Metagenomics-Metabolomics Reveals the Alleviation of Indole-3-Ethanol on Radiation-Induced Enteritis in Mice
Hua Zhong, Yipeng Song, Shanliang Hu, Chengxu Zhang, Lili Li

TL;DR
This study shows that indole-3-ethanol reduces radiation-induced gut inflammation in mice by improving gut health and reducing inflammation.
Contribution
The novel finding is that IEt alleviates radiation enteritis through Roseburia and anti-inflammatory metabolites like indole-3-carbinol.
Findings
IEt reduced pro-inflammatory factors like IL-17A and improved colonic structure in irradiated mice.
IEt enriched Roseburia, an anti-inflammatory gut bacteria, and increased beneficial metabolites like indole-3-carbinol.
Roseburia was positively correlated with indole-3-carbinol and negatively with IL-17A, suggesting a protective mechanism.
Abstract
Indole-3-ethanol (IEt), a small molecule metabolite from intestinal microbial tryptophan metabolism, has been established to have anti-inflammatory properties. However, its effect on radiation-induced enteritis has not been reported. Here, we aim to explore the effects and potential mechanisms of IEt on radiation enteritis. C57BL/6J mice were orally administered an IEt solution before radiation exposure. Inflammatory factors, including IL-17A, IFN-γ, IL-6 and IL-1β, were detected using enzyme-linked immunosorbent assay. Colonic histopathology was assessed through H&E staining. Subsequently, gut microbiota and its metabolites were analyzed using metagenomics and metabolomics. The results suggested that IEt alleviated radiation-induced enteritis, as evidenced by improved colonic structural integrity, decreased levels of pro-inflammatory factors like IL-17A, and the restoration of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
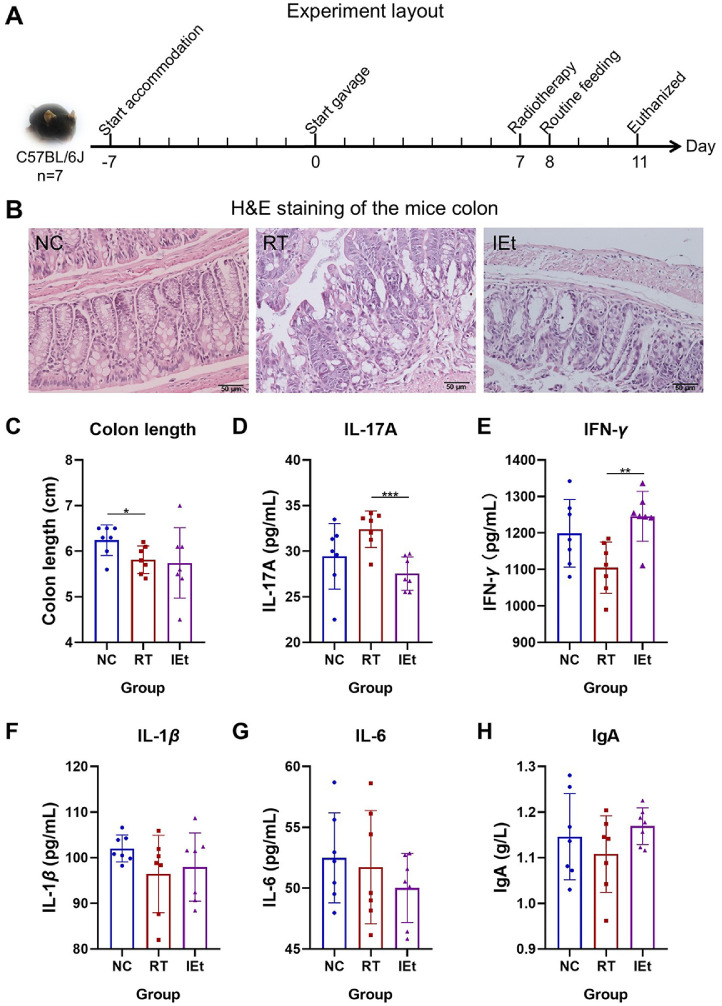 Figure 1
Figure 1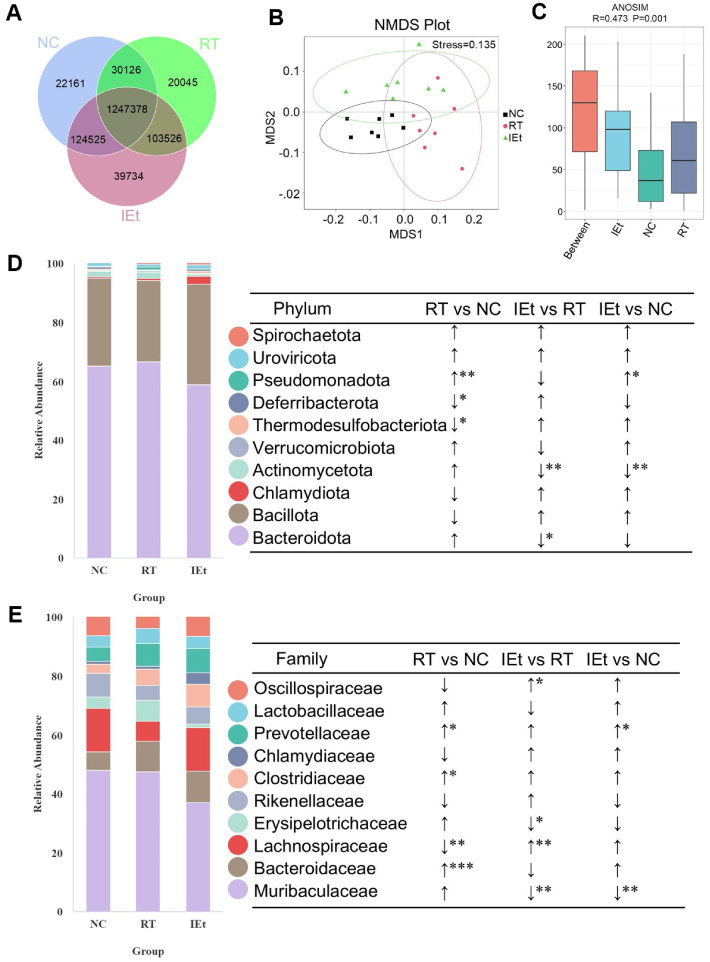 Figure 2
Figure 2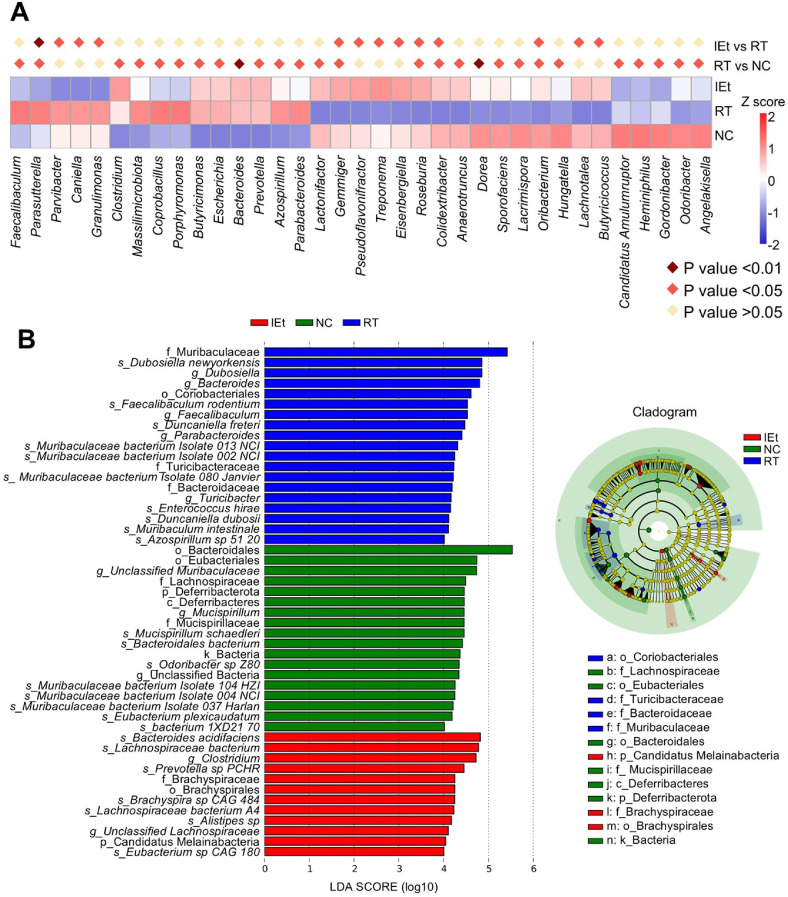 Figure 3
Figure 3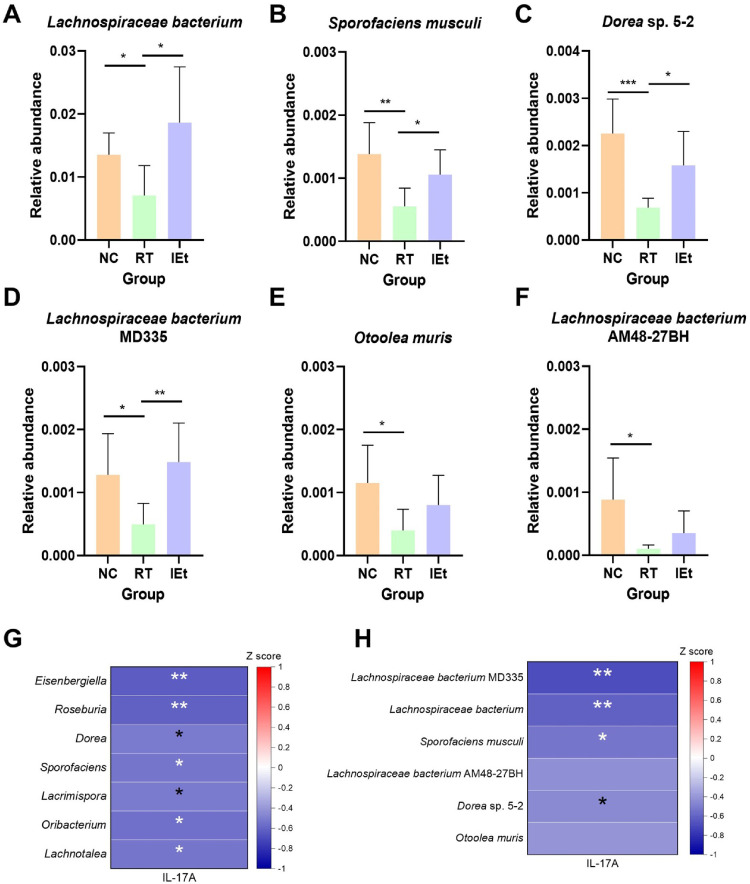 Figure 4
Figure 4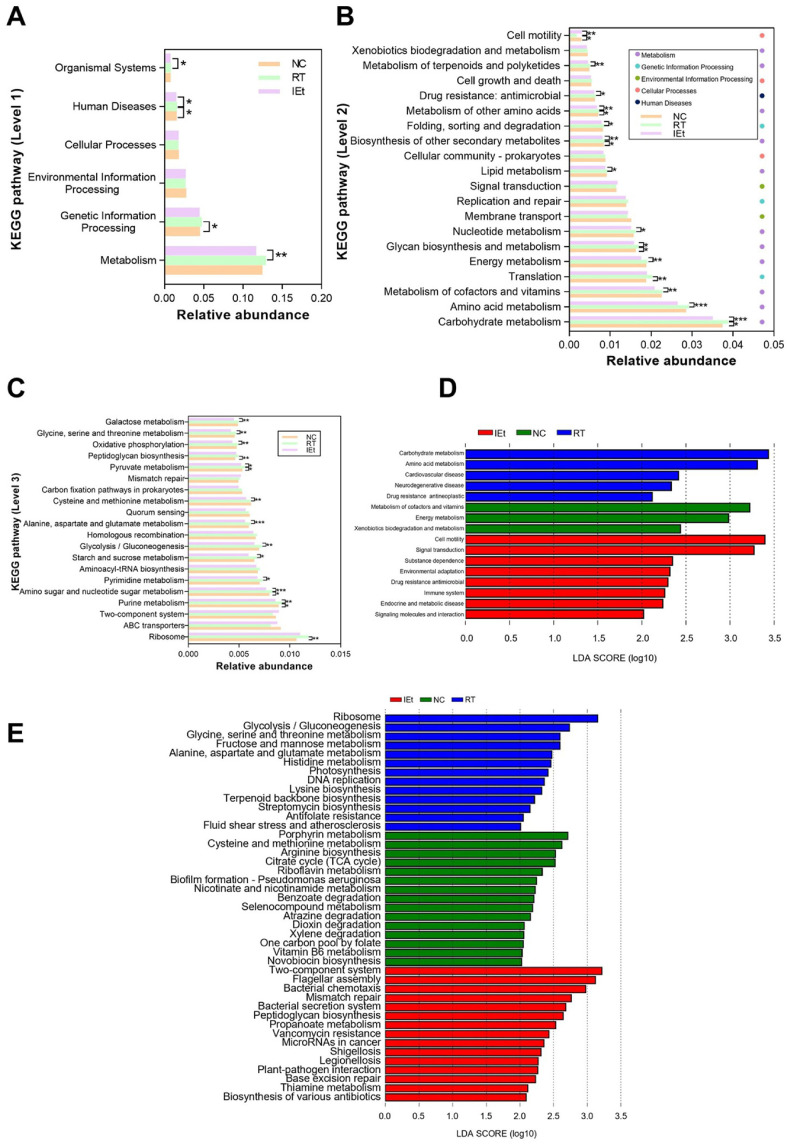 Figure 5
Figure 5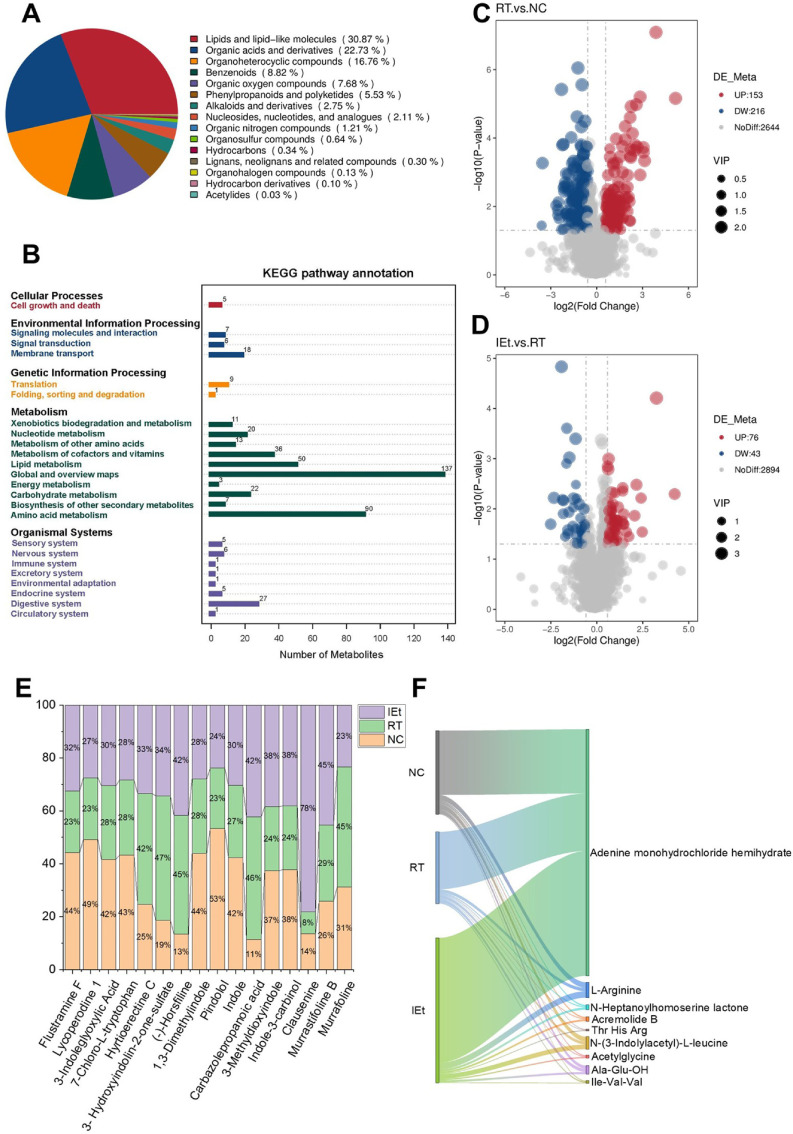 Figure 6
Figure 6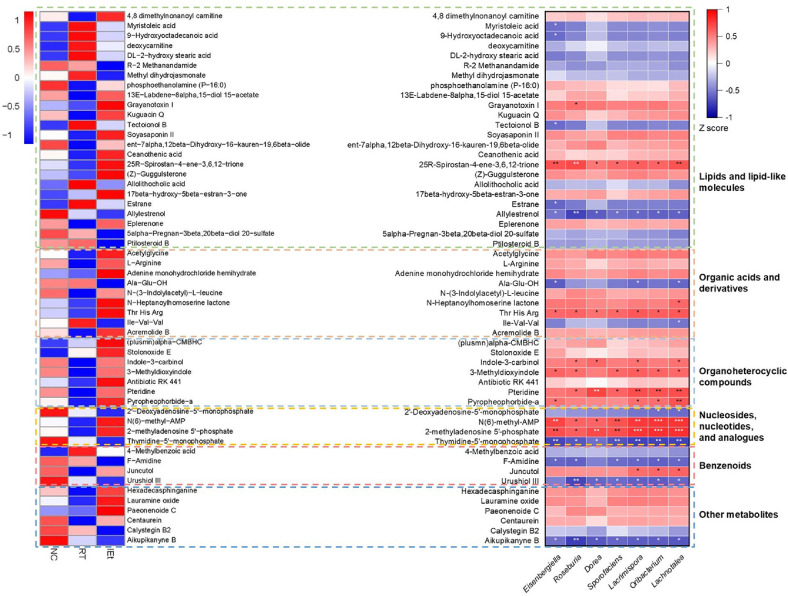 Figure 7
Figure 7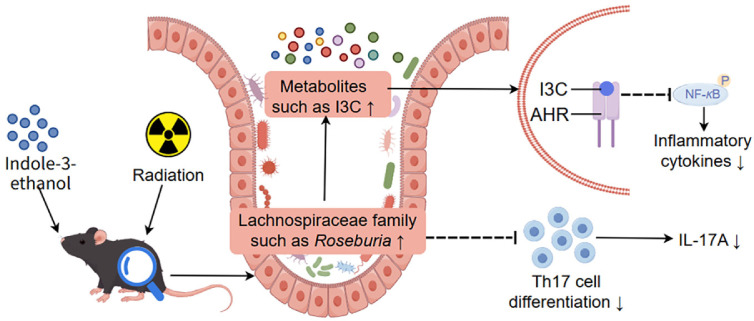 Figure 8
Figure 8Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGut microbiota and health · Effects of Radiation Exposure · Genomics, phytochemicals, and oxidative stress
Introduction
Cancer remains a major global health burden, with the International Agency for Research on Cancer estimating approximately 19.974 million new cancer cases worldwide in 2022 [1]. Radiotherapy, a cornerstone of cancer therapy, is utilized by over 50% of patients [2, 3]. However, when applied to pelvic tumors, radiation exposure often affects adjacent intestinal tissues due to their anatomical proximity, significantly increasing the risk of radiation enteritis (RE) [4]. Acute RE manifests in 60%–80% of patients within three months post-treatment, while 5–20%may progress to chronic cases [5, 6]. For intra-abdominal and pelvic malignancies, the irradiated field is often close to the intestine, resulting in radiation-induced damage that disrupts intestinal microecology and can lead to radiation enteritis [7]. The occurrence of radiation enteritis can lead to weight loss and a compromised immune response in patients receiving radiotherapy, potentially leading to treatment interruptions [8].
