Investigating the Relationship Between Maternal Smoking During Pregnancy and Offspring Handedness: Extending the Proxy Gene-by-Environment Mendelian Randomization Study Design to Include Polygenic Risk Scores
Daisy C. P. Crick, Sarah E. Medland, George Davey Smith, David M. Evans

TL;DR
This study explores if maternal smoking during pregnancy influences offspring handedness using genetic and environmental data from UK Biobank.
Contribution
Extends proxy gene-by-environment Mendelian randomization to include polygenic risk scores for smoking behavior.
Findings
Maternal smoking during pregnancy was associated with increased right-handedness in offspring using traditional epidemiological methods.
No strong causal effect of maternal smoking on offspring handedness was found using gene-by-environment Mendelian randomization.
Factors like birth year, birthweight, and breastfeeding impact hand preference more than maternal smoking.
Abstract
Hand preference first appears in early development, yet twin studies and GWAS show that only a minority of variance is explained by heritable genetic factors. Using UK Biobank data and multivariable logistic regression to test associations between potential causes of handedness and offspring hand preference, we then investigated the potential causal effect of maternal smoking during pregnancy on offspring handedness using a proxy gene-by-environment (GxE) Mendelian randomization design. We used rs16969968 in the CHRNA5 gene and a polygenic risk score of genome-wide significant smoking-heaviness variants to proxy smoking behaviour. We stratified based on reported maternal smoking during pregnancy because, regardless of genotype, any causal effect of maternal smoking heaviness on offspring handedness should only manifest in individuals whose mothers smoked during pregnancy. Using…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
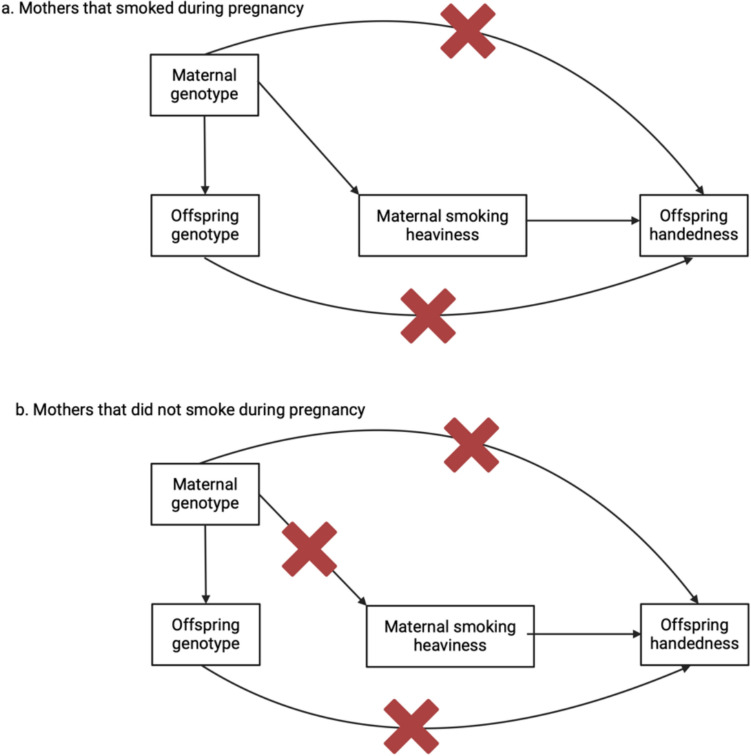 Figure 1
Figure 1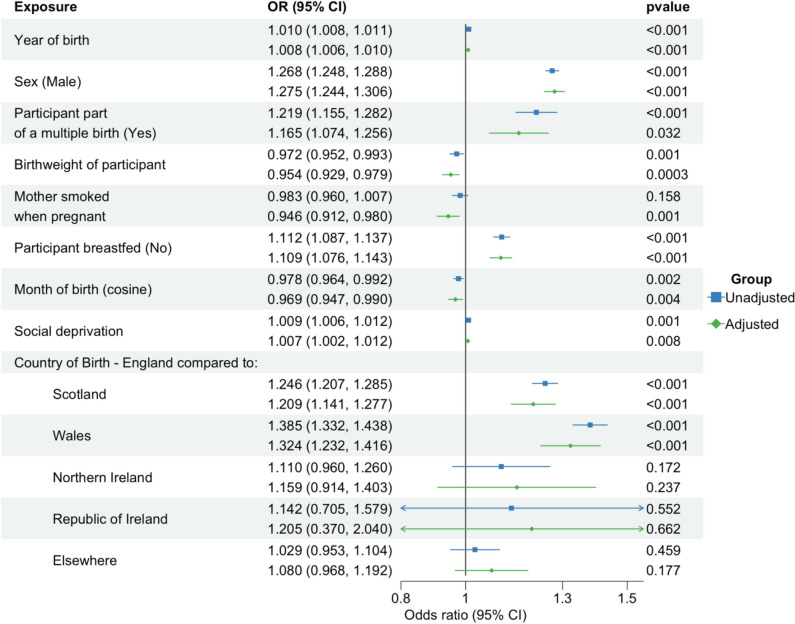 Figure 2
Figure 2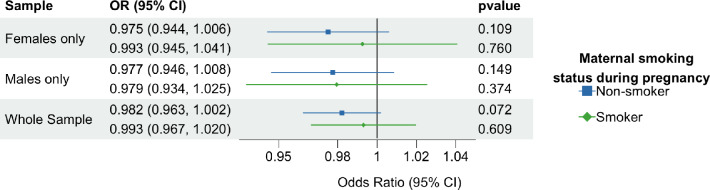 Figure 3
Figure 3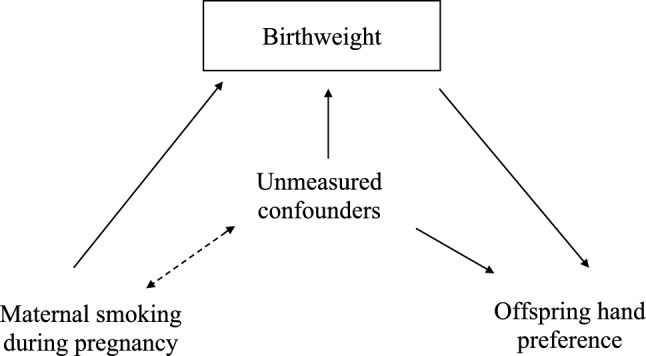 Figure 4
Figure 4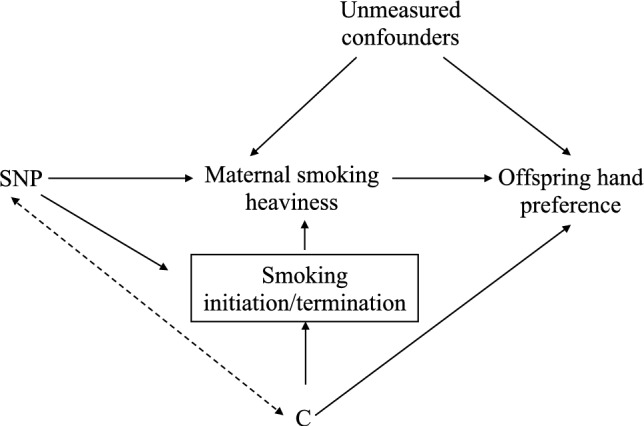 Figure 5
Figure 5- —The University of Queensland
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHemispheric Asymmetry in Neuroscience
Introduction
The majority of people (90%) are right-handed, meaning that they use their right hand for complex manual tasks such as writing (Papadatou-Pastou 2011; Papadatou-Pastou et al. 2020). The rest of the population is split into those who are left-handed (~ 10%) and a small fraction of individuals who are ambidextrous and can use either hand equally well (~ 1%) (Papadatou-Pastou et al. 2020). The first signs of hand preference and motor asymmetry in humans emerge at around 15 weeks gestation in the form of preferential lateralized thumb sucking behaviours in utero *(*Hepper et al. 1991). By 5–6 months post-partum, infants exhibit clear hand preferences for target-directed behaviours (Michel et al. 1985) which then tend to remain stable throughout childhood (Michel and Harkins 1986). Certain early life influences are thought to affect hand preference. For instance, males are more likely than females to be left-handed (Papadatou-Pastou et al. 2008; Tan 1988), and lower birthweight and antenatal complications are associated with a higher prevalence of left-handedness (Strien et al. 1987; O'Callaghan et al. 1987). Cultural pressures also appear to impact hand