Mannose receptor RpMR1 of Manila clam (Ruditapes philippinarum) defense against Vibrio anguillarum infection
Zhihui Yin, Hongtao Nie

TL;DR
This study identifies and characterizes the mannose receptor RpMR1 in Manila clams, showing its role in defending against bacterial infection.
Contribution
The first functional evidence of mannose receptor-mediated immunity in mollusks is provided.
Findings
RpMR1 significantly reduces mortality in Manila clams infected with Vibrio anguillarum.
RpMR1 exhibits antibacterial activity against Gram-negative bacteria but not Gram-positive bacteria.
RpMR1 interacts with TLR4 in the immune response, as shown by gene silencing and expression analysis.
Abstract
The mannose receptor (MR) is a member of the C-type lectin superfamily and a type I transmembrane protein that functions as a pattern recognition receptor (PRR) in immune responses. In this study, we identified 13 MR genes (RpMR1-13) in the genome of Ruditapes philippinarum and investigated their expression profiles following Vibrio anguillarum challenge. Notably, RpMR1, RpMR2, RpMR3, and RpMR4 exhibited peak expression at 72 h post-infection. We successfully purified the recombinant RpMR1 protein and demonstrated its antibacterial activity against three Gram-negative bacteria (V. splendidus, V. alginolyticus, and V. anguillarum), though it had no effect on Gram-positive bacteria. Furthermore, in vivo injection of RpMR1 significantly reduced mortality in R. philippinarum following V. anguillarum infection. To explore role of RpMR1 in immune signaling, we performed RNA interference…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
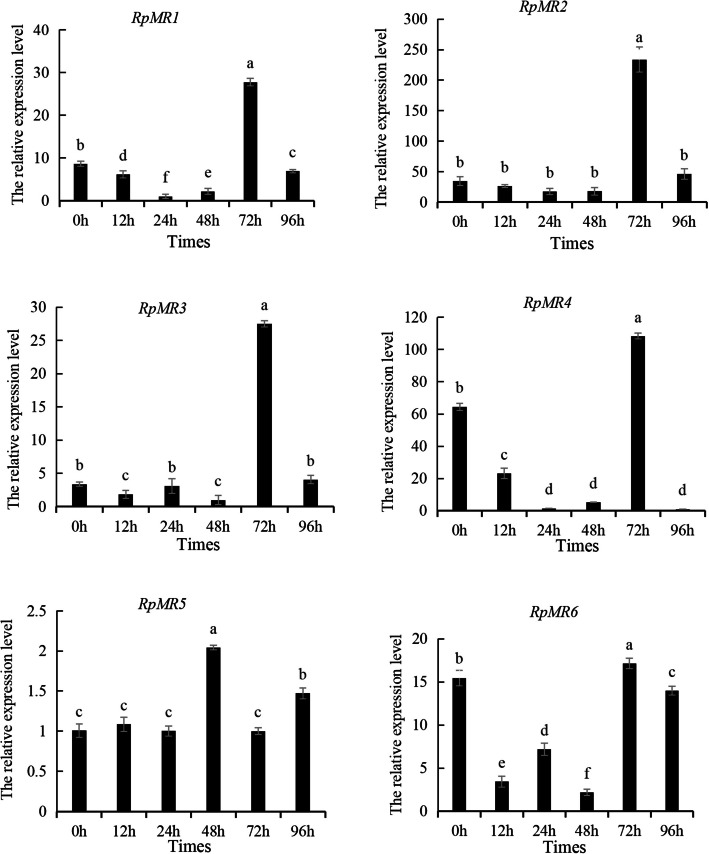 Figure 1
Figure 1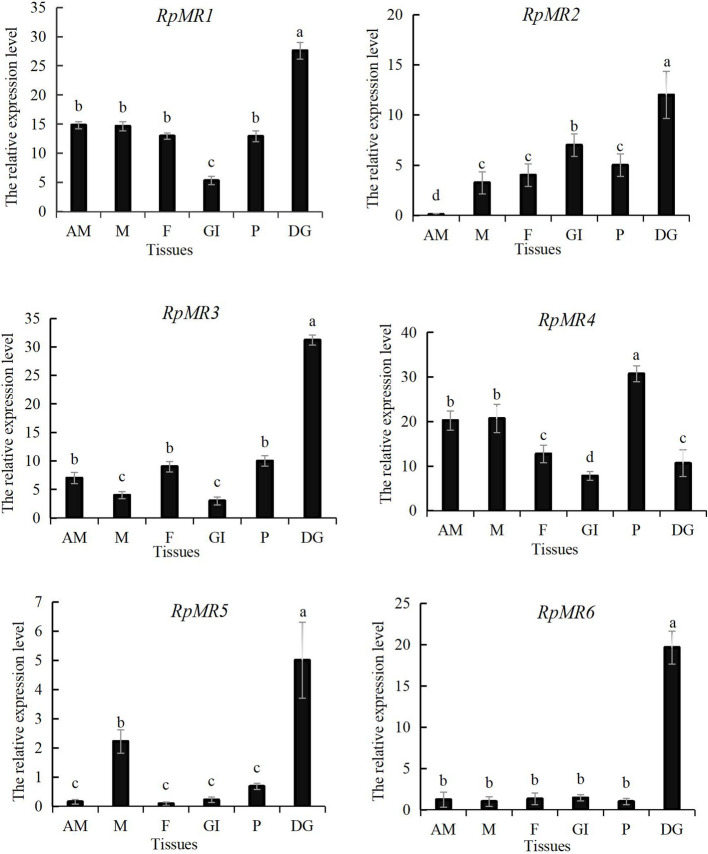 Figure 2
Figure 2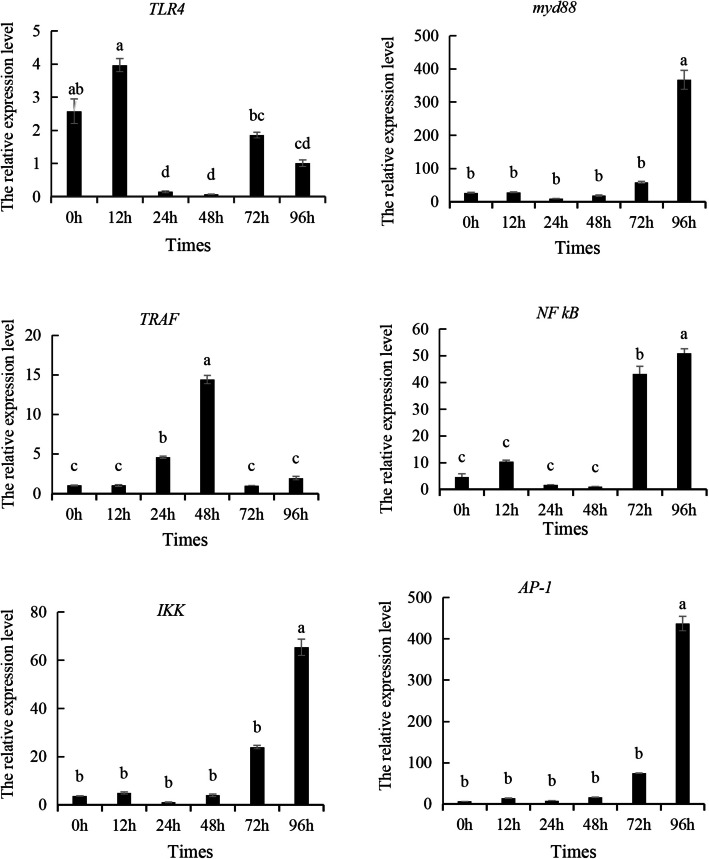 Figure 3
Figure 3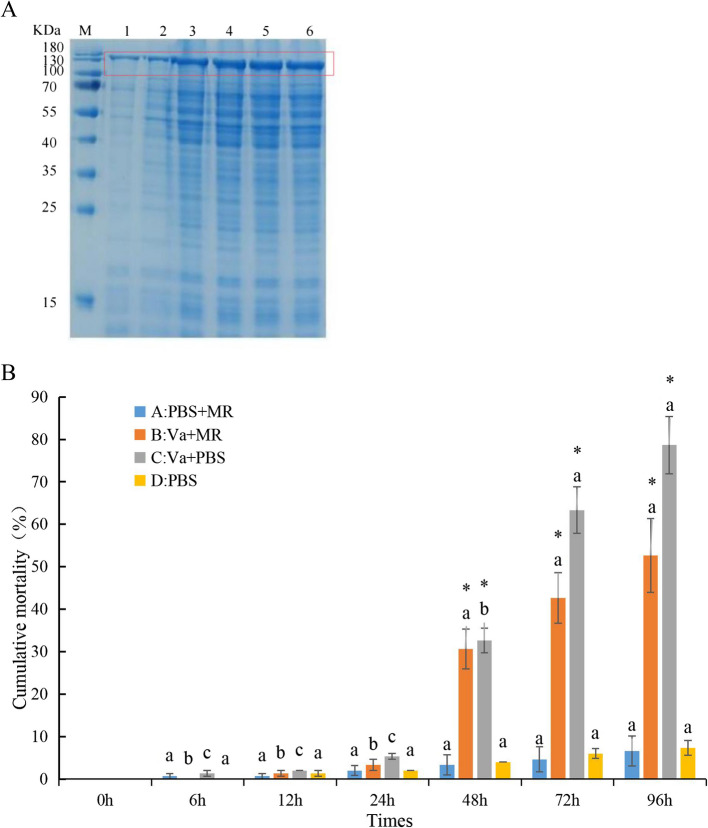 Figure 4
Figure 4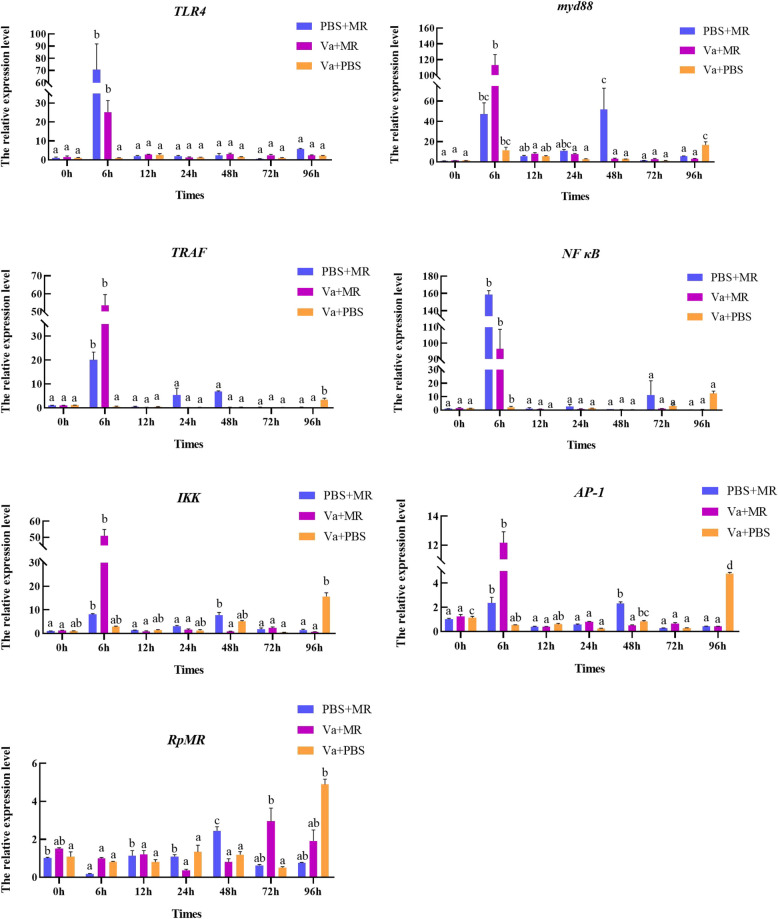 Figure 5
Figure 5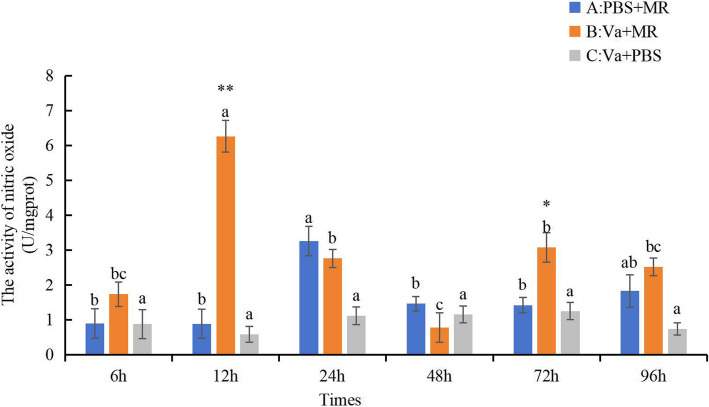 Figure 6
Figure 6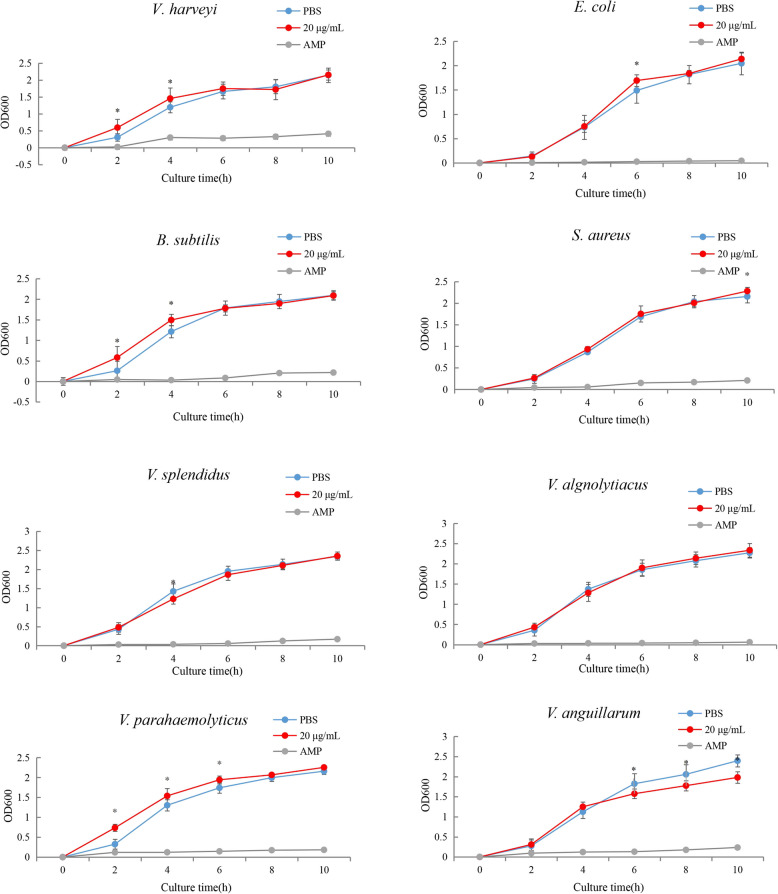 Figure 7
Figure 7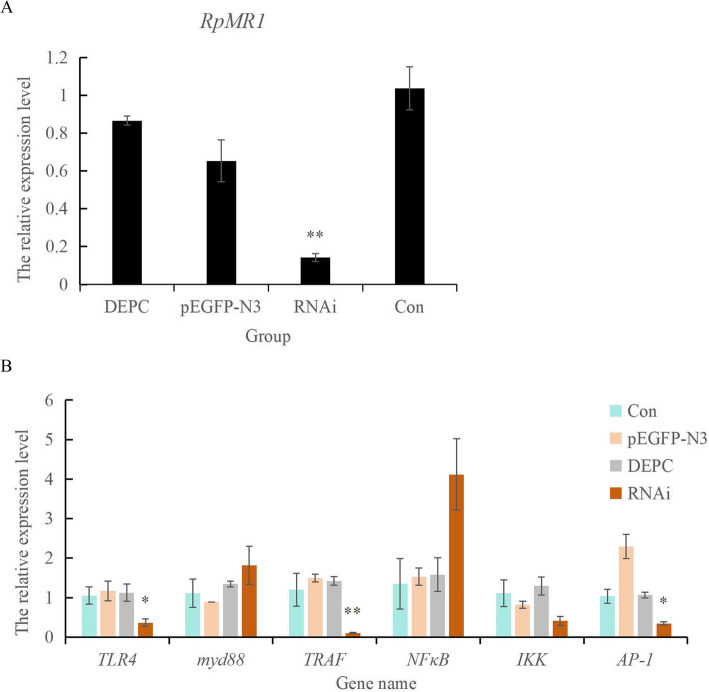 Figure 8
Figure 8- —Outstanding Young Scientific and Technological Talents Foundation of Dalian
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInvertebrate Immune Response Mechanisms · Aquaculture disease management and microbiota · Antimicrobial Peptides and Activities
Introduction
