
| CBS 142772T |
|
| NA |
|
|
|
|
| DLUCC:970T | bamboo |
| NA | NA | NA |
|
|
| MFLUCC 11-0477 | bamboo |
|
|
|
|
|
| MFLUCC 11-0637 | bamboo |
|
|
| NA |
| |
|
| MFLUCC 14-0436 | bamboo |
|
| NA | NA | Direct Submission |
|
| MFLUCC 21-0065 | bamboo |
|
| NA | NA |
|
|
| bamboo |
|
| NA |
|
| |
| bamboo |
|
| NA |
|
| ||
|
| bamboo |
|
| NA |
|
| |
| bamboo |
|
| NA |
|
| ||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
| MFLUCC 12-0544T | bamboo |
|
|
|
|
|
| MFLUCC 21-0088 | bamboo |
|
|
| NA |
| |
|
| GMB6237 | bamboo |
|
| NA |
|
|
| GMB6236T | bamboo |
|
| NA |
|
| |
|
| MFLUCC 13-0497T | bamboo |
|
|
|
|
|
| MFLUCC 11-0475 | bamboo |
|
|
|
|
| |
|
| MFLU 18-0103T | bamboo |
|
|
|
|
|
|
| bamboo |
|
|
|
|
| |
| bamboo |
|
|
|
|
| ||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
| HKAS 130304T | bamboo |
|
|
| NA |
|
|
| GZCC 21-0892T | bamboo |
|
| NA | NA |
|
| MFLU23-0261 | bamboo |
|
| NA |
|
| |
|
| HKAS 134926 | wood |
|
|
|
|
|
|
| KUMCC 18-0008T | bamboo |
|
|
| NA |
|
| Species | Voucher / information | Host / Substrate | GenBank Accession Number | Reference | ||
|---|---|---|---|---|---|---|
|
|
|
| ||||
|
| SMH 1565T | decorticated wood |
| NA | NA |
|
|
| CBS 143.54T | soil |
| NA |
|
|
|
| FMR_13126 | leaves |
| NA |
|
|
|
| SMH 4666T | dead branches |
| NA | NA |
|
|
| FMR_12150 | soil |
|
|
|
|
|
| TAMA 450 | rotten wood |
| NA | NA |
|
|
| CBS 178.33T | soil |
|
| NA |
|
|
| SMH 4609T | soil |
| NA | NA |
|
|
| SMH 1542T | rotten wood |
| NA | NA |
|
|
| MFLUCC 15-0330T |
|
| NA |
|
|
|
| MFLUCC 12-0846T | bamboo |
| NA |
|
|
|
| MFLUCC 15-0816T | cocoies | NA |
|
|
|
|
| SMH 4290T | rotten wood |
| NA | NA |
|
| SMH 4673 | rotten wood |
| NA | NA |
| |
|
| MFLUCC 15-0331T |
|
|
| NA |
|
|
| MFLUCC 15-0812T |
|
|
|
|
|
|
| MFLUCC 15-0323T |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| |
|
| HKUCC2983 | dead petiole |
| NA | NA |
|
|
| HKUCC2983 | decaying intertidal fronds |
|
| NA |
|
|
| GMB6204 |
|
|
|
|
|
| GMB6203T |
|
|
|
|
| |
|
| MFLUCC 17-00073a |
|
|
| NA |
|
| MFLUCC 17-00073b |
|
|
| NA |
| |
| MFLUCC 17-00074T |
|
|
| NA |
| |
|
| MFLUCC 15-0332T |
|
|
| NA |
|
| MFLUCC 15-0814a |
|
|
|
|
| |
| MFLUCC 15-0814b |
|
|
| NA |
| |
|
| SMH4606 | rotten wood |
| NA | NA |
|
|
| CBS 508.50 | wood |
| NA |
|
|
|
| SMH 3962 | wood |
| NA | NA |
|
|
| CBS 102664 | decayed wood |
| NA | NA |
|
|
| FMR_11584T | wood |
|
|
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Mycorrhizal Fungi and Plant Interactions · Plant Diversity and Evolution
Introduction
The genus Vamsapriya was introduced by Gawas and Bhat (2005) to accommodate synnematous hyphomycetes colonizing bamboo in India, with V.indica designated as the type species. Dai et al. (2017) described the first sexual morphs of Vamsapriya and placed the genus within Xylariaceae, as accepted by Hyde et al. (2020). Later, multi-locus phylogenetic analyses by Sun et al. (2021) revealed that Vamsapriya, along with its closely related genera Diabolocovidia and Didymobotryum, formed a distinct monophyletic clade within Xylariales, clearly separated from the Xylariaceae. This finding led to the establishment of the new family Vamsapriyaceae, with Vamsapriya designated as the type genus. The asexual morph of Vamsapriya is characterized by colonies on natural substratum that are black, mycelium immersed, septate, branched, conidiophores macronematous, synnematous, erect, dark brown, cylindrical, synnemata with cylindrical to clavate apical fertile part composed of compactly arranged conidiophores; conidiogenous cells monotretic, integrated, terminal, brown, cylindrical to clavate; conidia catenate, acrogenous, cylindrical, broadly fusiform, or obclavate, and brown to dark-brown conidia (Sun et al. 2021). The sexual morph of the genus is characterized by black perithecial, immersed ascomata, sub-globose, ostiolate, surrounded by dark brown peridium, 8-spored short pedicellate asci, each with a J+ apical ring and fusiform to broadly fusiform, apiosporous ascospores, hyaline, pointed at both ends, with or without a mucilaginous sheath (Sun et al. 2021). Fourteen species of Vamsapriya have been documented worldwide, but only eight species of the genus are known to possess a sexual morph (Dai et al. 2017; Sun et al. 2021; Samarakoon et al. 2022; Dissanayake et al. 2024; Liu et al. 2025). Species of the genus are predominantly found in tropical and subtropical regions, with records from China, Cuba, India, and Thailand, with the majority of species reported from Thailand and China. Ecologically, Vamsapriya species are primarily saprobes on decaying bamboo culms (Sun et al. 2021; Samarakoon et al. 2022; Dissanayake et al. 2024; Liu et al. 2025).