preference (Porac et al. 1990; Medland et al. 2004), evident through the lower frequency of left-handers in Asia compared to North America or Europe (Porac et al. 1990) and through the steady increase in left-handedness over time as societal demands for right handedness diminish (Hugdahl et al. 1993). A positive relationship between non-breastfeeding and left-handedness has also been observed (Denny 2012) a result subsequently replicated in a meta-analysis of several large scale studies (Hujoel 2019). Further, observational, associations have been reported between left-handedness/ambidexterity and a number of neurological and psychiatric disorders such as dyslexia (Packheiser et al. 2023), schizophrenia (Dragovic and Hammond 2005; Webb et al. 2013), depression (Denny 2009), bipolar disorder (Ravichandran et al. 2017), and migraines (Geschwind and Behan 1982).
There is now overwhelming evidence that handedness is at least partially genetically determined and probably highly polygenic (i.e. as opposed to being the result of a single genetic variant or a small number of genetic variants (Annett 2013)). In the largest genome-wide association study (GWAS) of handedness to date, Cuellar-Partida et al. identified 48 genetic variants (single nucleotide polymorphisms; SNPs) associated with individual hand preference (Cuellar-Partida et al. 2021). Of these, 41 influenced a person’s likelihood of being left-handed/right-handed, while the other seven were associated with using both hands for tasks. In addition, the authors estimated that additive genetic factors explained ~ 12% (95% CI 7.20, 17.70) of the variance in the underlying liability of being left-handed (Cuellar-Partida et al. 2021) by examining genome-wide identity-by-descent sharing and handedness concordance in close relatives in the UK Biobank (Sudlow et al. 2015). Although this figure is slightly lower than estimates from the largest twin study of handedness to date (~ 25% heritability (95% CI 15.69, 29.51) (Medland et al. 2009), the evidence clearly shows that residual (including unshared environmental), rather than heritable genetic factors are responsible for most of the variation in handedness. Given that hand preference appears early in development, the corollary is that early life environmental exposures (i.e. in utero and shortly after birth) may be important determinants of handedness (Kovel et al. 2019).
One potential cause for a shift towards left-handedness is foetal hypoxia as a result of maternal smoking during pregnancy (Bakan 1991). Maternal smoking during pregnancy can produce foetal hypoxia in several ways: it can reduce oxygen supply to the foetus due to the production and binding of carbon monoxide (a constituent of tobacco smoke) to haemoglobin (El-Ardat et al. 2014); nicotine can reduce blood flow to the foetus due to vasoconstriction (El-Ardat et al. 2014), and/or increased blood viscosity (i.e. as a result of red blood cells increasing in size to compensate for reduced oxygen) (Shimada et al. 2011); and smoking can damage the placenta which can then reduce blood flow and therefore the amount of oxygen and nutrients delivered to the developing foetus (Suter and Aagaard 2020). Thus, because left-handedness is associated with gestational and birth complications (Strien et al. 1987) and many birth complications are a result of prenatal or perinatal hypoxia (Bakan 1991), foetal hypoxia due to maternal smoking may also increase the frequency of left-handedness.