Mollusks, like many metazoans, rely on conserved immune strategies, including cellular and humoral responses, to combat pathogens (Tiscar et al. 2004). Collectively, cellular and humoral immunity enable mollusks to mount effective defense responses against environmental stressors, facilitating phagocytosis of foreign particles and detoxification of harmful substances (Soares-da-Silva et al. 2002). While mollusks possess a well-developed immune system, their defense mechanisms differ fundamentally from vertebrate immunity. Unlike vertebrates, which employ both adaptive and innate immune responses, mollusks rely exclusively on innate immunity—including enhanced phagocytic activity, pathogen recognition receptors, and effector molecules—to combat pathogenic invasion (Hoffmann et al. 1999; Little et al. 2005). Shellfish are widely recognized as susceptible to infection by various bacterial species during their growth, including V. tasmaniensis, V. crassostreae, V. parahaemolyticus, and V. anguillarum. Numerous studies have investigated the molecular mechanisms underlying shellfish resistance to these pathogens, focusing primarily on molecular genetic processes (Islam et al. 2022; Namwong et al. 2022). Accordingly, our previous transcriptomic studies on Ruditapes philippinarum subjected to Vibrio anguillarum stress have identified toll-like receptor (TLR) and MR as potential candidate genes for screening disease resistance in clams (Yin et al. 2022a, 2022b). Indeed, pattern recognition receptors (PRRs), including TLRs and mannose receptors (MRs), function as molecular sensors for the specific recognition of pathogens and play a pivotal role in the elimination of invading microorganisms (Li et al. 2020a; Ni et al. 2007).
The MR is a type of pattern recognition receptor (PRR) that functions as a "non-standard" PRR, primarily contributing to host defense by recognizing and binding to endogenous ligands or pathogen-associated molecules. Through this interaction, MR plays a key role in modulating the immune response and maintaining immune homeostasis under external stress (Gazi et al. 2009; Huo et al. 2024b; Yin et al. 2022b). The immunological relevance of MR-mediated responses has been highlighted in studies reporting antiviral and antibacterial activities against Vibrio species in aquatic animals such as Procambarus clarkii and Epinephelus coioides (Man et al. 2018; Zhang et al. 2021). As a member of