The genus Neolinocarpon was introduced by Hyde (1992a) to accommodate many Linocarpon-like species, with N.globosicarpum as the type species. The genus is characterized by a clypeus with a dense, blackened, shiny central papilla; deeply immersed, oval to globose ascomata; 8-spored unitunicate asci, each with a reflective apical ring; and filiform, fasciculate ascospores containing refringent bands, with or without appendages (Hyde 1992a, b; Hyde et al. 1998; Vitória et al. 2013; Konta et al. 2017). Neolinocarpon is morphologically similar to Linocarpon, particularly in the apical structure of the ascus and ascospore morphology, but differs in having deeply immersed ascomata that form below a slightly raised or flattened clypeus, with a refractive globose body beneath the ascus apical ring (Hyde 1992b). Nearly all species of Neolinocarpon are found on hosts in the Arecaceae, except for N.penniseti and N.phayaoense, which occur on hosts from Poaceae and Euphorbiaceae, respectively (Hyde 1992b; Hyde et al. 1998; Hyde and Alias 1999; Bhilabutra et al. 2006; Vitória et al. 2013; Senwanna et al. 2018). As of March 10, 2025, Index Fungorum lists 13 species in this genus (www.indexfungorum.org).
During a mycological investigation of ascomycetes associated with decaying bamboo in Yunnan Province, Southwestern China, we found three novel saprobic bambusicolous fungi belonging to the genera Vamsapriya and Neolinocarpon, each with distinct morpho-anatomy that does not fit with any of the previously described species within their respective genera. Saprobic fungi play an important role in the natural breakdown of organic materials and often harbor biotechnologically relevant enzymes and secondary metabolites with potential applications in agriculture, medicine, and environmental management (Corbu et al. 2023). This study provides the basis for future investigations into these fungi, which may lead to valuable applied research and potential biotechnological innovations.
Materials and methods
Sample collection and morphological study
The specimens were collected during surveys conducted in Yunnan Province in China from August to November in 2024. All related habitat information was recorded. Photographs of the collected materials were taken using a Canon G15 camera (Canon Corporation, Tokyo, Japan). Materials were placed in paper bags and taken to the lab for morphological characterization and isolation. The specimens were placed in a ventilated area at room temperature for drying. All specimens were deposited at the Herbarium of Guizhou Medical University (GMB) and the Herbarium of Cryptogams, Herbarium of Kunming Institute of Botany, Chinese Academy of Sciences (KUN HKAS). Living cultures were deposited at the Guizhou Medical University Culture Collection (GMBC). All scientific names of fungi follow the entries in MycoBank and Index Fungorum; hence, no authorities and year of publication are given in the text.
Morphological characterization and isolation
Macroscopic characteristics were examined under an Olympus SZ61 stereomicroscope (Japan) and photographed with a Canon 700D digital camera (Canon Inc., Tokyo, Japan). The morphological features were studied as described by Li et al. (2024a). The samples were mounted in water for microscopic examination; Melzer’s reagent was added when necessary. The Tarosoft Image Framework (v.0.9.7) program and Adobe Photoshop CS6 software (Adobe Systems, USA) were used for measuring and processing images. Axenic cultures were obtained from single spores or tissues as described by Senanayake et al. (2015). Pure cultures on potato dextrose agar (PDA) were isolated using single-ascospore isolation techniques (Liu et al. 2025). The cultures were incubated at 25–30 °C for 4–6 weeks, with weekly observations of growth and development (Senanayake et al. 2020).