Studies investigating the relationship between maternal smoking and offspring handedness are sparse and conflicting (Kovel et al. 2019; Bakan 1991; McManus 1981). In addition, observational studies are susceptible to bias and confounding and any reported associations may not reflect causal relationships between the phenotypes (Mann 2003). Mendelian randomization (MR) studies (Smith and Ebrahim 2003), in which genetic variants are used as instrumental variables (IVs) to proxy the exposure of interest, provide an alternative way to investigate potential causal relationships between traits (Smith and Ebrahim 2003; Sanderson et al. 2022). MR studies are, in theory, less prone to confounding compared to traditional observational epidemiological studies because genetic variants segregate randomly and assort independently of potential genetic and environmental confounders (Sanderson et al. 2022). However, MR relies on three key assumptions to produce valid results: 1. The genetic variants must associate with the risk factor of interest (Relevance assumption), 2. There must be no unmeasured confounders for the associations between genetic variants and outcomes (Independence assumption) and 3. The genetic variants must only affect the outcome through their effect on the risk factor of interest (Exclusion-restriction assumption).
In the present study we used data from the UK Biobank (UKB) to investigate a possible causal effect of maternal smoking during pregnancy on offspring handedness. We first examined the association between maternal smoking and other early life factors on offspring hand preference using multivariable logistic regression. We then used a proxy gene by environment (GxE) Mendelian randomization method (Spiller et al. 2019) where an individual’s own genotype for smoking heaviness (i.e. at the rs16966968 variant in the CHRNA5 gene) (Ware et al. 2011) was used to proxy the genotype of their mothers at the same locus, and therefore their mother’s smoking behaviour, to examine causality (Fig. 1). If the heaviness of maternal smoking increases the likelihood of left-handed offspring, then the association between rs16966968 and handedness should only be observed in individuals who reported that their mothers smoked during pregnancy. In contrast, the presence of an association between the variant and handedness in individuals whose mothers did not smoke during pregnancy would suggest the association is a result of confounding through genetic pleiotropy (Yang et al. 2019).Fig. 1. Proxy gene-by-environment Mendelian randomization conceptual framework. Offspring genotype is used as a proxy for maternal genotype and analyses are stratified on maternal smoking behaviour during pregnancy. Red crosses indicate closed pathways, for example, in b, maternal genotype for smoking heaviness did not influence smoking heaviness behaviour because the mothers did not smoke during pregnancy. Hence a red cross over the path from maternal genotype to maternal smoking heaviness. If heaviness of smoking in mothers is causal for offspring handedness, then an association between offspring genotype and offspring handedness should only be present in the children of mothers who smoked during pregnancy. The design assumes that offspring genotype only potentially associates with offspring handedness through maternal genotype and not through offspring phenotype. Since smoking initiation occurs well after hand preference manifests, offspring smoking behaviour should have no influence on offspring handedness (although this does not rule out the possibility of other pleiotropic actions on offspring handedness through the offspring genome). The design also assumes the absence of pleiotropic paths between maternal genotype and offspring handedness outside of maternal smoking behaviour. However, stratification on maternal smoking behaviour during pregnancy provides a test for the presence of horizontal pleiotropy in the maternal and/or offspring genomes. Specifically, if the above assumptions hold, the estimated causal effect of maternal smoking on offspring handedness in the offspring of mothers who did not smoke during pregnancy should be zero. If this is not the case, then the true causal effect of maternal smoking on offspring handedness (adjusted for pleiotropy) can be estimated by subtracting the causal estimate obtained in the offspring of non-smoking mothers from the causal estimate obtained in the offspring of smoking mothers. We also assume no paths from paternal genotype at the same (or correlated loci) to offspring handedness (not shown) and that the SNPs used are not associated with smoking initiation and/or termination (see the Discussion for further explication of these points)
Whilst the genotype by proxy GxE MR design provides a degree of robustness to horizontal pleiotropy, it suffers from low statistical power due to the reliance on offspring SNPs to proxy maternal genotype and because of the stratification into groups (smoking and non-smoking). We therefore extended the basic proxy GxE method to incorporate a polygenic risk score comprised of genome-wide significant SNPs for smoking heaviness (P ≤ 5 × 10^–8^) (PRS) (Evans et al. 2009; Evans et al. 2013). We reason that by extending the classic single SNP GxE design, the score may explain more of the phenotypic variance in maternal smoking heaviness, and therefore increase the power of our analysis. Additionally, the existence of a “no-relevance” control group (i.e., individuals who reported that their mothers did not smoke in pregnancy) should enable causal effect estimates to be corrected for the effect of horizontal pleiotropy.
Methods
Study Participants
The UKB is a community-based, prospective study ((https://www.ukbiobank.ac.uk/) Sudlow et al. 2015). Recruitment of ~ 500,000 participants and baseline assessments were completed between 2006 and 2010. Participants were aged 40–70 years at baseline, registered with a general practitioner and lived close to 22 assessment centres in England, Scotland, and Wales. Baseline assessments included demographics, lifestyle, and disease history, with linkages to electronic medical records. UK Biobank’s ethical approval was from the Northwest Multi-centre Research Ethics Committee.