the MR family within the C-type lectin receptor (CLR) superfamily, MR is classified as a type I transmembrane protein (Stahl et al. 1998). Structurally, it comprises an extracellular cysteine-rich (CR) domain, a fibronectin type II (FN II) domain, and eight tandem C-type lectin-like domains (CTLDs), along with a transmembrane segment and a short cytoplasmic tail (Gazi and Martinez-Pomares 2009). Functional studies have demonstrated a synergistic interaction between the FN II and CTLD2 domains, which enhances the uptake of glycosylated collagen. Additionally, CTLD4 has been identified as a critical domain involved in modulating T cell cytotoxicity (Martinez-Pomares 2012; Schuette et al. 2016). The MR protein also possesses ligand-binding capabilities that enable it to transduce signals from the extracellular environment into the cytoplasm. Its CTLDs can specifically recognize and bind carbohydrate ligands such as mannan, trehalose, and N-acetylglucosamine on various endogenous and exogenous molecules, subsequently initiating immune responses (Zhang et al. 2021). In Megalobrama amblycephala (bream), it has been shown that MR specifically binds to chitosan oligosaccharide (COS) and mediates COS6 uptake by macrophages via lectin-dependent endocytosis. This interaction modulates the expression of tumor necrosis factor receptor-associated factor (TRAF), interleukins (ILs), and nitric oxide synthase (NOS), underscoring the pivotal role of MR in ligand recognition, internalization, and immune regulation (Ouyang et al. 2021).
R. philippinarum commonly known as the Manila clam, belongs to the phylum Mollusca, class Bivalvia, order Eulamellibranchia, family Veneridae, and genus Ruditapes (Li et al. 2019). As an ecologically and economically important species, R. philippinarum exhibits several advantageous traits, including broad salinity and temperature tolerance, resistance to pollution, and rapid growth (Yin et al. 2022b). However, in recent years, the aquaculture industry of R. philippinarum has faced increasing threats due to bacterial infections, protozoan parasites, and environmental stressors (Le Roux et al. 2016; Nie et al. 2016; Waki et al. 2012). Among these, vibriosis—caused by the Gram-negative pathogen Vibrio anguillarum—is particularly detrimental, leading to hemorrhagic septicemia, high mortality in farmed clams, and significant economic losses (Ren et al. 2021).