DNA extraction, PCR amplification, and sequencing
Mycelium was scraped from pure culture plates using a sterilized scalpel and used for DNA extraction following the manufacturer’s instructions for the BIOMIGA fungus genomic DNA extraction kit. The DNA samples were kept at –20 °C. Internal transcribed spacer region (ITS), large subunit (LSU), ribosomal small subunit (SSU), the second largest subunit of RNA polymerase II (rpb2), and β-tubulin (tub2) were amplified by PCR with primers ITS1/ITS4 (White et al. 1990; Gardes and Bruns 1993), LR0R/LR5 (Vilgalys and Hester 1990), NS1/NS4 (White et al. 1990), fRPB2-5F/fRPB2-7cR (Liu et al. 1999; Bischoff et al. 2006), and Bt2a/Bt2b (Glass and Donaldson 1995), respectively. The components of a 25 μL PCR mixture were 9.5 μL of double-distilled water, 12.5 μL of PCR Master Mix, 1 μL of each primer, and 1 μL of template DNA. The PCR amplification was conducted as reported by Samarakoon et al. (2022). PCR products were checked through 1.5% agarose gel electrophoresis stained with Golden View and were sent to Sangon Co., China, for sequencing.
Sequence alignments and phylogenetic analyses
All the obtained sequences were compared with known sequences in GenBank using the BLAST algorithm for precise identification (https://www.ncbi.nlm.nih.gov) (Altschul et al. 1990). The reference sequences retrieved from open databases originated from recently published data (Liu et al. 2025) and the BLASTn results of close matches. Sequences were aligned using the MAFFT v.7.110 online program (Katoh et al. 2019) with default settings. The alignments were adjusted manually using BioEdit v.7.0.5.3 (Hall 1999) where necessary. Maximum likelihood (ML) analyses were implemented in RAxML v.8.2.12 using the GTRGAMMA substitution model with 1,000 bootstrap replicates (Stamatakis 2014). Phylogenetic analyses were also performed for Bayesian inference in MrBayes v.3.2.2 (Ronquist et al. 2012) online. The Markov chain Monte Carlo (MCMC) sampling in MrBayes v.3.2.2 was used to determine the posterior probabilities (PP). Six simultaneous Markov chains were run for 1,000,000 generations, and trees were sampled every 1,000^th^ generation. The first 25% of the trees were discarded as burn-ins. The remainder was used to calculate the posterior probabilities (PPs) for individual branches. The phylogenetic tree was visualized in FIGTREE v.1.4.4 (Rambaut 2018). All analyses were run on the CIPRES Science Gateway v.3.3 web portal (Miller et al. 2010). All the obtained sequences were deposited in GenBank (Tables 1, 2).
Results
Phylogenetic analyses 1
The aligned dataset of Vamsapriyaceae (Fig. 1), comprising 3,080 characters (ITS/LSU/tub2/rpb2), contains 29 strains, including the newly generated sequences and outgroup taxa. Barrmaeliaoxyacanthae and B.rhamnicola were selected as outgroup taxa. The tree topology from the ML analysis was similar to that from the BI analysis. The best-scoring RAxML tree is presented in Fig. 1. The sequences of our specimen, Vamsapriyazhaotongensis, formed a well-supported (ML/BI: 100/1.00) clade in a sister relationship with V.sichuanensis. The lineage branch of this clade contains the second collection, Vamsapriyatongluobaensis, which shows a close relationship with V.mucosa and V.kailiensis. Although this clade lacks robust support, it remained stable in repeated phylogenetic analyses.
RAxML tree of Vamsapriya and its related taxa based on a combined LSU, ITS, rpb2, and tub2 sequences dataset. Bootstrap support values for maximum likelihood (MLB) greater than 75% and Bayesian posterior probabilities (BYPP) greater than 0.90 are displayed above the respective branches (ML/BI). The newly described species are marked in bold.
Phylogenetic analyses 2
The aligned dataset of Linocarpaceae comprised 2,454 characters (ITS/LSU/SSU) after exclusion of ambiguously aligned regions and long gaps. Sordariafimicola and Gelasinosporatetrasperma were selected as outgroup taxa. In our phylogram (Fig. 2), the sequences of our collection, Neolinocarponbambusicola, nested within the phylogenetic branch of the genus Neolinocarpon, formed a separate clade outside a group comprised of N.phayaoense and N.rechedis, demonstrating its status as an independent species.