Although the complete etiology of radiation enteritis is not fully understood, emerging evidence indicates that the gut microbiome plays a crucial role in the onset of this condition. Radiotherapy may alter the diversity of intestinal microbiota and reduce the abundance of beneficial bacteria [4]. These alterations in the microbial community may increase intestinal permeability, allowing substances like lipopolysaccharide that should not normally pass through the intestinal wall to enter the body, provoking an immune response and consequently heightening inflammation [9, 10]. Furthermore, the long-term surviving mice after radiation exhibited a unique gut microbiota enriched in the family Lachnospiraceae compared to the control group [11]. Fecal microbiota transplantation (FMT) experiments revealed that germ-free mice receiving irradiated donor microbiota developed more severe intestinal mucosal damage and higher inflammation scores than those receiving control microbiota [12]. Therefore, gut microbiota is crucial in the development of radiation enteritis.
In addition, gut microbiota-derived metabolites also play an important role in radiation enteritis. Indole-3-carboxaldehyde, a crucial tryptophan metabolite, has been shown to increase the levels of beneficial bacteria and stimulated the AhR/IL-10/Wnt pathway, thereby fostering the growth of intestinal epithelial cells in irradiated mice [13]. Moreover, another study reported that certain intestinal microbial metabolites such as short-chain fatty acids (SCFAs) and derivatives of tryptophan like indole-3-carboxaldehyde and kynurenic acid serve as radioprotective agents [11]. It has been reported that intestinal microbial metabolites such as indole-3-propionic acid, can mediate some effects induced by FMT, thereby playing a protective role against gastrointestinal toxicity [14]. These findings suggest that gut microbial metabolites protect against the adverse side effects of radiation exposure. Although microbial metabolites are known to modulate host physiology in health and disease [15, 16], the functional roles and mechanistic underpinnings of most gut-derived small molecules remain poorly characterized. Indole-3-ethanol (IEt), a small molecule metabolite produced by intestinal microorganisms, is also an intermediate in the tryptophan metabolic pathway. Its effects on radiation-induced enteritis have not been reported. Based on the close relationship between radiation enteritis and intestinal microecological disorder, this study explored the effects and potential mechanisms of IEt on radiation-induced enteritis in mice through integrated metagenomics and metabolomics analysis of gut microbiota, providing theoretical support for the prevention of radiation enteritis.
Materials and Methods
Animals and Experimental Design
Six-week-old female C57BL/6J mice, obtained from Jinan Pengyue Experimental Animal Technology Co., Ltd.,(China), were housed in a specific pathogen free environment, maintained at an ambient temperature ranging from 22°C–25°C, with 50%–55% humidity, and a lighting cycle of 12 h of light followed by 12 h of darkness. The mice were provided with sterile water and standard chow (Research Diets D12450B, HFK Bioscience Co., Ltd., China). Table S1 detailed the components of the standard chow. After a 7-day acclimation phase (Fig. 1A), the mice were randomly assigned to three groups (n = 7): a non-treated control (NC) group, a radiotherapy (RT) group, and a group that received IEt supplementation along with radiotherapy (IEt). During the first 7 days, the IEt group received 600 mg/kg/day [17] of IEt (Shanghai Yuanye Bio-Technology Co., Ltd., China) via oral administration, dissolved in a vehicle solution of 20% DMSO and 80% water. Simultaneously, the other two groups received the same volume of the vehicle solution through gavage. On day eight, the RT and IEt groups were subjected to a single dose of high-intensity whole-abdomen radiation using 6 MV X-rays produced by medical linear accelerators (Elekta, Sweden). Radiation was delivered at a total dosage of 12 Gy at a rate of 300 MU/min, with a source-to-skin distance of 100 cm, and the radiation field covered the area from the xiphoid to the pubic symphysis [18]. The NC group did not receive radiation. Following the treatment, the mice were fed a common diet with water and chow freely available for 3 days before euthanasia. Animal experiments were approved by the Animal Ethics Association of Yantai Yuhuangding Hospital (Approval number 2023-372).
Sample Collection
Upon completion of the experimental protocol, mice were euthanized via decapitation following anesthesia induced by intraperitoneal administration of pentobarbital sodium (50 mg/kg) [19]. Fecal samples were collected from mice prior to their euthanasia. Each mouse was housed in sterile cages, where the fecal samples were collected in sterile tubes, quickly frozen using liquid nitrogen, and stored at -80°C. These procedures were completed within a strict timeframe of 2 h. Blood samples were collected from the orbital sinus of the mice, combined with EDTA to prevent clotting, and centrifuged at 4°C, 3000 rpm for 10 min to separate the plasma. After dissection, the length of the colon was measured.
Enzyme-Linked Immunosorbent Assay (ELISA)
The levels of IL-17A, TNF-α, IFN-γ, IL-1β, IL-6, and IgA in the plasma of mice were measured using ELISA kits obtained from mlbio (China), in accordance with the protocols provided by the manufacturer.
Histopathological Analysis of the Colon in Mice
Following euthanasia, the colons were preserved in a 4% paraformaldehyde solution at 25°C. Afterward, 5 μm thick sections were prepared, embedded in paraffin, stained with hematoxylin and eosin (H&E) according to established protocols [20], and observed under an optical microscope (20×objective; Olympus Corp., Japan).
Metabolome Analysis
A total of 100 mg of feces was individually ground in liquid nitrogen and then suspended in prechilled 80%methanol with vigorous vortexing. The samples were kept on ice for 5 min, followed by centrifugation at 15,000 g for 20 min at 4°C. A portion of the supernatant was diluted to 53% methanol using LC-MS grade water. The samples were then transferred to new Eppendorf tubes and centrifuged again under the same conditions. The supernatant was subsequently analyzed using LC-MS/MS [21]. UHPLC-MS/MS analyses were conducted with a Vanquish UHPLC system (Thermo Fisher Scientific, Germany) paired with either an Orbitrap Q Exactive HF (Thermo Fisher Scietific) at Novogene Co., Ltd., (China). The samples were injected onto a Hypersil Gold column (100 × 2.1 mm, 1.9 μm) employing a 12-min linear gradient at a flow rate of 0.2 ml/min. Metabolite identification was conducted against a high-quality secondary spectrum database based on adduct ions with a mass deviation of 10 ppm. Background ions were eliminated using blank sample data, and quantitative results were normalized to achieve relative peak areas. Ultimately, the results for metabolite identification and relative quantification were obtained. Data processing was performed on a Linux operating system (CentOS version 6.6), using R and Python. These metabolites were annotated using the KEGG database (https://www.genome.jp/kegg/pathway.html). We applied univariate analysis (t-test) to calculate the statistical significance (p-value).The metabolites with VIP>1, p-value<0.05, and fold change≥2 or FC≤0.5 were considered to be differential metabolites. Volcano plots were used to filter metabolites of interest based on log2 (FoldChange) and -log10 (p-value) of metabolites by ggplot2 in R language. The raw data of untargeted metabolomics were deposited in the National Microbiology Data Center (NMDC) with the accession number NMDC10019551.