Understanding the immune defense mechanisms of R. philippinarum against bacterial pathogens is essential for the development of genetically improved, disease-resistant strains. Transcriptomic analyses have provided key insights into the clam’s immune responses. For instance, studies investigating the immune response to V. parahaemolyticus infection have highlighted the critical role of the C1q gene in disease resistance(Yu et al. 2023b). Other transcriptomic investigations have demonstrated that exposure to various pathogen-associated molecular patterns (PAMPs) triggers the activation of immune effectors such as apextrin, along with downstream activation of the NF-κB signaling pathway (Jiang et al. 2020b, 2022). Our previous comparative transcriptomic study revealed the involvement of MR in the phagosome pathway, suggesting a significant role for MR in Vibrio tolerance in R. philippinarum (Yin et al. 2022a, b). Further supporting this, evidence from TLR studies underscores a cooperative interaction between MR and TLR4 in pathogen recognition, where crosstalk between PRRs enhances both innate and adaptive immune responses(Loures et al. 2015). Additionally, mannose-binding lectin (MBL)—another member of the C-type lectin receptor (CLR) family—has been shown to potentiate TLR4 signaling through direct interaction with its leucine-rich repeat (LRR) domain (Wang et al. 2011). Although MR and MBL differ in ligand specificity, their shared classification within the CLR family suggests conserved immunomodulatory functions in host defense.
In the present study, we identified and characterized the expression patterns of mannose receptor (MR) genes in R. philippinarum following V. anguillarum challenge, revealing a significant role for MR in the host immune response. In vitro antibacterial assays demonstrated that the recombinant MR protein (RpMR1) exerted inhibitory effects against three Gram-negative bacterial strains. Furthermore, in vivo overexpression of RpMR1 via injection significantly increased the survival rate of R. philippinarum under V. anguillarum infection, underscoring its protective function. Conversely, silencing of the RpMR1 gene through RNA interference led to a marked downregulation of TLR4 expression. This finding suggests that MR may activate the TLR signaling pathway during pathogen invasion, potentially interacting with TLR4 to facilitate phagocytosis and coordinate innate immune responses. Together, these data support a synergistic role for MR and TLR4 in mediating host defense against bacterial infection. Overall, this study provides novel insights into the immune mechanisms of R. philippinarum and highlights MR as a potential target for enhancing disease resistance in molluscan aquaculture.
Materials and methods
The clam genome data used in this study were obtained from our previous work (Yan et al., 2019), with the NCBI biological project number PRJNA479743(Yan et al. 2019). Candidate MR genes were screened using Hidden Markov Models (HMMs) (Jiang et al. 2021). The putative isoelectric point (PI) and molecular weight of the proteins were calculated using the Compute pI/Mw tool (http://web.expasy.org/compute_pi/, http://web.expasy.org/protparam/) (Jiang et al. 2020a). Genome sequences of MR genes from other organisms were downloaded, with relevant database identifiers provided in Table S1. Multiple sequence alignments of RpMR genes were performed using DNAMAN software (https://www.lynnon.com/dnaman.html). Phylogenetic analysis was conducted using MEGA 7.0 (http://www.megasoftware.net) employing the maximum likelihood (ML) method, followed by 1000 bootstrap replicates for tree construction (Bai et al. 2020).
RpMR gene structure, chromosome location and evolution analysis
The gene structure of RpMR was analyzed using the Gene Structure Display Server (GSDS) 2.0 (http://gsds.gao-lab.org/index.php), which allowed for the visualization of the gene’s coding sequence (exons), untranslated regions (UTRs), and introns, derived from the gene annotation file(Hu et al. 2015; Jiang et al. 2021; Yin et al. 2022a). Functional and evolutionary analysis of characterized sequences was performed using the MEME web server, which provides a comprehensive platform for sequence motif discovery and analysis (Bailey et al. 2009; Jiang et al. 2021; Yin et al. 2022a). Chromosomal location data for RpMR genes were retrieved from the R. philippinarum genome annotation file(Jiang et al. 2021; Wang et al. 2020; Yin et al. 2022a). The chromosomal positions and relative distances of RpMR genes were visualized using MG2C 1.1 (http://mg2c.iask.in/mg2c_v1.1/).
Infection experiment of V. anguillarum in R. philippinarum
Infection experiments were conducted at various time points following V. anguillarum exposure using wild clam populations collected from Jinshitan, Dalian City, Liaoning Province. The clams (shell length: 30.8 ± 0.3 mm, shell width: 12.4 ± 0.3 mm, shell height: 20.1 ± 0.4 mm, total weight: 7.4 ± 0.4 g) were acclimatized in tanks containing seawater (water temperature: 21 ± 0.3 °C, pH: 8.2 ± 0.1, salinity: 32 ± 0.2 ppt). The experimental system consisted of nine 30-L aquaria, arranged in triplicate groups (three tanks per treatment), each housing 70 R. philippinarum specimens. During the 7-day acclimatization phase, the clams were reared in clean seawater and fed daily with Chlorella vulgaris. For the challenge experiment, clams were immersed in V. anguillarum at a concentration of 1 × 10⁷ CFU/mL. Samples were collected at 0-, 12-, 24-, 48-, 72-, and 96-h post-challenge, with each time point sampled in triplicates. Hepatopancreas tissues were harvested, snap-frozen in liquid nitrogen, and stored at −80°C for subsequent RNA extraction.
Collection of samples from different tissues and different developmental stages
Wild clams (Ruditapes philippinarum) were collected from Jinshitan, Dalian, Liaoning Province. Tissue samples from adults included adductor muscle (AM), mantle (M), foot (F), gill (GI), pipe (P), and digestive gland (DG). Developmental stages analyzed were embryos and larvae, categorized as fertilized egg (FE), first polar body (PB1), second polar body (PB2), 2-cell (TC), 8-cell (EC), blastula (B), gastrula (G), trochophore larvae (T), D-larvae (D), Umbo larvae (U), pediveliger larvae (P), single siphon spat (S) and juveniles (J). The expression of MR gene (MR1-MR6) was assessed across tissues and developmental stages by qPCR and RNA-seq analysis, respectively. The expression characteristics of MR genes were performed based on transcriptome sequencing data at different developmental stages of R. philippinarum (Zhang et al. 2022).