We excluded individuals of non-white European ancestry (n = 59,910) and those who had withdrawn their consent (n = 52). We also excluded participants who reported a birthweight heavier than 6.0 kg (n = 236) and lighter than 1 kg (n = 1013) as, although some reports may have been accurate, the others would likely be self-report errors. This cut-off aimed to reduce the effects of outliers and measurement error in the model fitting. Participants were asked “Are you right or left-handed?" during a baseline questionnaire and self-reported their handedness (right-handed = 0, left-handed = 1 or uses both right and left hands equally = 2). Individuals who did not report their hand preference (n = 92) were removed, as were individuals who reported using both hands equally (n = 7105) because of consistencies across time points (Kovel et al. 2019), and because it is questionable the degree to which this simple self-report measure in UK Biobank reflects true ambidexterity (i.e. the ability to perform tasks equally well with either hand). We therefore had 434,104 individuals available for our study (see Fig. 1 of the Supplement for flowchart of exclusions).
Traditional Observational Epidemiological Analyses
Potential Early Life Predictors of Handedness
Exposure variables examined for being potential early life predictors of handedness were as follows: sex, being part of a multiple birth, maternal smoking while pregnant, if the participant was breastfed, country of origin (England, Wales, Scotland, Republic of Ireland, Northern Ireland or Elsewhere), year of birth, birth weight, month of birth and social deprivation. We modelled the month of birth as a cosine function in order to represent a continuous seasonal effect (i.e. months next to each other are more similar than other months) with peaks in the UK summer and winter (see (Kovel et al. 2019) for more details). Social deprivation was modelled using the Townsend score (area-level deprivation) and was derived from data on unemployment, car ownership, household overcrowding and owner occupation aggregated at postcode area (Townsend et al. 1988). Townsend scores were assigned to participants based on their address at recruitment and were calculated immediately prior to recruitment using data from the preceding national census data (2001). Higher Townsend scores equate to higher levels of socioeconomic deprivation. All the variables were self-reported (other than sex and date of birth which were acquired from the central registry at recruitment). Responses of “do not know” and “prefer not to answer” were treated as missing values. See Supplementary Table 1 for the variables included in the analysis and how each variable was coded.
Statistical Analysis
We performed univariable logistic regressions between each of the variables and hand preference. We then performed a multivariable analysis of hand preference, where all variables were included in the model, adjusted for the first ten genome-wide principal components based on the genome-wide SNP data. In this model, participants with missing values for any of the predictor variables were removed (final sample: n = 193,770, 37.93% male). The skew in the sex ratio is a result of males under reporting their birthweight (20.18% increase in missingness). We ran the multivariable analyses on the whole sample and in sex stratified samples. We also computed the McFadden pseudo R^2^ to assess how well the adjusted model explains the variation in the outcome.
Finally, we investigated the pairwise relationships between predictor variables and hand preference. We used Cramer’s V for categorical pairs, Pearson’s correlation coefficient for continuous variables, and Spearman’s rho when a pair comprised one categorical variable and one continuous variable. When examining the correlation between country of birth and the other variables, we looked at each location individually.
Proxy Gene by Environment Mendelian Randomization
Genotyping and Quality Control
DNA was extracted from stored blood samples that had been collected from participants on their first visit to a UKB assessment centre. Genotyping was carried out using the UK Biobank Axiom Array by Affymetrix Research Services Laboratory in 106 sequential batches of approximately 4,700 samples. This resulted in a set of genotype calls for 489,212 samples and approximately 850,000 variants were directly measured. The UKB performed imputation centrally using the Haplotype Reference Consortium (McCarthy et al. 2016), UK10K (Huang et al. 2015), and 1000 Genomes project reference panels (Bycroft et al. 2018). Genotype data were screened for genotyping quality (described elsewhere (Bycroft et al. 2018)), Hardy–Weinberg equilibrium failure (p < 5 × 10^–8^) and minor allele frequency (MAF < 0.01). SNPs used in our analysis were high-quality HRC-imputed dosage data provided by the UKB full release (IMPUTE4 INFO score ≥ 0.976) and extracted using plink 2.0 (Chang et al. 2015).
Participants were excluded following the same criteria as in the observational univariable logistic regression analysis. We also removed samples that had sex chromosome aneuploidy, a mismatch between genetically inferred sex and self-reported sex, high genotype missingness and samples that were excluded from kinship inference and autosomal phasing. Further, we removed individuals who had extreme heterozygosity as this can be an indication of poor-quality genotyping, sample contamination etc. One individual from each pair of participants whose genetic relatedness was inferred to be 3rd degree or closer were also excluded. This was done based on genotype data. This left a sample of 346,871 individuals (see Fig. 2 of the Supplement for a flowchart of exclusions).
Smoking Phenotypes
At baseline assessment, participants were asked the question ‘Did your mother smoke regularly around the time when you were born?’. This was used to index maternal smoking during pregnancy.
Outcomes in Participants
Handedness of participants was reported at baseline. As in the traditional observational epidemiological analysis, due to inconsistencies in the reporting of ambidexterity, only individuals who reported being left- or right-handed were retained.
Single SNP Genetic Association and Instrumental Variable Analyses
We used individuals’ genotype at rs16969968, which is robustly associated with smoking heaviness (measured as cigarettes per day) (Ware et al. 2011), as a proxy for maternal genotype at the same locus, and consequently heaviness of maternal smoking. The SNP was coded as the number of smoking heaviness-increasing alleles (allele A).