Phylogenetic tree based on combined partial SSU, LSU, and ITS sequences showing the relationship of one new species of Neolinocarpon from Yunnan Province with other species. Numbers at the branches indicate support values (RAxML-BS/BI-PP) above 60%/0.9. Ex-type materials are marked with “T.” Materials in bold type are those analyzed in this study.
Taxonomy
Vamsapriya
zhaotongensis
Taxon classificationFungiXylarialesVamsapriyaceae
X. Y. Luo, K. Habib & Q. R. Li sp. nov.
11FFEA35-6A59-5770-B13E-46ADC72A80B8
859145
Etymology.
The specific epithet “zhaotongensis” refers to the location, Zhaotong City, where the holotype specimen was collected.
Vamsapriyazhaotongensis (GMB6401, holotype). A. Type material; B. Ascomata on the host surface; C. Cross-section of ascoma; D, E. Longitudinal sections of ascomata; F. Peridium; G. Paraphyses; H. A J+ subapical ring bluing in Melzer’s reagent; I, J. Ascospores; K–N. Asci. Scale bars: 1 mm (B–D); 100 μm (E); 10 μm (F–N).
Type.
China • Yunnan Province, Zhaotong City, Shuifu County, Tongluo Dam National Forest Park (28°26'20.40"N, 104°5'12.09"E), altitude: 1315 m, on dead culms of bamboo, Jun 2024, collected by Xing Yu Luo (holotype: GMB6401; isotype: KUN-HKAS 146990; ex-type living culture: GMBC6401).
Description.
Saprobic on dead bamboo culm. Sexual morph: Ascomata 560–675 × 720–800 μm (x̄ = 606 × 761 μm, n = 5), immersed, visible as black, circular dots, solitary, scattered, in cross-section subglobose, with mostly flattened base. Ostioles centric, ostiolar canal periphysate. Peridium 12–17 μm (x̄ = 14.1 μm, n = 10) wide, two-layered, outer layer comprising dark brown, thick-walled cells of textura angularis, inner layer composed of hyaline, thin-walled cells of textura angularis. Paraphyses 1.7–4.2 μm (x̄ =3.3 μm, n = 30) wide, longer than asci, numerous, guttulate. Asci 125–180 × 8–14.5 μm (x̄ = 151.1 × 10.7 μm, n = 25), 8-spored, unitunicate, cylindrical, short pedicellate, with a 1.6–2.7 × 3.0–4.2 μm (x̄ = 2.1 × 3.4 μm, n = 30), trapezoid apical ring, J+ in Melzer’s reagent, apex rounded. Ascospores 17.5–21 × 5.4–8.2 μm (x̄ = 19.5 × 6.7 μm, n = 30), uniseriate, hyaline, ellipsoidal to broadly fusiform, rounded at both ends, apiosporous; smaller cell 4.4–5.1 μm (x̄ = 4.7 μm, n = 30) long, larger cell 13–16 μm (x̄ = 14.8 μm, n = 30) length, usually with large guttules, lacking mucilaginous sheath. Asexual morph: Undetermined.
Culture characters.
Ascospores cultured on PDA medium at 27 °C for 4–5 weeks, colony diameter 4–4.5 cm, circular, cottony, slightly raised in center, with a distinct margin. White from above; pale brown from below, white in center.
Paratype.
China • Yunnan Province, Zhaotong City, Shuifu County, Tongluo Dam National Forest Park (28°26'14.07"N, 104°5'20.06"E), altitude: 1253 m, on bamboo, Jun 2024, collected by Xingyu Luo (GMB6402; GMBC6402).
Notes.
Phylogenetically, Vamsapriyazhaotongensis is closely related to V.sichuanensis. In the BLAST search, the closest match for the ITS sequence of Vamsapriyazhaotongensis was V.sichuanensis (HKAS 130304) with 95.59% similarity, followed by V.damingshanensis (GMB5627) with 89.84% similarity. The LSU sequences of V.zhaotongensis showed 99.51% similarity to V.sichuanensis (HKAS 130304) and 98.39% to V.damingshanensis (GMB5627). The rpb2 sequences showed 99.59% similarity to V.sichuanensis (HKAS 130304). Morphologically, V.sichuanensis can be easily distinguished from V.zhaotongensis in having ascospores surrounded by a mucilaginous sheath. Moreover, the V.sichuanensis has smaller stromata (250–400 × 350–500 μm vs. 560–675 × 720–800 μm) and smaller asci (100–160 × 7–10 μm vs. 125.9–180.9 × 8.0–14.3 μm). Vamsapriyadamingshanensis differs from V.zhaotongensis in having larger ascospores (21–25 × 7–12.5 µm vs. 17.5–21 × 5.4–8.2 μm), with a large cell (16.4–19 μm long vs. 13–16 μm long) and basal cell (5–8.5 μm long vs. 4.4–5.1 μm long), and ascospores of V.damingshanensis surrounded by a thin gelatinous mucilaginous sheath vs. lacking in V.zhaotongensis.