Metagenomic Analysis
DNA was isolated from mouse fecal samples using TIANamp Stool DNA Kit (TIANGEN Biotech Co., Ltd., China), adhering to the manufacturer's instructions. The genomic DNA was then randomly fragmented, and libraries were created for sequencing purposes. These fragments underwent end repair, A-tails addition, and the incorporation of adapters compatible with Illumina technology. PCR amplification was performed on the fragments that had been ligated with adapters, followed by size selection and purification. The quality and quantity of the constructed library were assessed utilizing Qubit and real-time PCR, while the analysis of fragment size distribution was performed using a bioanalyzer. After quantification, the libraries were pooled and prepared for sequencing on Illumina platforms (Novogene Co., Ltd.) in line with the required library concentration and data volume.
The Raw Data acquired from Illumina sequencing platforms underwent preprocessing using Readfq (https://github.com/cjfields/readfq) to produce Clean Data for subsequent analysis. The steps involved in preprocessing comprised: a) elimination of reads exhibiting a high proportion of low-quality bases (quality threshold ≤38, with a default length of 40 bp); b) removal of reads containing an excessive number of N bases; and c) eradication of reads demonstrating considerable adapter overlap. To mitigate potential host contamination, Clean Data were aligned with a reference genome database specific to the host to filter out reads likely originating from the host. This mapping was executed utilizing Bowtie2 software (http://bowtie-bio.sourceforge.net/bowtie2/index.shtml) with standard parameters: --end-to-end, --sensitive, -I 200, and -X 400 [22, 23].
The assembly process of the Clean Data utilizing the MEGAHIT software with the following parameters: --presets meta-large (--end-to-end, --sensitive, -I 200, -X 400) [22, 24]. The resulting Scaffolds were segmented at N junctions to create Scaftigs that did not contain N bases.
The raw data of metagenomic sequencing were stored in the NCBI Sequence Read Archive with the accession number PRJNA1165328.
Statistical Analysis
Comparisons between groups were carried out using the t-test implemented in the SPSS program (version 24.0; SPSS Inc., USA). The findings are presented as mean ± standard deviation (SD), with a p-value below 0.05 considered statistically significant (*p < 0.05, **p < 0.01, ***p < 0.001).
Results
Effects of IEt on Colon Structure
H&E staining results demonstrated that the colon structure in the NC group was intact, exhibiting clear crypts. In contrast, the colon structure in the RT group displayed damage, characterized by concave morphology, reduced crypt numbers, and structural destruction of the crypts (Fig. 1B). The crypts were compromised after radiation, whereas IEt treatment effectively restored the crypt structure in the mice. Comparative analysis of colon length (Fig. 1C) revealed a significant decrease in the RT group compared to the NC group (p = 0.028). Additionally, no statistically significant differences in colonic length were observed when comparing the IEt group with either the RT or NC groups.
Anti-Inflammatory Effects of IEt
To further investigate radiation enteritis, we assessed the levels of inflammatory markers, including IL-17A, IL-6, IFN-γ, IL-1β, and IgA, in the plasma of mice (Fig. 1D-1H). Notably, the t-test results showed that the level of IL-17A in the RT group increased by 10.8% compared to the NC group (p = 0.087), while the level of IL-17A in the IEt group significantly decreased by 14.98% compared to the RT group (p = 0.0005). The RT group exhibited reduced IFN-γ levels compared to the NC group, a difference that was alleviated by IEt intervention. Additionally, no significant changes were detected in the levels of IL-1β, IL-6, and IgA.
IEt Improved the Microbiota Disorder
The Venn diagram illustrated the distribution of genes among the groups and highlighted the common genes shared (Fig. 2A). Subsequently, differences in the composition of intestinal microbiota across the three groups were assessed utilizing the non-metric dimensional scaling (NMDS) approach (Fig. 2B) and the analysis of similarities (ANOSIM) method based on Bray-Curtis distance at the family level (Fig. 2C). The results indicated significant inter-group variation in gut microbiota compared to intra-group differences (p < 0.05).
Evaluation of the intestinal microbiota at the phylum level revealed that Bacteroidota and Bacillota (Firmicutes) were the dominant phyla across all three study groups (Fig. 2D). After radiotherapy, a significant decline was observed in the prevalence of the Thermodesulfobacteriota and Deferribacterota phyla, while the Pseudomonadota phylum experienced a substantial increase in the RT group compared to the NC group. Furthermore, the IEt group exhibited significantly lower relative abundances of the Actinomycetota phylum and Bacteroidota when contrasted with the RT group.
Analysis of intestinal microbiota at the family level indicated that Muribaculaceae was the dominant microbial family across all three groups (Fig. 2E). After radiotherapy, the RT group exhibited a substantial increase in the prevalence of Bacteroidaceae, Clostridiaceae, and Prevotellaceae families, accompanied by a notable decrease in the Lachnospiraceae family (p = 0.0033) compared to the NC group. In the IEt group, in contrast to the RT group, there was a significant decline in the prevalence of Muribaculaceae and Erysipelotrichaceae families, while Lachnospiraceae (p = 0.0037) and Oscillospiraceae families experienced a significant increase.
The Core Strain Affected by IEt Was Lachnospiraceae
Analysis of MetaStat at the genus level (Fig. 3A) revealed a significant increase (p <0.05) in the relative abundance of Parasutterella following radiotherapy in contrast to the decrease (p < 0.01) observed after IEt treatment. Additionally, the RT group exhibited a notable reduction (p < 0.05) in the relative abundances of Gemmiger (p = 0.0476), Roseburia (p = 0.0344), Colidextribacter (p = 0.0241) and Oribacterium (p = 0.0265), whereas the IEt group experienced a marked increase (p < 0.05) in their abundance. Among the top 35 genera identified in the MetaStat analysis, a total of seven genera belonging to the family Lachnospiraceae were identified, namely Eisenbergiella, Roseburia, Dorea, Sporofaciens, Lacrimispora, Oribacterium, and Lachnotalea. Among these genera, Roseburia (p = 0.0344), Dorea (p = 0.0075), Sporofaciens (p = 0.0265), Lacrimispora (p = 0.0241) and Oribacterium (p = 0.0265) significantly decreased after radiotherapy, and Eisenbergiella (p = 0.0466) and Roseburia (p = 0.0363) significantly increased following IEt treatment. The abundance of the genus Roseburia decreased by 55.87% in the RT group compared to the NC group, and increased by 155.14% in the IEt group compared to the RT group.