Extraction of RNA and synthesis of cDNA from R. philippinarum
Three biological replicates of clam samples were analyzed per experimental group. Total RNA was extracted using the phenol–chloroform method (Yu et al. 2023b; Zhang et al. 2022). TransScript ® IIOne Step gDNA Removal and cDNA Synthesis SuperMix (Beijing, all gold) reverse transcription kit is used to synthesize cDNA. The reverse transcription reaction (20 μL total volume) contained the following components: 500 ng of total RNA, use 0.5 μL of anchored Oligo (dT) 20 Primer (0.5 μg/μL), 0.5 μL of random primer (N9) (0.1 μg/μL), 10 μL of 2 × TSII Rection Mix, 1 μL of TranScript IIRT/RI Enzyme Mix, 1 μL of gDNA remover, RNase-free water was added to adjust the final volume to 20 μL. The reaction mixture was incubated at 50 °C for 15 min and heat at 85 °C for 5 s, and the resulting cDNA was stored at −20° C.
qPCR analysis
Fluorescent quantitative primers were designed using Primer 5 software (Premier Biosoft International). Primer sequences are listed in Table S2 (Li et al. 2019, 2020b). The β-actin was used as the internal reference gene (Yin et al. 2022b). Real -time quantitative PCR uses TB Green Premix Extaqii (TAKARA, Tokyo, Japan) (Jiang et al. 2022). The total reaction volume was 20 μL, comprising 2 μL of cDNA (50 ng/μL), 1 μL of positive and negative primers, 10 μL of TB Green PCR Master Mix, and 6 μL of H₂O. The amplification conditions were as follows: initial denaturation at 95 °C for 3 min, followed by 40 cycles of denaturation at 94 °C for 30 s, annealing at 60 °C for 30 s, and extension at 72 °C for 30 s. Relative gene expression levels were analyzed using the 2^^−ΔΔCT^ method (Schmittgen et al. 2008). Statistical analysis of the data was performed using SPSS 20.0. Single-factor analysis of variance (ANOVA) followed by the Tukey's test was used to assess significant differences between the groups. A p-value of less than 0.05 (P < 0.05) was considered statistically significant.
RpMR1 prokaryotic expression technology
The prokaryotic expression of the RpMR1 gene was carried out at Shanghai Sangon Bioengineering Co., Ltd. The coding region of the gene was amplified by PCR to obtain a sufficient quantity of the product, which was then inserted into the PET-28A (+) vector using restriction enzymes. The recombinant plasmid was introduced into E. coli Rosetta (DE3) cells through transformation. The transformed cells were plated on LB agar containing 30 µg/mL kanamycin and incubated at 37 °C. A single colony was selected and cultured in liquid medium with kanamycin at 37 °C. Once the OD600 reached 0.6, protein expression was induced by adding 0.5 mM IPTG, and the cells were incubated for an additional 6 h. After harvesting, the cells were resuspended in 1 × PBS (pH 7.4) and subjected to ultrasonic disruption. The suspension was centrifuged to separate the supernatant and pellet, and the pellet was resuspended in 50 mM Tris, 300 mM NaCl, and pH 8.0. The protein was solubilized in a buffer containing 8 M urea, 50 mM Tris, 300 mM NaCl, and 0.1% Triton X-100 (pH 8.0). After ultrasonic treatment, the mixture was centrifuged to obtain the supernatant. Protein purification was achieved through dialysis against increasing concentrations of urea in Tris-NaCl buffer. Protein expression and purity were confirmed by SDS-PAGE using a 12% separation gel and 5% stacking gel, with molecular weight estimation by comparison to protein standards. The purified protein was concentrated using PEG20000 after dialysis, filtered through a 0.45 µm filter, and stored at −80°C for future use (1:1000, Sangon Biotech, China).
Injection of RpMR1 recombinant protein experiment and NOS detection
Wild clams from Jinshi Beach in Dalian were used for V. anguillarum injection experiments. The clams were randomly assigned to four groups: A (PBS + MR), B (Va + MR), C (Va + PBS), and D (PBS control group). The injection volumes of V. anguillarum, recombinant protein, and PBS were 50 μL each, with concentrations of 1 × 10⁷ CFU/mL for V. anguillarum, 200 μg/mL for the recombinant protein, and PBS as the control. Mortality rates were recorded at 0, 6, 12, 24, 48, 72, and 96 h post-injection. Hepatopancreas tissues were collected, immediately frozen in liquid nitrogen, and stored at −80°C. Nitric oxide (NO) activity was measured using the nitric oxide assay kit (Nanjing Jiancheng, China) following the manufacturer's instructions.
RpMR1 protein bacteriostatic experiment
The recombinant RpMR1 protein was tested for bacteriostatic activity against eight different bacterial strains, including B. subtilis, S. aureus, V. anguillarum, V. parahemolyticus, V. harveyi, V. splendidus, V. algnolytiacus and E. coli. After the bacteria were activated for a second time, a coating experiment was performed, followed by inoculation of each bacterial strain into 10 mL of LB broth. The recombinant protein was added to a final concentration of 20 μg/mL. The cultures were incubated for 10 h under constant shaking conditions (37°C, 200 rpm). During the experiment, bacterial growth was monitored by measuring the optical density at 600 nm (OD600) every 2 h (Jiang et al. 2022).
RNA silencing
For the dsRNA injection experiments, wild clams collected from Jinshi Beach in Dalian were used. The clams were divided into four groups: experimental group (RNAi), negative control group (pEGFP), positive control group (DEPC), and blank control group (Con), with three clams in each group. In the experimental group, 25 μg/100 μL of synthetic dsRNA RpMR1 interference strand was injected. The negative control group received 25 μg/100 μL of dsRNA-pEGFP-N3 interference strand (purchased from Dalian Jima Company, China) (Yu et al. 2023a). The positive control group was injected with 100 μL of DEPC, while the blank control group received no treatment. After 36 h post-injection, hepatopancreas tissues were collected, snap-frozen, and stored at −80°C for subsequent analysis.
Ethic statement
All animal procedures in this experiment were conducted in accordance with the ethical guidelines set by the Experimental Animal Center of Dalian Ocean University to minimize animal suffering. The study adhered to national laws and regulations governing the use of experimental animals.
Results
Identification of RpMR gene family
In this study, a total of 13 mannose receptor (MR) genes, designated RpMR1 to RpMR13, were identified in the R. philippinarum genome (NCBI BioProject: PRJNA479743). These genes were identified based on the presence of C-type lectin-like domains (CTLDs) using the HMMER search with the Pfam model PF00059.23. The detailed characteristics of the identified RpMR genes are summarized in Table 1. The predicted RpMR proteins exhibit a wide range of molecular weights, from 11.27 kDa to 320.37 kDa, and theoretical isoelectric points (pI) ranging between 4.42 and 6.82, suggesting functional and structural diversity among the MR family members in R. philippinarum. Table 1. The detail information of MR gene family members in R. philippinarum genomeGene nameGene IDChr numberChromosome lengthChromosomal locationGene standMolecular weight (kDa)PIRpMR1xfSc0000262.81041,560,981 33,350,163–33378259-311.114.85RpMR2xfSc0000020.91041,560,98110,863,413–10884496 + 131.828.35RpMR3xfSc0001192.5456,440,29418,273,111–18,310,054 + 70.595.01RpMR4xfSc0000495.61041,560,98117,403,407–17415810-83.334.85RpMR5xfSc0005707.10---62.144.54RpMR6Sc0000045.9937,028,85435,020,290–35027760 + 53.115.04RpMR7xfSc0000495.71041,560,98117,418,034–17480613-320.375.19RpMR8xfSc0000519.41550,027,94135,425,935–35,455,941 + 142.816.37RpMR9xfSc0000519.51550,027,94135,409,151–35424993 + 74.436.82RpMR10xfSc0000108.301233,673,78723,660,287–23663732 + 31.476.74RpMR11xfSc0001787.6158,439,1403,112,136–3,113,018 + 18.175.75RpMR12xfSc0002266.50-- + 51.715.59RpMR13xfSc0003897.10-- + 11.274.42
RpMR gene family structure, conserved motif analysis and chromosome location
The 13 identified RpMR genes exhibited considerable structural diversity, with exon numbers ranging from 2 to 60. Phylogenetic analysis and intron–exon structure comparisons are presented in Figure S1, illustrating evolutionary relationships among the RpMR family members. Conserved motif analysis revealed 10 distinct motifs (designated motifs 1–10) across the RpMR proteins (Fig. S2), while domain architecture analysis identified seven major structural domains: C-type lectin domain (CLECT), tetratricopeptide repeat (TR), coagulation factor 5/8 C-terminal domain (FA58C), fibrinogen C-terminal domain (FTP), kringle domain (KR), signal peptide, and transmembrane region (Fig. S3). All RpMR proteins contained at least one CLECT domain, with copy numbers ranging from 1 to 14 per protein, indicating potential functional redundancy or diversification. Multiple sequence alignment of RpMR1–RpMR13 highlighted several conserved amino acid residues, including cysteine residues, Ca^2^⁺-binding sites, acidic amino acids, and aromatic residues such as phenylalanine and tryptophan (Fig. S4), which are characteristic features of C-type lectin family proteins. Chromosomal localization analysis revealed that RpMR genes are distributed across several chromosomes, with 10 genes mapped to six annotated chromosomes. Notably, chromosome 10 harbors three RpMR genes, indicating a possible gene cluster (Fig. S5). The remaining three genes—RpMR5, RpMR12, and RpMR13—were located on unplaced scaffolds. Comprehensive annotations, including chromosome lengths and genomic coordinates, are provided in Table 1.