Polygenic Risk Score Analyses
We used GWAS summary statistics for smoking heaviness generated from a meta-analysis of 60 genome wide association studies of European ancestry (UKB and 23andMe participants removed)(Saunders et al. 2022). 12 Independent SNP signals were identified using PLINK (r^2^ = 0.001, clump-kb = 1000). We created a genome-wide significant weighted polygenic risk score (PRS) of smoking heaviness for each individual in our sample (UKB) and used this as a proxy for the mothers’ PRS scores. Individuals were scored on the total number of smoking increasing alleles they carried across all genome-wide significant clumped variants weighted by regression coefficients from the GWAS of smoking heaviness (i.e., a weighted PRS). See Supplementary Table 2 for the variants used to construct the PRS, their closest gene and putative function.
Statistical Analysis
First, we investigated whether the SNP rs16969968 was associated with the likelihood of the mother smoking during pregnancy using logistic regression. Second, we stratified the mothers by their smoking status during pregnancy and tested the association between rs16969968 and handedness in each group using univariable regression and multivariable regression adjusting for year of birth, sex and the first ten genetic principal components. We repeated this analysis stratified by sex. Given that the participant’s genotype should not affect hand preference through their own smoking behaviour, (i.e., assuming hand preference long precedes smoking initiation), we did not account for the participant’s own smoking status in analyses.
We replicated this analysis using our PRS of smoking heaviness. As in the single-SNP analysis, we first investigated whether the PRS was associated with mother’s reported smoking status during pregnancy. We then stratified on mother’s reported smoking status during pregnancy and ran a univariable regression and a multivariable regression accounting for year of birth, sex and the first ten principal components.
We conducted follow-up single SNP analyses using Regenie (Mbatchou et al. 2021) which can account for population structure and cryptic relatedness across participants. The methods and results for these analyses are presented in the Supplement and did not differ substantively from the main results.
As a positive control and to demonstrate proof of principle for this approach, we separately regressed birthweight on the SNP rs16969968 and the PRS of smoking heaviness. This is because it is generally accepted that maternal smoking in pregnancy leads to lower offspring birthweight (Zheng et al. 2016). Therefore, if the SNP/PRS is a true proxy for maternal smoking behaviour during pregnancy we should see a negative association with offspring birthweight in offspring of mothers who smoked during pregnancy. To investigate the exchangeability assumption, we regressed the other predictors of hand preference (sex, birth year, being part of a multiple birth, being breastfed, birth month, social deprivation, and country of origin) on rs16969968 and the PRS. Under the exchangeability assumption, these phenotypes should not be associated with the SNP or the PRS. In both the positive and negative control analyses, we included the first ten genome-wide genetic principal components because its plausible that the frequency of the variants and the rates of the phenotypes may vary across ancestries.
We estimated the causal effect of maternal smoking heaviness during pregnancy on offspring hand preference using the Wald Ratio. We did this separately for both rs16969968 and the PRS. For the denominator of the Wald ratio, we calculated the association between rs16969968/PRS and own smoking heaviness (self-reported cigarettes per day) among smokers in the UK Biobank.
Results
Early Life Exposures of Handedness
The distribution of handedness in the whole cohort for the logistic regression analysis and the genetic subset is presented in Table 1. The distribution of characteristics in the whole cohort for the logistic regression analysis and the genetic subset is presented in Table 2. The distributions of characteristics in the retained cohort after excluding participants who have any missing values is presented in Supplementary Table 3. The pairwise relationships between predictor variables are presented in Supplementary Table 4.Table 1. Distribution of responses to question about hand preferenceHand useMales (%)Females (%)Total (%)Logistic regression analysis Right-handed175,965 (89.09)215,757 (91.19)391,722 (90.23) Left-handed21,546 (10.91)20,836 (8.81)42,382 (9.76) Total197,511236,593434,104Genetic subset Right-handed142,134 (89.02)170,665 (91.16)312799 (90.18) Left-handed17,523 (10.98)16,549 (8.84)34,072 (9.82) Total159,657187,214346,871Table 2Distribution of observed characteristics at baseline assessment in UK BiobankVariableCategoriesN (mean for continuous variables and % for categorical variables)Frequency of left-hand preference (%)Logistic regression Analysis SexFemale236,593 (54.50)8.81Male197,511 (45.50)10.91 Part of multiple birthsNo418,068 (97.75)9.71Yes9602 (2.25)11.59 Maternal smoking during pregnancyNo258,003 (69.17)9.77Yes114,976 (30.83)9.62 BreastfedNo94,905 (28.80)10.42Yes234,753 (71.21)9.47 Birthweight247,222 (3.33 kg) Birth Year434,104 (1951) Age at assessment centre434,104 (56.85) Birth Month434,104 (June) Social deprivation index^a^433,586 (− 1.53) Country of OriginEngland365,661 (84.24)10.08Wales20,804(4.79)7.33Scotland37,246 (8.58)8.12Northern Ireland2113 (0.49)8.80Republic of Ireland253 (0.06)8.30Elsewhere7966 (1.84)9.50Genetic Subset Maternal smoking during pregnancyNo206,424 (69.37)9.84Yes91,158 (30.63)9.63 Number of smoking heaviness increasing alleles0155,732 (44.89)9.881153,448 (44.23)9.85237,763 (10.88)9.48 SexMale159,701 (46.03)10.98Female187,242 (53.97)8.84 Year of birth346,943 (1951) Social deprivation346,943 (-1.56)^a^Townsend deprivation index(z-score) where higher numbers denote higher levels of socioeconomic deprivationNB: Maternal smoking during pregnancy was reported by offspring
Traditional Observational Epidemiological Analyses
There was evidence that a later year of birth, being male, being from a more deprived area, being part of a multiple birth, having a lower BW and not being breastfed all increased the likelihood of being left-handed. Further, being born in the summer/winter months and being born in England (as opposed to Scotland or Wales) also increased the likelihood of being left-handed. (Fig. 2).Fig. 2. Association between early-life predictors of handedness and left-hand preference. An odds ratio > 1 indicates propensity to be left-handed. Unadjusted refers to results from univariable logistic regression analyses. Adjusted refers to results from multivariable logistic regression analyses. NB. Variable “Mother smoked when pregnant” was reported by offspring
Results from the multivariable analysis were similar to results from the univariable analysis. However, having a mother who smoked during pregnancy increased the likelihood of being right-handed after accounting for covariates. Results when stratised to proxy maternal smoking were related to the number of cigarettes smoked per day. Additionally, there was evidence of an association between the SNP and being part of a multiple birth when conditioning on maternal smoking during pregnancy (Supplementary Table 6). This is likely to be an effect of collider bias and is examined in greater detail in the discussion.