Vamsapriya
tongluobaensis
Taxon classificationFungiXylarialesVamsapriyaceae
X.Y. Luo, K. Habib & Q.R. Li sp. nov.
A8A590BA-A13B-55C4-8BC3-928BDE2C7539
859144
Etymology.
The specific epithet “tongluobaensis” refers to the geographical location, Tongluo Dam National Forest Park, where the holotype specimen was collected.
Vamsapriyatongluobaensis (GMB6404, holotype). A. Type material; B. Ascomata on the host surface; C. Cross-section of ascoma; D, E. Longitudinal sections of ascomata; F. Peridium; G. A J+ subapical ring bluing in Melzer’s reagent; H. Paraphyses; I–K. Ascospores; L–N. Asci. Scale bars: 0.5 mm (B–D); 100 μm (E); 10 μm (F–N).
Type.
China • Yunnan Province, Zhaotong City, Shuifu County, Tongluo Dam National Forest Park (28°26'18.26"N, 104°5'20.28"E), altitude: 1306 m, on dead culms of bamboo, Jun 2024, collected by Xingyu Luo (holotype: GMB6404; isotype: KUN-HKAS 146991; ex-type living culture: GMBC6404).
Description.
Saprobic on dead bamboo culms. Sexual morph: Ascomata 600–875 × 440–610 μm (x̄ = 791 × 550 μm, n = 5), immersed, visible as black dots, with clypeus-solitary, scattered, in cross-section globose to subglobose. Ostioles centric, raised, ostiolar canal periphysate. Peridium 46–133 μm (x̄ = 83.7 μm, n = 10) wide, two-layered, outer layer comprising dark brown, thick-walled cells of textura angularis, inner layer composed of hyaline, thin-walled cells of textura angularis. Paraphyses 2.8–6.2 μm (x̄ = 4.3 μm, n = 30) wide, long, septate, constricted at septa, guttulate. Asci 115.5–170 × 7.6–11.5 μm (x̄ = 146.5 × 8.9 μm, n = 25), 8-spored, unitunicate, cylindrical, short pedicellate, with a 2–3.4 × 1.5–2.3 μm (x̄ = 3.1 × 1.9 μm, n = 30), trapezoid apical ring, J+ in Melzer’s reagent. Ascospores 15–24 × 4.4–6.8 μm (x̄ = 19.3 × 5.9 μm, n = 30), L/W 3.3, uniseriate, hyaline, ellipsoidal, apiosporous; smaller cell 2.8–5.1 μm (x̄ = 3.7 μm, n = 30) long, conical; larger cell 13.4–21 μm (x̄ = 15.6 μm, n = 30) length, usually with large guttules, lacking mucilaginous sheath. Asexual morph: Undetermined.
Culture characters.
Ascospores cultured on PDA medium at 27 °C for 3–4 weeks, colony diameter 4–5 cm, circular, cottony surface, slightly raised in center, with a distinct margin. White from above; yellow-white to pale brown from below.
Paratype.
China • Yunnan Province, Zhaotong City, Shuifu County, Tongluo Dam National Forest Park (28°26'22.16"N, 104°5'10.06"E), altitude: 1411 m, on dead bamboo culms, Jun 2024, collected by Xingyu Luo (GMB6405; paratype living culture: GMBC6405).
Note.
Phylogenetically, Vamsapriyatongluobaensis is closely related to V.mucosa. In the BLAST search, the closest match of the ITS sequences of V.tongluobaensis showed 89.37% similarity to V.mucosa MFLU 18-0103, followed by V.kailiensis GMB6236 with 85.90% similarity. Its LSU sequences demonstrated 98.23% and 97.66% similarity to V.mucosa (MFLU 18-0103) and V.kailiensis (GMB6236), respectively, while its rpb2 sequences showed 91.64% similarity to V.mucosa (MFLU 18-0103). Morphologically, V.mucosa differs in having smaller ascomata (260–300 × 320–380 μm vs. 600–875 × 440–610 μm), smaller asci (80–95 × 5–7.5 μm vs. 115.5–170 × 7.6–11.5 of V.tongluobaensis), smaller ascospores (10–14 × 3.5–4.5 μm vs. 15–24 × 4.4–6.8 μm of V.tongluobaensis), and ascospores surrounded by a 4–10 μm wide mucilaginous sheath vs. lacking in V.tongluobaensis (Samarakoon et al. 2022).