Phylogenetic analysis of the RpMR gene family
To investigate the evolutionary relationships of the RpMR gene family, a phylogenetic tree was constructed using 142 MR amino acid sequences from nine representative species, selected based on the presence of the C-type lectin-like domain (HMM model PF00059.23) (Fig. S6). The phylogenetic analysis revealed that MR genes cluster into two major branches. Notably, MR sequences from molluscan shellfish, including R. philippinarum, Crassostrea virginica, C. gigas, and Biomphalaria glabrata, grouped predominantly within a single large clade. These results suggest that the MR genes in mollusks have undergone evolutionary conservation, reflecting shared ancestry and potentially conserved functional roles across molluscan species.
Expression pattern of RpMR gene at different time points after V. anguillarum stress
The expression pattern of RpMR gene at different time points after V. anguillarum challenge was shown in Fig. 1. The results showed that RpMR expression was greatly increased following V. anguillarum challenge. Notably, the expression levels of all RpMR1, RpMR2, RpMR3, RpMR4, and RpMR6 peaked at 72 h post-infection, reaching 3.2-, 6.8-, 8.3-, 1.68-, and 1.1-fold increases, respectively, relative to the baseline expression at 0 h. These findings suggest a time-dependent transcriptional regulation of RpMR genes, particularly during the late phase of the immune response to V. anguillarum.Fig. 1. Expression of RpMR genes at different time points after V. anguillarum stress. Note: Different letters indicate that there are significant differences in gene expression at different time points (P < 0.05)
Expression pattern of RpMR gene in different tissues
The expression of RpMR gene in different tissues of clam was examined by qPCR (Fig. 2). Results showed that RpMR was expressed in adductor muscle, mantle, foot, gill, siphon, and digestive gland, which mainly expressed in the hepatopancreas. The expression of RpMR1 was significantly higher in hepatopancreas tissue (twofold), compared with adductor muscle, mantle, foot and gill tissue (P < 0.05). The expression level of RpMR2 gene was the highest in the hepatopancreas, followed by the gill tissue, and the expression level in the adductor muscle was the lowest. The expression of RpMR2 was significantly higher in hepatopancreas tissue (12-fold), compared with adductor muscle tissue (P < 0.05). The expression level of RpMR3 gene was the highest in the hepatopancreas and the lowest in the gills. The tissue-specific expression analysis revealed that RpMR3 was significantly upregulated in the hepatopancreas, with expression levels exceeding a threefold increase compared to other tissues (P < 0.05). Similarly, RpMR4 and RpMR5 also exhibited their highest expression levels in the hepatopancreatic tissue, suggesting that this organ may serve as a key site for mannose receptor-mediated immune responses in R. philippinarum.Fig. 2. The expression of the RpMR gene in different tissues of Manila clam. Note: Different letters indicate that genes are significantly different in different tissues (P < 0.05). Adductor muscle (AM), mantle (M), foot (F), gill (GI), pipe (P), digestive gland (DG)
Detection of differentially expressed of TLR pathway genes under stress of V. anguillarum
The expression levels of genes related to TLR pathway had different expression characteristics after V. anguillarum stress. As shown in Fig. 3, the expression of the TLR gene in response to V. anguillarum stress exhibited a dynamic pattern. Expression levels significantly increased at 12 h post-infection, decreased at 24 and 48 h, and rose again at 72 h. Furthermore, most genes within the TLR signaling pathway were activated during the infection. Notably, the downstream signaling components to the TLRs-including myeloid differentiation primary response gene 88 (MyD88), nuclear factor kappa-B (NF-κB), IκB kinase (IKK), activator protein-1 (AP-1), and tumor necrosis factor receptor-associated factor 6 (TRAF6)—were significantly upregulated, highlighting the involvement of the TLR pathway in orchestrating immune responses to V. anguillarum.Fig. 3. Expression of Toll-like receptor pathway related genes in V. anguillarum under stress. Note: Different letters indicate that there are significant differences in gene expression at different time points (P < 0.05)
Expression patterns of RpMR genes in different developmental stages
The RpMR gene expression at different developmental stages were plotted into a heat map (Fig. S7). The results showed that all RpMR genes expressed with the growth and development of the clam, and the expression of most RpMR genes increased significantly in the D larva. The expression of the RpMR1 gene in the pediveliger larvae stage was decreased compared to the Umbo-larvae stage, but higher than other RpMR genes in the Umbo-larvae stage.
RpMR1 recombinant protein by prokaryotic expression
RpMR1 recombinant protein was successfully expressed in PET-28a (+) (Fig. S8). The purified RpMR1 recombinant protein was obtained with a concentration of (0.5 mg/mL). After purification, a band appeared at the corresponding position of theoretical 123 kDa by SDS-PAGE electrophoresis analysis, indicating that the protein was successfully purified (Fig. 4A). Western blot analysis confirmed the successful expression of the target protein (Fig. S9).Fig. 4SDS-PAGE electrophoresis of RpMR1 recombinant protein. (A) Note: M: Protein Marker; 1: uninduced sample; 2-6: induced samples. (B) The effect of RpMR1 on the survival rate of R. philippinarum infected by V. anguillarum. Note: A: PBS + RpMR, B: V. anguillarum + RpMR, C: V. anguillarum + PBS and D: PBS. Use letters to indicate significant differences at different time points, with an asterisk indicating differences between different groups at the same time point (P < 0.05)
Survival rate of clam after V. anguillarum infection with injection of RpMR1 recombinant protein
To confirmed the function of recombinant protein RpMR1, survival rate of clam after V. anguillarum infection with injection of RpMR1 was performed. Each experimental group had 150 clams, divided equally into 3 groups, with 50 clams in each group. As shown in Fig. 4, the cumulative mortality rate of group C (V. anguillarum + PBS) was 78.6% at the 96 h of the experiment, and the cumulative mortality rate of group B (V. anguillarum + MR) was 52.7%. Group C has a cumulative mortality rate of 15.9% higher than Group B. R. philippinarum in group A (PBS + MR) and group D (PBS) showed the same mortality rate (Fig. 4B). It indicated that the RpMR1 recombinant protein may enhance the ability of clams to resist V. anguillarum.