In the unadjusted models and models adjusted for year of birth, sex and the first ten principal components, we found no strong association between rs16969968 and offspring hand preference in the offspring of mothers who smoked during pregnancy or in the offspring of mothers who did not smoke during pregnancy. Sex stratified results were similar to the whole cohort analysis. Results from model 1 (unadjusted) are presented in Table 3 and model 2 (adjusted for year of birth, sex and the first ten genetic principal components) are presented in Fig. 3.Table 3. Genetic association between rs16969968 and left handedness in the unadjusted modelOR95% CIpWhole sampleNon-smokers0.9830.963, 1.000.083Smokers0.9940.968, 1.020.682Females onlyNon-smokers0.9750.944, 1.010.110Smokers0.9940.946, 1.040.818Males onlyNon-smokers0.9780.947, 1.010.167Smokers0.9830.937, 1.030.459NB: The odds ratio refers to being left-handed and the effect of the “A” allele at rs16969968Mothers smoking was reported by offspringFig. 3Odds ratio for offspring being left-handed per A allele at rs16969968. NB. Genetic association results show decreased odds of offspring being left-handed per additional allele A at rs16969968, adjusted for year of birth, sex and the first 10 principal components (N = 346,943). Maternal smoking during pregnancy was reported by offspring
We then estimated the causal effect of maternal smoking during pregnancy on offspring hand preference using the Wald Ratio. We estimated the causal estimate in offspring who reported that their mothers smoked during pregnancy, as well as offspring who reported that their mothers did not smoke during pregnancy. Under certain assumptions (e.g., the absence of collider bias, that maternal smoking does not affect offspring handedness pre-conception and postnatally etc.) the Wald ratio in this latter group should provide an estimate of bias due to pleiotropy, since any effect of the SNP on hand preference in offspring cannot be due to maternal smoking behaviour during pregnancy. We therefore calculated the estimated causal effect of maternal smoking during pregnancy on offspring handedness accounting for pleiotropy by subtracting the estimated causal effects in the offspring of mothers who did and did not smoke during pregnancy. Again, we fit two models: model 1 which provided unadjusted estimates and model 2, where the SNP-exposure and SNP-outcome associations were adjusted for sex, year of birth and the first ten principal components. Results are presented in Table 4.Table 4. Estimated causal effect of maternal smoking heaviness during pregnancy on offspring left-hand preferenceCausal effect of genetically predicted smoking heaviness on offspring left-hand preference in mothers who smoked during pregnancyCausal effect of genetically predicted smoking heaviness on offspring left-hand preference in mothers who did not smoke during pregnancyCausal effect of genetically predicted smoking heaviness on left-hand preference accounting for pleiotropyβ_IV_95% CIβ_IV_95% CIβ_IV_95% CIrs16969968 model 1− 0.0111− 0.0641, 0.0420− 0.0337− 0.0740, 0.004550.0237− 0.0423, 0.0897rs16969968 model 2− 0.0135− 0.0650, 0.0381− 0.0351− 0.0732, 0.003110.0216− 0.0426, 0.0857PRS model 1− 0.0113− 0.0604, 0.0378− 0.0246− 0.0611, 0.01180.0113− 0.0498, 0.0725PRS model 2− 0.0133− 0.0625, 0.0360− 0.0247− 0.0597, 0.01030.0114− 0.0490, 0.0719NB: Results show the log odds of offspring left handedness per cigarette smoked per day during pregnancy in mothers as calculated using the Wald estimator. Model 1 refers to the Wald estimator calculated without adjustment for covariates. Model 2 refers to the Wald estimator adjusting for sex, year of birth and the first 10 principal components (in the regression coefficients in both the numerator and denominator).Maternal smoking during pregnancy was reported by offspring (Table 6).
The PRS was slightly more predictive of own cigarettes smoked per day among self-reported smokers as reported in the UKB, compared to the single SNP rs16969968 (R^2^ = 0.005 vs 0.004). As in the single SNP analyses, we found a positive association between the PRS and maternal smoking during pregnancy and a negative association between the PRS and birthweight, our positive control (Supplementary Table 6). We also found associations between the PRS and other phenotypes, such as social deprivation (Supplementary Table 6). Such associations have been reported previously (Reed et al. 2025) and may reflect upstream effects captured in the original smoking GWAS and/or horizontal pleiotropy. A positive association between the smoking heaviness variants and year of birth appeared when stratifying by maternal smoking status, however, this is likely to be a collider effect (see Discussion).