Morphologically, Vamsapriyazhaotongensis resembles V.tongluobaensis in the lack of a mucilaginous sheath around the ascospores. However, V.zhaotongensis differs in having shorter ascospores (17.5–21 × 5.4–8.2 μm vs. 15–24 × 4.4–6.8 μm of V.tongluobaensis) with a ratio of basal to apical cell in V.zhaotongensis of 3.1, while in V.tongluobaensis, it is 4.2. Additionally, the ITS sequences of Vamsapriyatongluobaensis show 90.94% similarity to V.zhaotongensis. LSU sequences show 98.17% similarity. Based on these morphological differences and phylogeny, we introduce V.tongluobaensis as a new species.
Neolinocarpon
bambusicola
Taxon classificationFungiChaetosphaerialesLinocarpaceae
X.Y. Luo, K. Habib & Q.R. Li sp. nov.
2E5BC92E-513D-5135-A9F5-9EE14BC59945
859146
Etymology.
The specific epithet “bambusicola” refers to the host genus Bambusa (Poaceae) on which the holotype specimen was collected.
Neolinocarponbambusicola (GMB6407; holotype) A. Type material; B. Ostioles; C. Cross-section of ascoma; D, E. Longitudinal sections of ascomata; F. Peridium; G–J. Ascospores; K. Paraphyses; L. Asci. Scale bars: 1 mm (C, D); 100 μm (E); 10 μm (F–L).
Type.
China • Yunnan Province, Zhaotong City, Shuifu County, Tongluo Dam National Forest Park (28°26'14.48"N, 104°5'22.40"E), altitude: 1273 m, on dead culms of bamboo, Jun 2024, collected by Xingyu Luo (holotype: GMB6407; isotype: KUN-HKAS 146992; ex-type living culture: GMBC6407).
Description.
Saprobic on dead bamboo culm. Sexual morph: Ascomata 410–690 × 380–745 µm (x̄ = 519 × 576 µm, n = 5), immersed, perithecial, solitary, superficial, globose to subglobose, black, clypeus slightly raised. Ostioles papillate, central, black. Peridium 19–55 μm wide (x̄ = 40 μm, n = 10), outer cells merging with host epidermal cells, composed of dark brown to black cells of textura angularis. Paraphyses 2.5–4 µm wide (x̄ = 3 μm, n = 20), numerous, hyaline, hypha-like, long-cylindrical, unbranched, septate, longer than asci. Asci 142–230 × 10–15 μm (x̄ = 176.8 × 13.3 μm, n = 25), 8-spored, unitunicate, cylindrical, short-pedicellate, with a J-, wedge-shaped, subapical ring. Ascospores 106–144 × 3–5 μm (x̄ = 130 × 4.5 μm, n = 30), filiform, straight or curved, hyaline, aseptate, containing numerous refringent septum-like bands up to 50, ends rounded, lacking polar mucilaginous appendages, smooth-walled. Asexual morph: Undetermined.
Culture characteristics.
Ascospores germinating on PDA within 24 h. Colonies on PDA reached 50 mm in diameter after one month at 27 °C, colonies circular, edge entire, velvety to woolly, from above white, below yellow-white to light brown.
Paratype.
China • Yunnan Province, Zhaotong City, Shuifu County, Tongluo Dam National Forest Park (28°26'20.44"N, 104°5'16.28"E), altitude: 1098 m, on bamboo, Jun 2024, collected by Xingyu Luo (GMB6408; paratype living culture: GMBC6408).
Note.
Phylogenetic analysis (Fig. 2) based on ITS, LSU, and SSU sequences revealed Neolinocarponbambusicola to be closely related to N.phayaoense. In the BLAST search, the closest match for the LSU sequence of N.bambusicola was N.phayaoense (MFLUCC 17-00073a) with 93.2% similarity, followed by N.arengae (MFLUCC 15-0323) with 91.5% similarity. The SSU sequence of N.bambusicola showed 98.88% similarity to N.phayaoense (MFLUCC 17-00073a) and 99.29% similarity to N.arengae (MFLUCC 15-0323).
Morphologically, Neolinocarponbambusicola resembles N.phayaoense in having filiform ascospores lacking mucilaginous appendages. However, N.phayaoense can be distinguished from N.bambusicola by its host Heveabrasiliensis in Euphorbiaceae vs. bamboo, smaller ascomata with 1–3 locules (250–550 × 120–300 μm vs. 410–690 × 380–745 µm), longer asci (115–130 μm vs. 142–230 μm), and smaller ascospores (77–92 × 2–5 μm, x̄ = 85 × 4 μm vs. 106–144 × 3–5 μm, x̄ = 130 × 4.5 μm) (Senwanna et al. 2018).