Effects of RpMR1 protein injection on TLR pathway gene expression and nitric oxide synthase activity in R. philippinarum
Expression of TLR pathway-related genes after injection of RpMR1 recombinant protein was shown in Fig. 5. The results showed that the gene expressions of TLR, MyD88, TRAF, NF-κB, IKK, and AP-1 in the (Va + MR) group peaked at 6 h of post infection (P < 0.05). The expression of TLR, TRAF, and NF-κB in the (PBS + MR) group peaked at 6 h and the amount reached the peak, and MyD88, IKK, AP-1 reached the highest peak at 48 h. Most TLR pathway genes in (Va + PBS) group peaked at 96h. The nitric oxide synthase NOS activity of (Va + MR) group was significantly increased at 12 h and 72 h (P < 0.05), and was significantly higher than that of (PBS + MR) group and (Va + PBS) group (P < 0.01) (Fig. 6).Fig. 5. Expression of Toll-like receptor pathway related genes after injection of RpMR1 recombinant protein. Note: Different lowercase letters indicate that there are significant differences in gene expression at different time points (P < 0.05)Fig. 6NOS activity after injection of RpMR1 recombinant protein. Note: Different letters indicate the difference of the same treatment at different time points (p < 0.05)."*"indicates extremely significant difference group at the same time point (p < 0.01)
RpMR1 protein antibacterial activity
In vitro antibacterial activity assays of the Ruditapes philippinarum recombinant protein RpMR1 were conducted (Fig. 7). The RpMR1 protein exhibited significant inhibitory effects against several Gram-negative bacteria, including Vibrio anguillarum, V. splendidus, and V. alginolyticus. Significant growth inhibition was observed against V. splendidus at 4 h (P < 0.05), with sustained suppression of V. anguillarum at 6 h, 8 h, and 10 h (P < 0.05). However, no inhibitory activity was detected against Bacillus subtilis, Staphylococcus aureus, Vibrio parahaemolyticus, V. harveyi, or Escherichia coli.Fig. 7. The inhibition curve of RpMR1 protein on the growth of different bacteria was determined using. Note: Ampicillin as a positive control and PBS as a negative control"*"p < 0.05
Changes of RpMR1 and TLR pathway gene expression levels in different
Following RNA interference (RNAi) targeting the RpMR1 gene, its expression was significantly reduced (P < 0.01). Concurrently, key genes in the TLR signaling pathway were also downregulated: TRAF6 expression decreased significantly (P < 0.01), while TLR4 and AP-1 expression levels were significantly reduced (P < 0.05) (Fig. 8). These results suggest that RpMR1 positively regulates components of the TLR pathway during the immune response (Fig. 8).Fig. 8. Changes of gene expression levels in different experimental groups by injecting RNA interference probes. A is the change of MR expression, B is the change of TLR pathway gene expression. Note: The RNAi group was injected with 25 μg/100 μl of synthetic dsRNA RpMR interference strand, and the negative control group was injected with 25 μg/100 μl of dsRNA-pEGFP-N3 interference strand. The positive control group was injected with 100 μl of DEPC. Con is no treatment control group."*"indicate significant differences between genes after MR interference and the control group (p < 0.05),"**"(p < 0.01)
Discussion
The RpMR genes contain conserved aromatic amino acids, including phenylalanine (Phe) and tryptophan (Trp). Studies have shown that these two amino acids are critical for the MR gene to bind to collagen types I, II, III, and IV in the extracellular matrix (Zheng et al. 2015). The MR gene can degrade and metabolize endogenous glycoproteins to maintain body stability, while enzymes released from inflammatory sites are recognized by the CLECT domain, which can further decompose and clear them to alleviate inflammatory responses (Zheng et al. 2015). Structural domain analysis revealed that all RpMR genes contain CLECT domains, indicating that the immune response of clams induced by MR genes may be inseparable from the function of CLECT domains. In this study, we identified 13 mannose receptor genes (RpMR1–13) in R. philippinarum through genome-wide analysis of publicly available data (BioProject PRJNA479743). Integrated transcriptomic datasets (BioProject PRJNA738278) revealed that six genes (RpMR1–6) exhibited differential expression following V. anguillarum challenge. The remaining genes (RpMR7–13) showed no detectable expression post-infection and were excluded from downstream analyses. We hypothesize that the absence of RpMR7–13 transcripts may stem from either: (1) simplified domain architectures reducing functional necessity and mRNA instability (Ibrahim et al., 2020), or (2) excessively complex domain structures hindering transcriptional coordination (Bickmore et al., 2013; Dong et al. 2024), collectively limiting their involvement in antimicrobial immune responses.
According to transcriptomic analysis of clam resistance to V. anguillarum stress, we conclude that the RpMR gene plays a critical role in the clam's resistance to this pathogen (Dong et al. 2024). In some species, such as P. trituberculatus and C. idella, the full-length of MR gene has been cloned and its immune characteristics analyzed (Wang et al. 2014; Zhang et al. 2019). Functional studies in P. clarkii have demonstrated that the mannose receptor (MR) gene critically determines antimicrobial capacity. MR knockdown experiments revealed severe impairment of antibacterial defenses (P < 0.05), directly linking its activity to pathogen resistance (Man et al. 2018). Downregulation of MR disrupts immune signaling pathways, suppressing inflammatory cytokine production and weakening host antimicrobial responses. Furthermore, MR deficiency impairs antigen presentation, attenuating T cell activation and adaptive immunity against Leishmania infection. These findings highlight the MR play important roles in innate and adaptive immunity (Gazi and Martinez-Pomares 2009). The immunological importance of MR genes has been reported in vertebrates and crustaceans, such as C. idella, P. clarkii, and E. coioides (Cano et al.,2024; Man et al. 2018; Wang et al. 2014; Zhang et al. 2023, 2021). Our study characterized the immune function of the RpMR gene in R. philippinarum following V. anguillarum challenge. Tissue-specific expression analysis revealed predominant RpMR expression in the hepatopancreas, with detectable levels in other tissues. This expression pattern parallels the conserved immunological roles of mannose receptors documented in D. rerio, L. hoevenii, and P. clarkii (Gazi and Martinez-Pomares 2009; Man et al. 2018; Zheng et al. 2015).