We found no strong association between the PRS and offspring handedness in mothers who smoked during pregnancy or in mothers who did not smoke during pregnancy (Table 5). We found the PRS was negatively associated with birthweight, and this was only in mothers that smoked during pregnancy (Table 6). As in the single-SNP analysis, we used the Wald Ratio to estimate the causal effect of maternal smoking during pregnancy on offspring hand preference and to estimate the causal estimate in mothers that did not smoke during pregnancy (the pleiotropic effect). We were then able to estimate the causal effect of the maternal smoking during pregnancy on offspring hand preference, accounting for pleiotropy. We found there was no strong causal effect of maternal smoking during pregnancy on offspring hand preference in either subgroup. Results are presented in Table 4.Table 5. Genetic association between the PRS for smoking heaviness and left handednessGWAS phenotypeUnadjustedAdjusted for year of birth, sex and first 10 principal componentsp-value thresholdN SNPsOR95% CIPOR95% CIpSmokers during pregnancy5 × 10^–8^120.8950.412, 1.3780.6520.8730.389, 1.360.581Non-smokers during pregnancy5 × 10^–8^120.7850.427, 1.140.1850.7760.417, 1.1350.166Table 6Genetic association between genome-wide significant SNPs for smoking heaviness and birthweight in the GxE MR analysisBeta95% CIPBeta95% CIpSmokers during pregnancy5 × 10^–8^12− 0.208− 0.339, − 0.0760.002− 0.189− 0.318, − 0.0590.004Non-smokers during pregnancy5 × 10^–8^12− 0.028− 0.117, 0.0610.533− 0.016− 0.104, 0.0720.715NB: Maternal smoking during pregnancy was reported by offspring
Discussion
In this study, we investigated the relationship between early life exposures and handedness with a specific focus on maternal smoking during pregnancy. Our logistic regression analyses suggested that the odds of being left-handed were higher in males, those that were part of a multiple birth, had a lower birthweight or were bottle fed rather than breastfed. Additionally, social deprivation, being born more recently, being born in the summer/winter months or being born in England (as opposed to those born elsewhere) also increased the likelihood of being left-handed. These findings were maintained after controlling for the other predictors of handedness in the multivariable logistic regression analyses and are consistent with previous investigations in the UK Biobank (Kovel et al. 2019). However, we also found evidence to suggest that, after adjusting for covariates, and contrary to expectation, maternal smoking during pregnancy was associated with an increased likelihood of offspring being right-handed.
The unexpected result that maternal smoking during pregnancy is associated with an increased likelihood of offspring being right-handed, may reflect collider bias. Specifically, maternal smoking during pregnancy is associated with some of the variables we conditioned on in the multivariable model and is likely to be causal for some of these too e.g., birthweight (Tyrrell et al. 2012). If one or more unmeasured confounders were also causal for e.g., both birthweight and hand preference, then conditioning on birthweight in the multivariable model could generate a spurious association between maternal smoking during pregnancy and offspring hand preference. Indeed, when introducing birthweight into the multivariable model, the association between maternal smoking during pregnancy and offspring hand-preference became stronger which is consistent with this hypothesis. This potential effect of collider bias is illustrated in Fig. 4.Fig. 4. Graphic to demonstrate potential collider bias in the traditional observation epidemiological analysis as a result of the inclusion of unmeasured confounders and factors conditioned on, such as birthweight. Diagram illustrating the potential issue of collider bias in the traditional observational epidemiological analysis. The issue will arise if we condition on factors that are associated with both the exposure (Maternal smoking during pregnancy) and unmeasured confounders (such as birthweight). This is because if there are unmeasured confounders that cause birthweight and are also associated with offspring hand preference, this will create a pathway between maternal smoking during pregnancy and offspring hand preference. This is because birthweight becomes a collider. NB. Maternal smoking during pregnancy was reported by offspring
We also investigated the association between maternal smoking during pregnancy and offspring handedness using genetic association analyses and a proxy GxE MR design (Yang et al. 2019). These analyses suggested no strong causal effect of maternal smoking on offspring handedness, despite similar analyses showing a causal effect of maternal smoking on offspring birthweight (i.e. our positive control analysis). However, statistical power is limited in these designs for a number of reasons, complicating the interpretation of null results.
First, genetic variants typically explain only a small proportion of the phenotypic variance in the exposure variable, and consequently most MR studies require extremely large sample sizes to detect modest causal effects (Brion et al. 2013). Further, proxy GxE MR studies use offspring variants as a surrogate for maternal genotype. As offspring variants are only expected to explain ~ 25% of the variance in the maternal exposure compared to the same genotypes in the mother, power is reduced even further in this type of design. Third, proxy GxE MR studies require some sort of sample stratification for informative causal inference (i.e. maternal smoking status during pregnancy in the present study). This further reduces sample size and statistical power to detect a causal effect (particularly as mothers who smoked during pregnancy are a minority in the UK Biobank). Whilst it would be more statistically powerful to examine the relationship between directly genotyped mothers and their phenotyped offspring (simultaneously controlling for offspring genotype), this is only practically feasible in the very few cohorts that contain very large numbers of genotyped mother–offspring pairs (Evans et al. 2019; Moen et al. 2020; Moen et al. 2019; D’Urso et al. 2024). For now, the proxy MR GxE design remains a valuable and practical addition to the expanding range of epidemiological and statistical methods aimed at enhancing the power to identify causal effects of maternal exposures on offspring outcomes (Hwang et al. 2020).
We performed a number of data simulations to investigate the power of the proxy MR GxE design to detect causal effects of maternal smoking on offspring handedness (Supplementary Note and Supplementary Table 7). Our results suggest that whilst a cohort the size of the UK Biobank would be well powered to detect strong effects of maternal smoking on offspring risk of left-handedness (e.g. a one standard deviation increase in number of cigarettes smoked per day during pregnancy increasing risk of offspring left handedness by ~ 44%), much larger sample sizes would be required to detect weaker causal effects. Since most currently available cohorts with genome-wide SNP data are smaller than the UK Biobank (with the exception of the 23andME data resource), it seems like meta-analysis across different studies would seem like a sensible way to improve statistical power, much like it has in GWAS.