Morphologically, Neolinocarponbambusicola resembles N.arengae in ascal size and the lack of mucilaginous appendages on ascospores. However, the latter differs in host preference, being reported on dead leaflets of Arengapinnata (a palm, Arecaceae), and in having slightly smaller ascospores (114–134 × 3–4 μm), whereas N.bambusicola is found on dead bamboo culms (Poaceae) and has slightly larger ascospores (106–144 × 3–5 μm). Although both species occur on monocotyledonous hosts, the two species are phylogenetically distinct. Based on these morphological differences and phylogenetic evidence, we introduce N.bambusicola as a new species (Konta et al. 2017).
Discussion
China possesses the richest bamboo resources in the world, accounting for about one-third of the global bamboo forest area (Dlamini et al. 2022). This ecological dominance is reflected in its rich fungal diversity and hosts over one-third of the world’s documented bambusicolous Ascomycota (Jiang et al. 2022). Up to 2022, 174 bambusicolous ascomycete taxa had been documented in Southwest China, representing approximately 34% of China’s total known bambusicolous Ascomycota (Jiang et al. 2022). Since then, about 150 new bambusicolous ascomycete species have been described from China, mostly from southwestern regions (Liang et al. 2023; Wu et al. 2023; Yu et al. 2023; Dissanayake et al. 2024; Habib et al. 2024; Li et al. 2024a, b; Shen et al. 2024; Yu et al. 2024a, b; Zhou et al. 2024; Liu et al. 2025). This remarkable increase highlights the hyperdiversity of bambusicolous fungi in China, and Yang et al. (2024) reported that bamboo forests in southwestern China are still unexplored for bamboo-associated fungi, with many species yet to be discovered.
This study contributes to the knowledge of bambusicolous fungal diversity in southern China through the description of three novel species assigned to the genera Vamsapriya and Neolinocarpon. All species of Vamsapriya have been reported on bamboo, except V.yadongensis, whose host remains unidentified. The two new species described in this study were also found on bamboo, suggesting that the genus may exhibit a strong host specificity. Globally, 14 species of Vamsapriya have been documented, and among these, eight species are known to possess a sexual morph (Dai et al. 2017; Sun et al. 2021; Samarakoon et al. 2022; Dissanayake et al. 2024; Liu et al. 2025). In this study, we added another Vamsapriya species from its sexual morph. The species in this genus with sexual morphs can be divided into two sections: those with a mucilaginous sheath on the ascospore and those without. Among the previously known species, V.clypeata, V.kailiensis, and V.shiwandashanensis lack a mucilaginous sheath, whereas V.bambusicola, V.chiangmaiensis, V.damingshanensis, V.mucosa, and V.sichuanensis are characterized by its presence. The species of the genus can be further differentiated by the size of ascomata and asci and the shape and size of their ascospores. The two newly recognized species of Vamsapriya do not possess a mucilaginous sheath. Among the species lacking a sheath on the ascospores, V.kailiensis possesses the smallest ascospores in the genus (<14 µm), whereas V.shiwandashanensis has the largest (20–29 × 5.5–7.5 µm). Vamsapriyaclypeata can be confused with V.zhaotongensis because of their similar ascospore size (18.5–21 × 7.2–8.9 µm) and shape. However, V.zhaotongensis differs in having slightly smaller asci (115.5–170 × 7.6–11.5 µm vs. 129–181 × 8.2–9.6 µm in V.clypeata) and is phylogenetically distantly related. Vamsapriyatongluobaensis can be easily distinguished from V.clypeata by its larger ascospores (15–24 × 4.4–6.8 µm) pointed at both ends, vs. rounded ends in V.clypeata (Liu et al. 2025).
Nearly all previously known species of Neolinocarpon have been found on hosts in the Arecaceae, indicating a strong host preference within this genus. The exceptions are N.penniseti, which occurs on the dead stems of Pennisetumpurpureum (Poaceae), and N.phayaoense, found on branches of Heveabrasiliensis (Euphorbiaceae) (Hyde 1992b; Hyde et al. 1998; Hyde and Alias 1999; Bhilabutra et al. 2006; Vitória et al. 2013; Senwanna et al. 2018). Within the Arecaceae, Neolinocarpon species are reported on different plants and are associated with various plant parts. Neolinocarponaustraliense is associated with dead rattan of Calamusmoti, while N.attaleae occurs on dead rachises of Attaleafunifera. Similarly, N.calami and N.enshiense inhabit dead petioles of Calamusconirostris and Trachycarpusfortunei, respectively. Neolinocarponglobosicarpum and N.nypicola are reported on decaying intertidal fronds and intertidal petioles of Nypafruticans. Neolinocarponinconspicuum and N.nonappendiculatum both occur on Archontophoenixalexandrae but occupy different tissues: dead rachises and dead petioles, respectively (Hyde 1992b; Hyde et al. 1998; Hyde and Alias 1999; Vitória et al. 2013). Recently, a new species, N.huaxiense, has been reported from China, isolated from bamboo (Habib et al. 2025). This study expands the known ecological range of the genus by reporting N.bambusicola on dead bamboo (Poaceae) culms, marking the third documented occurrence of Neolinocarpon on Poaceae.