LEfSe analysis (Fig. 3B) identified 49 distinct taxa across the three study groups (LDA score > 4). The NC group exhibited an enrichment of Mucispirillum schaedleri, Bacteroidales bacterium, Eubacterium plexicaudatum, and other bacterial species. After radiotherapy, there was an enrichment of Dubosiella newyorkensis, Faecalibaculum rodentium, Duncaniella freteri, and additional bacteria. After IEt supplementation, Bacteroides acidifaciens, Lachnospiraceae bacterium, and other species were enriched.
Analysis of microbiota at the species level (Fig. S1) showed that, compared to the NC group, the abundances of several species, including Lachnospiraceae bacterium, Lachnospiraceae bacterium AM48-27BH, Sporofaciens musculi, Hungatella hathewayi, and Dorea sp. 5-2, were significantly altered post-radiotherapy, returning to baseline levels following IEt. Among the 35 most abundant species, six species belonging to the family Lachnospiraceae were noted: Lachnospiraceae bacterium MD335, Lachnospiraceae bacterium, Sporofaciens musculi, Lachnospiraceae bacterium AM48-27BH, Dorea sp. 5-2, and Otoolea muris, all of which exhibited significant changes in abundance (p < 0.05) after radiotherapy or IEt treatment. In addition, the t-test was conducted with a 95% confidence interval among the three groups (Fig. 4A-4F). The results showed that the abundance of species belonging to the Lachnospiraceae family, including Lachnospiraceae bacterium (p = 0.0140), Lachnospiraceae bacterium MD335 (p = 0.0200), Sporofaciens musculi (p = 0.0037), Lachnospiraceae bacterium AM48-27BH (p = 0.0210), Otoolea muris (p = 0.0170), and Dorea sp. 5-2 (p = 0.0009) were significantly decreased (p < 0.05) after radiotherapy. Among these, Lachnospiraceae bacterium (p = 0.0140), Lachnospiraceae bacterium MD335 (p = 0.0047), Sporofaciens musculi (p = 0.0190), and Dorea sp. 5-2 (p = 0.0920) significantly increased in the IEt group compared to the RT group.
Correlations between Lachnospiraceae and IL-17A
Spearman correlation analysis (Fig. 4G-4H) revealed that, at the genus level, IL-17A exhibited significant negative correlations with genera from the Lachnospiraceae family, including Eisenbergiella (p = 0.0025), Roseburia (p = 0.0029), Dorea (p = 0.0190), Sporofaciens (p = 0.0150), Lacrimispora (p = 0.0180), Oribacterium (p = 0.0110), and Lachnotalea (p = 0.0120). At the species level, IL-17A was significantly negatively correlated with species from the Lachnospiraceae family, such as Lachnospiraceae bacterium MD335 (p = 0.0007), Lachnospiriceae bacterium (p = 0.0036), Sporofaciens muculi (p = 0.0150), and Dorea sp. 5-2 (p = 0.0440).
Effects of IEt on Microbial Metabolic Functions
We analyzed the top 20 metabolic pathways of microbial metabolic functions (Fig. 5). Notably, metabolism exhibited the highest abundance at level 1 (Fig. 5A). Moreover, at level 2 (Fig. 5B), pathways such as carbohydrate metabolism, amino acid metabolism, and energy metabolism were particularly prominent. After radiotherapy, the abundance of metabolic pathways was significantly increased, including the biosynthesis of secondary metabolites, glycan biosynthesis and other amino acid metabolism, which were reversed by IEt. In contrast, the pathway of cell motility exhibited a significant decrease in abundance after radiotherapy, which was subsequently increased by IEt treatment. At level 3 (Fig. 5C), the abundance of amino sugar and nucleotide sugar metabolism, purine metabolism and pyruvate metabolism significantly increased in the RT group relative to the NC group, but these changes were reversed by IEt treatment.
LEfSe analysis at level 2 (Fig. 5D) revealed that the NC group exhibited enrichment in pathways such as metabolism of cofactors and vitamins, energy metabolism, and xenobiotics biodegradation. Additionally, the RT group showed enrichment in pathways related to cardiovascular disease, neurodegenerative disease, and drug resistance antineoplastic, while the IEt group displayed enrichment in pathways related to drug resistance antimicrobial, immune system, endocrine and metabolic disease. An in-depth examination at level 3 (Fig. 5E) using LEfSe revealed the enrichment of various metabolic pathways in all three groups. Notably, the NC group demonstrated enrichment in metabolic processes associated with porphyrin metabolism, arginine biosynthesis, citrate cycle (TCA cycle), and other entries. The RT group exhibited enrichment in pathways related to ribosome, glycolysis/gluconeogenesis, and other entries. Meanwhile, the IEt group displayed enrichment in propanoate metabolism, base excision repair, mismatch repair, and other entries.
Effects of IEt on Carbohydrate-Active Enzymes
The collective relative abundances of carbohydrate-active enzymes (CAZymes) were determined for each group based on the Carbohydrate-Active enzymes (CAZy) database (Fig. S2A). After radiotherapy, there was a significant increase in the total abundance of CAZymes, which was notably reversed by IEt. Specifically, radiotherapy induced a substantial elevation in the abundance of glycoside hydrolases (GH), glycosyl transferases (GT), carbohydrate-binding modules (CBM), polysaccharide lyases (PL), and carbohydrate esterases (CE). In contrast, IEt resulted in a reduction in the abundance of GH, GT, CBM, and CE (Fig. S2B). The analysis of the relative abundance of CAZymes at level 2 (Fig. S2C) indicated that radiotherapy significantly increased the abundance of CAZymes such as GH10, GT106, and CBM6, all of which were reduced by IEt. Conversely, radiotherapy significantly decreased the abundance of CAZymes such as GH18, CE12, and GT32, which were significantly increased by IEt.