One strategy we employed to increase the power of our MR analyses was to use a smoking-related PRS that explained more of the variance in maternal smoking heaviness. Although it is highly likely that at least some of the SNPs that comprise this score were pleiotropic (see Supplementary Tables 2 and 6 and (Reed et al. 2025)), we reasoned that an unbiased estimate of the causal effect might still be obtained. We theorised this could be done by subtracting the estimated causal effect obtained in individuals whose mothers did not smoke during pregnancy from the causal effect obtained in the offspring of mothers who did smoke in pregnancy. However, one potential complication is that this strategy assumes that the variants used to proxy the exposure (smoking heaviness) are unrelated to smoking initiation or cessation (ever/never smoked). Our analyses showed that both the rs16969968 variant and the smoking heaviness PRS were both associated with the likelihood of mothers’ smoking during pregnancy-implying a relationship between the variants and ability to quit smoking. Additionally, some of the genetic variants have also been previously associated with smoking initiation (Supplementary Table 2). As we illustrate in Fig. 5, associations between genetic variants and smoking initiation/termination could result in collider bias when conditioning/stratifying on maternal smoking status during pregnancy. “Correcting” the causal estimates for pleiotropy (by the procedure outlined above) will not obviate this problem. The corollary is that our MR estimates are likely to have been biased through a collider effect.Fig. 5. Graphic to demonstrate potential collider bias in Gene by Environment Mendelian randomization analyses. Diagram illustrating the potential issue of collider bias in the gene x environment Mendelian randomisation (GxE MR) analysis. The issue will arise if SNPs that are related to smoking initiation/termination are used as instruments in the analysis. In the GxE model, we condition on smoking status during pregnancy. If there are additional confounders (C) that cause both smoking initiation/termination (ever/never smoked) and offspring hand preference, this will create a pathway between the SNP and offspring hand preference when SNPs included in the model are associated with smoking initiation/termination as well as maternal smoking heaviness. This is because smoking initiation/termination becomes a collider variable
There are some further limitations to our study. First, although the UKB is a prospective cohort, most of our variables were based on retrospective recall and self-report. Therefore, individuals may have been misclassified: e.g. participants may misreport if their mother smoked during pregnancy, especially due to the negative connotations surrounding such behaviour. We would expect that such misclassification might attenuate any association between maternal smoking and offspring handedness. Also, any misclassification will make the MR results from each group in the stratified analyses more similar and consequently reduce the effectiveness of any correction for pleiotropy.
Second, our analyses assume no relationship between the paternal genotype at rs16969968/GRS and offspring hand preference. However, any effect of paternal smoking on the foetus is likely to be far weaker than any maternal effect. Given that we found no effect of maternal smoking behaviour on offspring hand preference, we do not think that unmodelled paternal effects are a likely cause for concern.
Third we assume no assortative mating on smoking behaviour. It is well known that married couples tend to have more similar smoking habits than would be expected by chance (Homish and Leonard 2005). Assortative mating induces complex patterns of correlations between smoking related loci between and within individuals (Lynch and Walsh 1998). The consequence is that phenotypic assortment can result in a myriad of possible paths between an ostensible instrumental variable and the outcome. Within family MR can generate causal estimates that are robust to assortative mating (Brumpton et al. 2020; Davies et al. 2019; Hwang, et al. 2021). However, the GxE proxy MR design is not and as such, parental genotypes and behaviours may not be fully independent. Future work using mother-father-offspring genotype data could be used to explicitly model the parental genotype correlations and further refine the analysis. However, only in certain large cohorts is this possible (Evans et al. 2019; Moen et al. 2020) and we believe our approach (removing related individuals and correcting for population stratification) adequately reduces the potential bias.
Fourth, we recognise that triangulation of results using different methods and samples would be beneficial to increase confidence in our results. We aimed to do this using a combination of classic epidemiological and MR methods, given that they make different assumptions and are subject to different biases. However, the use of additional methods and/or datasets in the future would increase confidence in the current conclusions.
Future research investigating the aetiology of handedness could focus on other predictors of hand-preference such breast-feeding which showed association in the present study and in previous investigations (Denny 2012). The release of exome sequencing in UKB has enabled the detection of rare variants that contribute to variation in human hand preference (Schijven et al. 2024). The identification of other rare mutations that influence hand preference may be possible through the use of whole genome sequencing data which is also now available through the UKB.
In conclusion, this study contributes to the understanding of handedness and its potential early-life determinants. Our findings using the UK Biobank cohort align with previous findings and emphasise the impact of factors such as birth year, birthweight, being part of a multiple birth and breastfeeding on hand preference. The results using the cohort data diverged from previous findings in that they suggested that maternal smoking during pregnancy may be associated with an increased likelihood of right-handedness rather than left-handedness. This unexpected finding, however, needs to be interpreted cautiously, considering the potential influence of collider bias. Additionally, when further investigating this association using a proxy GxE MR design, we found no strong evidence for a causal link between maternal smoking and handedness. However, it is unclear whether this is a true null effect or a consequence of limited power. Meta-analyses incorporating similar data from other large-scale biobanks may help answer this question.
Supplementary Information
Below is the link to the electronic supplementary material.Supplementary file1 (DOCX 2281 KB)
The reference list from the paper itself. Each links out to its DOI / PubMed record.