The genus Neolinocarpon and its type species, Neolinocarponglobosicarpum, were introduced by the renowned mycologist K.D. Hyde, who also established several other species within the genus, including Neolinocarponarengae S. Konta & K.D. Hyde, Neolinocarponphayaoense Senwanna & K.D. Hyde, and Neolinocarponrachidis S. Konta & K.D. Hyde. The holotype sequences of these latter species have been verified and form a well-supported monophyletic clade, within which our newly described species also clusters. However, phylogenetic analyses reveal inconsistencies within the genus, particularly concerning N.globosicarpum and N.enshiense, whose available sequences, derived from non-type specimens, cluster distantly from the main Neolinocarpon clade and do not group within the familial boundaries, rendering the genus polyphyletic. This phylogenetic incongruence highlights a systematic issue within Neolinocarpon that can only be resolved by sequencing the holotype of N.globosicarpum. A detailed molecular study, including the re-examination and sequencing of existing type specimens, is urgently needed to clarify the true phylogenetic placement of these taxa. Until this is addressed, the delimitation of Neolinocarpon remains provisional, and the phylogenetic positions of N.globosicarpum and N.enshiense should be interpreted with caution rather than being excluded from the genus.
These findings not only highlight the vast, yet-to-be-explored fungal diversity in China’s bamboo ecosystems but also pave the way for further research to uncover additional species and clarify their ecological roles.
Supplementary Material
XML Treatment for Vamsapriya zhaotongensis
XML Treatment for Vamsapriya tongluobaensis
XML Treatment for Neolinocarpon bambusicola
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Altschul SF Gish W Miller W Myers EW Lipman DJ (1990) Basic local alignment search tool.Journal of Molecular Biology 215(3): 403–410. 10.1016/S 0022-2836(05)80360-22231712 · doi ↗ · pubmed ↗
- 2Bahl J (2006) Molecular evolution of three morphologically similar families in the Xylariomycetidae (Apiosporaceae, Clypeosphaeriaceae, Hyponectriaceae). HKU Theses Online.
- 3Bao DF Hyde KD Mc Kenzie EHC Jeewon R Su HY Nalumpang S Luo ZL (2021) Biodiversity of Lignicolous freshwater Hyphomycetes from China and Thailand and description of sixteen species. Journal of Fungi 7(8): e 669. 10.3390/jof 7080669 PMC 839927634436208 · doi ↗ · pubmed ↗
- 4Bhilabutra W Lumyong S Jeewon R Mc Kenzie EHC Hyde KD (2006) Neolinocarponpenniseti sp. nov. on the grass Pennisetumpurpureum (Poaceae). Cryptogamie.Mycologie 27(4): 305–310.
- 5Bischoff JF Rehner SA Humber RA (2006) Metarhiziumfrigidum sp. nov.: A cryptic species of M.anisopliae and a member of the M.flavoviride complex.Mycologia 98(5): 737–745. 10.3852/mycologia.98.5.73717256577 · doi ↗ · pubmed ↗
- 6Cai L Jeewon R Hyde KD (2006) Phylogenetic investigations of Sordariaceae based on multiple gene sequences and morphology.Mycological Research 110(2): 137–150. 10.1016/j.mycres.2005.09.01416378718 · doi ↗ · pubmed ↗
- 7Corbu VM Gheorghe-Barbu I DumbravăAȘVrâncianu COȘesan TE (2023) Current insights in fungal importance-A comprehensive review. Microorganisms 11(6): e 1384. 10.3390/microorganisms 11061384 PMC 1030422337374886 · doi ↗ · pubmed ↗
- 8Dai DQ Bahkali AH Li QR Bhat DJ Wijayawardene NN Li WJ Chukeatirote E Zhao RL Xu JC Hyde KD (2014) Vamsapriya (Xylariaceae) re-described, with two new species and molecular sequence data. Cryptogamie.Mycologie 35(4): 339–357. 10.7872/crym.v 35.iss 4.2014.339 · doi ↗