Effects of IEt on Metabolites
A total of 3,013 metabolites were detected, with lipids and lipid-like molecules accounted for the highest proportion (30.87%), followed by organic acids and derivatives (22.73%), and organoheterocyclic compounds, which accounted for 16.76% (Fig. 6A). The statistical diagram of KEGG annotation showed the number of metabolites annotated to each secondary category under the primary pathway classification (Fig. 6B). The results showed that at level 1, the item of metabolism was annotated with the most metabolites, totaling 389. Among these, 137 metabolites were annotated in global and overview maps at level 2. This was followed by amino acid metabolism, which was annotated with 90 metabolites. Analysis of the volcano plot (Fig. 6C and 6D) showed that 153 metabolites were significantly increased and 216 metabolites were significantly decreased in the RT group compared to the NC group. Compared to the RT group, 76 metabolites were significantly increased, and 43 metabolites were significantly decreased in abundance in the IEt group. For example, the t-test results showed that the level of indole-3-carbinol decreased by 36.30% in the RT group compared to the NC group (p = 0.0550), while it increased by 57.77% in the IEt group compared to the RT group (p = 0.0064). Additionally, indole and its derivatives were significantly altered (Fig. 6E). Compared to the NC group, the abundance of indole and its derivatives including flustramine F, lycoperodine 1,3-indoleglyoxylic acid, 7-chloro-L-tryptophan, 1,3-dimethylindole, pindolol, and indole, significantly decreased, while the abundance of hyrtioerectine C, 3-hydroxyindolin-2-one-sulfate, (-)-horsfiline, and carbazolepropanoic acid increased significantly in the RT group. Compared to the RT group, the abundance of 3-methyldioxyindole, indole-3-carbinol, clausenine, and murrastifoline B was significantly upregulated and murrafoline was significantly downregulated in the IEt group. The changes in organic acids and derivatives were shown in the Sankey diagram (Fig. 6F).
In addition, we analyzed the relative abundance of differential metabolites and their correlation with significantly changed genera belonging to the Lachnospiraceae family (Fig. 7). Spearman correlation analysis showed that the metabolites with anti-inflammatory and anti-cancer effects, including indole-3-carbinol, pteridine, and pyropheophorbide-a, which were significantly upregulated by IEt, were significantly positively correlated with genera of Lachnospiraceae. Specifically, indole-3-carbinol was significantly positively correlated with genera Roseburia, Dorea, Lacrimispora, and Lachnotalea; pteridine exhibited a significant positive correlation with the genera Roseburia, Dorea, Sporofaciens, Lacrimispora, Oribacterium, and Lachnotalea; and a positive correlation was observed between pyropheophorbide-a and the genera Eisenbergiella, Lacrimispora, Oribacterium, and Lachnotalea.
Discussion
In this study, a mouse model of radiation-induced enteritis was used to investigate the effect of IEt on radiation enteritis. Analysis of intestinal microbiome composition exhibited significant differences among the three groups, suggesting that radiotherapy and IEt intervention significantly altered the composition and structure of the intestinal microbiome. At the family level, radiotherapy significantly downregulated the abundance of family Lachnospiraceae, which was significantly upregulated by IEt. Lachnospiraceae is a family of anaerobic bacteria in the class Clostridia with potential to advance the intestinal therapeutics. Members of this taxonomic group can ferment dietary fiber to promote healthy gut and immune function [25]. Previous studies have linked Lachnospiraceae to inflammatory diseases in the host. Decreased Lachnospiraceae abundance have been documented in patients with ulcerative colitis (UC) compared to healthy human controls [26]. Another study reported significantly lower levels of Lachnospiraceae taxa, including Roseburia and Coprococcus in ileal Crohn's disease (CD) patients compared to healthy human controls [27]. These findings are consistent with our results showing that mice with radiation-induced enteritis have a lower abundance of Lachnospiraceae.
IEt may play a radioprotective role by reducing radiation enteritis, which is mediated by family Lachnospiraceae. Lachnospiraceae have been shown to alleviate mucosal inflammation and reduce intestinal radiation injury in mice [28]. In addition, an enrichment of Lachnospiraceae in "elite survival mice" (mice that recovered and survived for a long time after a high dose of whole-body radiation), and subsequent causal experiments showed that the survival rate and clinical score of mice given Lachnospiraceae were greatly improved [11]. Additionally, the radioprotective metabolite mediators were identified as SCFAs and tryptophan metabolites. Treatment of irradiated mice with SCFAs and tryptophan metabolites significantly improved survival and reduced clinical scores [11]. To validate the effects of SCFAs in microbiome-mediated radioprotection, mice were treated with the SCFA producer of Lachnospiraceae strains, and the results revealed that high-producing SCFAs strains ensured complete protection against radiation for all mice, whereas low-producing SCFAs strains provided only 50%protection [11]. Thus, IEt can increase the abundance of Lachnospiraceae to alleviate radiation enteritis and play a radioprotective role in irradiated mice.
IEt reduced the abundance of genera belonging to family Lachnospiraceae including genus Roseburia. Roseburia has been identified for its role in regulating immune responses and decrease inflammation. Roseburia is a beneficial gut bacterium that plays a critical role in protecting against the onset of inflammatory bowel disease by promoting a healthy gut environment and potentially alleviating associated inflammation [29]. Roseburia could reduce inflammation through the suppression of the TLR-NF-κB pathway in HT-29 and Caco-2 cells [30]. Additionally, R. intestinalis supernatant inhibited colitis, altered inflammatory macrophage activity and pro-inflammatory cytokines, and inhibited Th17 cell differentiation in DSS- and TNBS (2,4,6-trinitrobenzenesulfonic acid solution)-induced IBD mouse models. SCFAs, including butyrate, were subsequently detected in R. intestinalis supernatants, which can exert an anti-inflammatory effect [31]. Administration of R. intestinalis significantly decreased IL-17 expression and increased the Treg ratio in mice with TNBS-induced colitis [32]. Moreover, the therapeutic application of R. intestinalis significantly improved DSS-induced colitis in mice regarding clinical symptoms, histological inflammation, and immune response by regulating Treg/Th17 cell balance and intestinal barrier integrity [33]. Th17 cells are critical members in mediating immune responses of adaptive immunity. In humans and mice, gut is a main site where Th17 cells reside. Radiation can induce the generation and accumulation of Th17 cells in the gut, with IL-17A primarily produced by these cells [34]. In clinical studies, the abundance of Roseburia has been found to be reduced in pediatric patients with UC [35] and in those with Crohn's disease [36]. Furthermore, researches indicated that Roseburia can enhance the sensitivity of colorectal cancer patients to radiotherapy by generating butyrate [37]. Additionally, Roseburia can suppress intestinal inflammation by producing butyrate in patients with colorectal cancer [38]. All these studies demonstrated the beneficial effects of genus Roseburia on intestinal inflammation-related diseases. In our study, we found that radiotherapy significantly reduced the abundance of genus Roseburia and upregulated the level of pro-inflammatory factor IL-17A, whereas IEt significantly increased Roseburia abundance and downregulated IL-17A levels, with a significant negative correlation observed between Roseburia and IL-17A. Thus, combined with the findings regarding Roseburia’s role in reducing inflammation, IEt may alleviate radiation enteritis through the genus Roseburia-IL-17A axis.