In this study, mortality analysis revealed that intramuscular administration of recombinant RpMR1 protein significantly reduced R. philippinarum mortality by 26% following V. anguillarum challenge (P < 0.05), demonstrating its potential to enhance anti-Vibrio immunity in clams. Complementary studies in M. amblycephala have established that the mannose receptor (MR) interacts with chitooligosaccharides (COS) to modulate nitric oxide synthase (NOS) expression (Ouyang et al. 2021). Importantly, our findings align with recent evidence confirming NOS as a critical component of phagocytic machinery, where it facilitates pathogen clearance through reactive nitrogen species production (Yin et al. 2022b). Emerging evidence establishes nitric oxide (NO) as a pivotal mediator of neutrophil immunity, where inducible nitric oxide synthase (iNOS)-derived NO promotes phagolysosome maturation and enhances microbicidal activity (Kumar et al., 2024). Intriguingly, our experimental data revealed that the NOS activity in the group injected with recombinant RpMR1 protein was significantly increased at 12 h post-V. anguillarum stress. This investigation provides the first functional evidence of MR-dependent regulation of NO synthase pathways in bivalve immunity, bridging receptor-mediated recognition with downstream effector mechanisms in molluscan host defense. Our findings elucidate the critical role of the MR in enhancing phagocytic engulfment of V. anguillarum by hemocytes. However, the precise molecular interplay by which RpMR1 confers anti-Vibrio immunity through NOS-dependent mechanisms remains to be delineated, necessitating targeted investigations to unravel this tripartite interaction between MR activation, effector enzyme regulation, and pathogen clearance.
The antimicrobial efficacy of mannose receptor (MR) recombinant protein against Gram-negative bacteria has been well documented (Gazi and Martinez-Pomares 2009; Zheng et al. 2015). While our cell-free in vitro assays exclude cellular components, RpMR1 demonstrates bacterial-binding capacity through PAMP recognition, forming immune complexes that suggest dual mechanisms: 1) priming phagocyte activation via opsonin-like effects, and 2) potentiating complement-dependent bacteriolysis. These findings provide mechanistic insights for future investigations into receptor-mediated immune potentiation in vivo (Huo et al. 2024a). Our study corroborates the antimicrobial specificity of recombinant RpMR1 protein, demonstrating significant growth inhibition against Gram-negative Vibrio pathogens, including V. anguillarum, V. splendidus, and V. alginolyticus. This bactericidal activity likely stems from RpMR1's affinity for lipopolysaccharides (LPS), the dominant outer membrane component of Gram-negative bacteria. Notably, LPS serves as a conserved PAMP that is specifically recognized by *TLR4 *to initiate MyD88-dependent signaling cascades—a critical immune activation pathway conserved from invertebrates to mammals (Nunes-Alves 2014). It has been demonstrated that the main mechanism of MR recognition and binding to pathogens is due to the presence of phagocytic cells on the surface, which activates innate immunity through phagocytosis and simultaneously activates pro-inflammatory signaling pathways (Zheng et al. 2015). However, emerging evidence suggests that MR lacks intrinsic signaling capacity, requiring cooperative interactions with complement receptor 3 (CR3) and Fc receptors to transduce immune activation signals. This receptor clustering mechanism ensures precise control over inflammatory responses while maintaining immune homeostasis (Balderramas et al., 2014; Xaplanteri et al. 2009; Zheng et al. 2015). Cytokine profiling studies reveal a synergistic interaction between MR and TLR4 in orchestrating pro-inflammatory mediator release, particularly IL-1β, TNF-α, and *IL-6 *(Zhang et al. 2005; Zheng et al. 2015). Mechanistically, MR expression in dendritic cells modulates Th2 cell polarization through C-type lectin receptor (CLR)-mediated antigen presentation. Crucially, TLR4 signaling serves as a critical modulator of this immunoregulatory axis, with stromal cell-specific TLR4 activation being indispensable for initiating allergen-induced Th2 immunity against inhaled particulates (Li et al. 2010; Tan et al. 2010). Our results show that the expression of TLR4 gene decreased significantly after injection of RpMR dsRNA, demonstrating the potential interaction between MR and TLR in innate immune response. In addition, we determined the gene expression of TLR signaling pathway after injection of V. anguillarum and RpMR1 recombinant protein, with most of the genes in the TLR pathway reaching peak expression at 6 h in the early stage of infection. Mechanistic insights suggest that RpMR1 recombinant protein activates TLR signaling pathways through PRRs, triggering rapid antimicrobial responses against V. anguillarum infection. The immunological functions of RpMR1 recombinant protein are anticipated to be applied in the aquaculture of R. philippinarum, where it can be incorporated into feed to serve as an antimicrobial agent. However, for large-scale application, further research is needed to determine the optimal dosage of the recombinant protein and its capacity to maintain sustained antibacterial activity.
Conclusions
In this study, we investigated the function and mechanism of action of mannose receptors in R. philippinarum against V. anguillarum for the first time. The results showed that recombinant RpMR1 protein significantly reduced clam mortality following V. anguillarum infection. Recombinant RpMR1 protein exhibited inhibitory effects on Gram-negative bacteria, including V. splendidus, V. alginolyticus, and V. anguillarum. Furthermore, the protein rapidly activated the TLR signaling pathway, enhancing immune responses through increased nitric oxide synthase (NOS) activity. qPCR analysis revealed that dsRNA-RpMR1 injection suppressed the expression of both RpMR1 and TLR4 genes after V. anguillarum challenge, indicating a critical regulatory relationship between these immune components. These findings highlight the pivotal role of the mannose receptor in R. philippinarum's defense against V. anguillarum, underscoring its importance in bivalve immune defense mechanisms.
Supplementary Information
Supplementary Material 1: Fig. S1. The intron and exon information of RpMR gene family.Supplementary Material 2: Fig. S2. The distribution of conserved motifs of RpMR protein.Supplementary Material 3: Fig. S3. The evolutionary relationship and protein domain analysis of RpMR protein.Supplementary Material 4: Fig. S4. Multiple alignment of RpMR1-13. Red rectangle is labeled as conserved cysteine residue, green rectangle as aromatic amino acid Phe and Trp, blue rectangle as Ca^2+^ binding site, yellow rectangle as conservative acid modified amino acid residue.Supplementary Material 5: Fig. S5. The chromosomal localization of RpMR gene.Supplementary Material 6: Fig. S6. Phylogenetic tree of MR protein sequences from 9 species.Supplementary Material 7: Fig. S7. Heat map of RpMR gene expression at different developmental periods. Note: Fertilized egg (FE), 1 st polar body (PB1), 2 st polar body (PB2), 2-cell (TC), 8-cell (EC), blastula (B), Gastrula (G), Trochophora (T), D larva (D), Umbo veliger (U), Pediveliger (P), Single pipe juvenile (S), and Juvenile (J).Supplementary Material 8: Fig. S8. The restriction enzyme map of recombinant plasmid (Digested with NdeI-XhoI).Supplementary Material 9: Fig.S9. The Western blot method for detecting target proteins.Supplementary Material 10.Supplementary Material 11.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cano I, Blaker E, Hartnell D, Farbos A, Moore KA, Cobb A, Santos EM. Transcriptomic Responses to Koi Herpesvirus in Isolated Blood Leukocytes from Infected Common Carp. Viruses. 2024;16 (3):380.10.3390/v 16030380 PMC 1097427738543746 · doi ↗ · pubmed ↗
- 2Wang X, Liu Y, Li Z, Gao X, Dong J, Zhang J, Zhang L. Genome-Wide Identification and Expression Profile Analysis of the Phospholipase C Gene Family in Wheat (Triticum aestivum L.). Plants (Basel). 2020;9 (7):885.10.3390/plants 9070885 PMC 741211532668812 · doi ↗ · pubmed ↗