Subsequent analysis of the metabolic function of gut microbiota revealed that many metabolites, including indole-3-carbinol, pteridine, and pyropheophorbide-a, were significantly upregulated in irradiated mice treated with IEt. Notably, indole-3-carbinol is a bioactive substance with anti-inflammatory properties. An in vitro study showed that indole-3-carbinol ameliorated necroptosis and inflammation in NCM460 cells (a normal human colorectal cell line) [39]. In addition, multiple in vivo studies assessed the anti-inflammatory activity of indole-3-carbinol. A study in C57BL/6 mice treated with indole-3-carbinol for TNBS-induced colitis showed that indole-3-carbinol reduced the severity of the disease in female mice, as indicated by reduced weight loss and clinical symptom severity [40]. Likewise, indole-3-carbinol attenuated necroptosis and inflammation in intestinal epithelial cells (IECs) by activating aryl hydrocarbon receptor (AHR), which played a protective role in a mouse model of DSS-induced ulcerative colitis [39]. Furthermore, indole-3-carbinol has been reported to alleviate symptoms through stimulating IL-22 expression in a murine colitis model, which resulted in the generation of anti-inflammatory butyrate by the intestinal microbiome [41]. Moreover, another experiment revealed that indole-3-carbinol reduced the mRNA expression levels of pro-inflammatory cytokines, including IL-17A, IL-6, IL-1β, TNF-α and IFN-γ, in colon tissues of mice [42]. In our study, indole-3-carbinol levels significantly increased after intragastric administration of IEt, and a significant positive correlation was observed between indole-3-carbinol levels and genus Roseburia. Hence, combined with the anti-inflammatory effects of indole-3-carbinol, IEt may alleviate radiation enteritis through the genus Roseburia-indole-3-carbinol axis.
Interestingly, indole-3-carbinol has also been reported as a promising cancer preventive agent [43]. In vitro experiments showed that indole-3-carbinol, as an inhibitor of NF-κB and Akt activation, triggered G1 cell cycle arrest and apoptosis in prostate cancer cells [44]. In addition, several in vivo experiments have confirmed the anti-cancer effects of indole-3-carbinol. Indole-3-carbinol inhibited tumor growth in nasopharyngeal carcinoma in mice [45]. Indole-3-carbinol inhibited the development of cervical cancer in mice by upregulating PTEN expression [46]. In short, IEt can exert anti-inflammatory and radioprotective effects by regulating the abundance of Lachnospiraceae, and it also plays an anti-tumor role mediated by indole-3-carbinol during radiotherapy in mice.
Although this study identified IEt as a promising radioprotective metabolite, several limitations require consideration. Due to the lack of mechanistic studies involving bacterial depletion or pathway inhibition, causal claims regarding the Roseburia-indole-3-carbinol and Roseburia-IL-17A axes have not been verified. Furthermore, the scope of inflammatory profiling was restricted to select cytokines, omitting broader immune cell or spatial tissue analyses. To address these gaps, subsequent investigations will prioritize large animal validation in porcine radiation models – a critical step for clinical translation – alongside mechanistic studies in gnotobiotic mice and AHR-knockout systems to dissect microbial-metabolite-immune crosstalk. Targeted metabolomics will further resolve SCFA dynamics and other bioactive metabolites, ensuring comprehensive mechanistic elucidation.
In conclusion, IEt decreased the levels of the pro-inflammatory factor, such as IL-17A, and increased the abundance of anti-inflammatory Lachnospiraceae, such as Roseburia, as well as the levels of the anti-inflammatory metabolite indole-3-carbinol. Additionally, Roseburia showed a significant positive correlation with indole-3-carbinol and a significant negative correlation with IL-17A. Therefore, IEt may alleviate radiation enteritis through genus Roseburia-indole-3-carbinol and Roseburia-IL-17A axes (Fig. 8). This study identified a new tryptophan-derived microbial metabolite, IEt, as a protective agent against radiation enteritis, providing theoretical support for the prevention of radiation enteritis.
Supplemental Materials
Supplementary data for this paper are available on-line only at http://jmb.or.kr.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bray F Laversanne M Sung H Ferlay J Siegel RL Soerjomataram I 2024 Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries CA Cancer J. Clin.7422926310.3322/caac.2183438572751 · doi ↗ · pubmed ↗
- 2Hauer-Jensen M Denham JW Andreyev HJ 2014 Radiation enteropathy--pathogenesis, treatment and prevention Nat. Rev. Gastroenterol. Hepatol.1147047910.1038/nrgastro.2014.4624686268 PMC 4346191 · doi ↗ · pubmed ↗
- 3Gong L Zhang Y Liu C Zhang M Han S 2021 Application of radiosensitizers in cancer radiotherapy Int. J. Nanomedicine 161083110210.2147/IJN.S 29043833603370 PMC 7886779 · doi ↗ · pubmed ↗
- 4Jian Y Zhang D Liu M Wang Y Xu ZX 2021 The impact of gut microbiota on radiation-induced enteritis Front. Cell. Infect. Microbiol.1158639210.3389/fcimb.2021.58639234395308 PMC 8358303 · doi ↗ · pubmed ↗
- 5Yi Y Lu W Shen L Wu Y Zhang Z 2023 The gut microbiota as a booster for radiotherapy: novel insights into radio-protection and radiation injury Exp. Hematol. Oncol.124810.1186/s 40164-023-00410-537218007 PMC 10201781 · doi ↗ · pubmed ↗
- 6Andreyev J 2005 Gastrointestinal complications of pelvic radiotherapy: are they of any importance?Gut 541051105410.1136/gut.2004.06259616009675 PMC 1774900 · doi ↗ · pubmed ↗
- 7Moraitis I Guiu J Rubert J 2023 Gut microbiota controlling radiation-induced enteritis and intestinal regeneration Trends Endocrinol. Metab.3448950110.1016/j.tem.2023.05.00637336645 · doi ↗ · pubmed ↗
- 8Shadad AK Sullivan FJ Martin JD Egan LJ 2013 Gastrointestinal radiation injury: symptoms, risk factors and mechanisms World J. Gastroenterol.1918519810.3748/wjg.v 19.i 2.18523345941 PMC 3547560 · doi ↗ · pubmed ↗