Visual physiology of Australian stingless bees
Bhavana Penmetcha, Laura A. Ryan, Yuri Ogawa, Nathan S. Hart, Ajay Narendra

TL;DR
This paper compares the visual abilities of two Australian stingless bee species, finding differences in contrast sensitivity that may relate to their foraging behaviors.
Contribution
The study provides new insights into the visual physiology of Australian stingless bees using pattern electroretinography.
Findings
Austroplebeia australis has higher contrast sensitivity (13.07) compared to Tetragonula carbonaria (5.99).
A. australis has more and larger ommatidial facets than T. carbonaria.
The two species do not differ in spatial resolving power (0.53 cycles deg−1).
Abstract
Stingless bees engage in a range of visually guided behaviours that require relatively high spatial resolution and contrast sensitivity. Although the eyes of honeybees, bumblebees, carpenter bees, and sweat bees have been studied extensively, there is limited knowledge of stingless bees. Here, we studied two sympatric Australian species, Tetragonula carbonaria and Austroplebeia australis, which are important crop pollinators. The bigger A. australis had more and larger ommatidial facets compared to T. carbonaria. Using pattern electroretinography, we showed that A. australis had higher contrast sensitivity (13.07) compared to T. carbonaria (5.99), but their spatial resolving power did not differ (0.53 cycles deg−1). We discuss these differences in visual physiology in the context of the distinct foraging behaviours of the two species.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
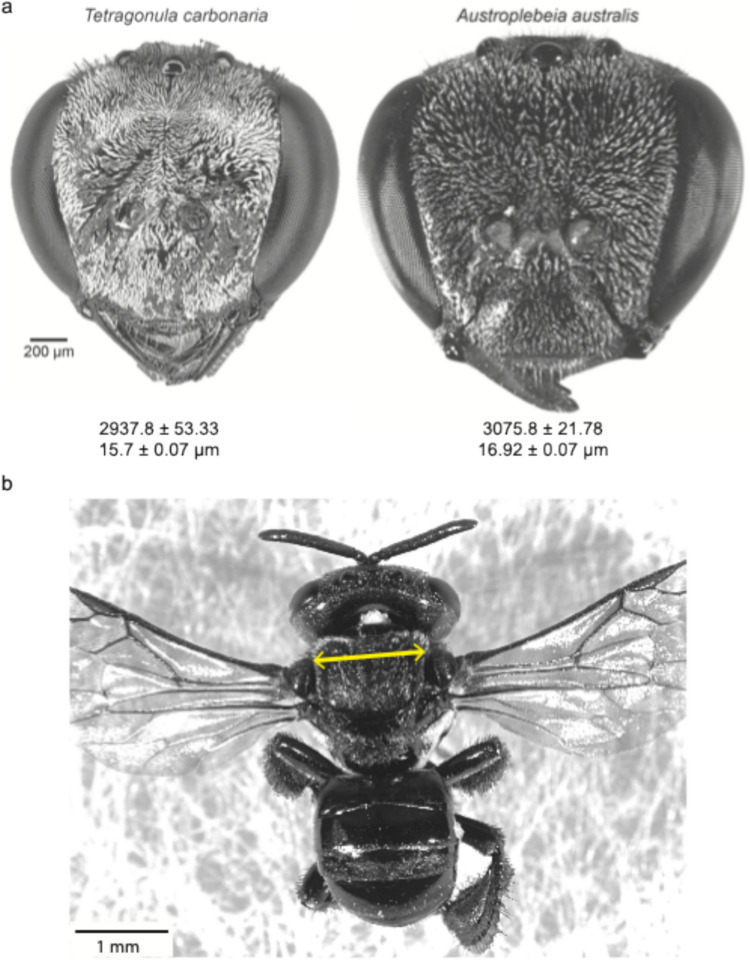 Figure 1
Figure 1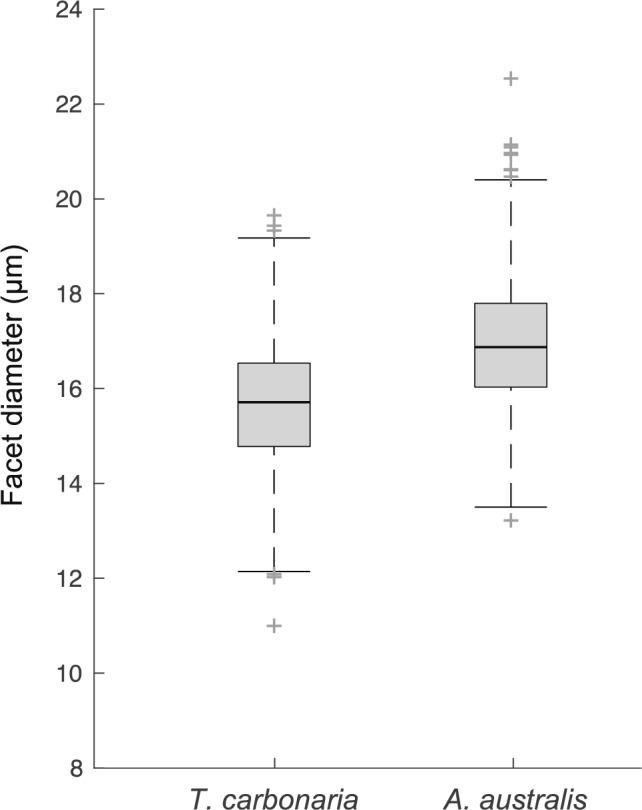 Figure 2
Figure 2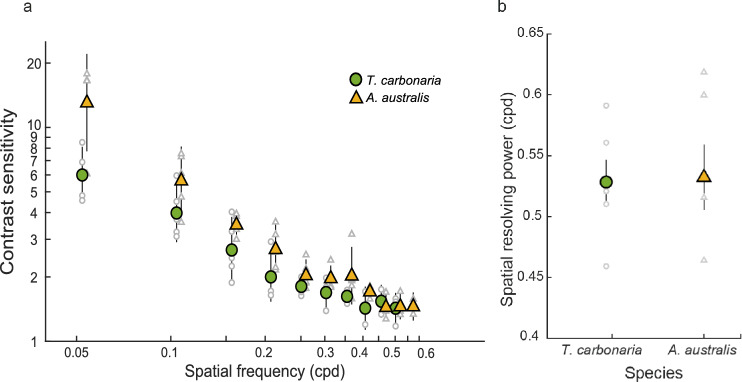 Figure 3
Figure 3- —http://dx.doi.org/10.13039/501100000923Australian Research Council
- —Macquarie University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and animal studies · Insect and Arachnid Ecology and Behavior · Neurobiology and Insect Physiology Research
Introduction
Vision is a fundamental sensory modality used by insects for foraging, sexual selection, navigation and communication. Two significant visual capabilities for an insect are spatial resolving power and contrast sensitivity. Spatial resolving power is the ability to distinguish two closely spaced points in its visual field which enables insects to resolve fine details in a natural scene. Contrast sensitivity is a measure of the ability to discriminate objects as their achromatic contrast decreases (O’Carroll and Wiederman 2014; Ogawa et al. 2019). These properties are dictated by the physiological and anatomical characteristics of the visual system. For instance, insect compound eyes with larger facets have improved sensitivity due to increased light gathering ability, which in turn improves their ability to distinguish changes in contrast (Kirschfeld 1974). Conversely, smaller facets may reduce optical sensitivity, but can be associated with smaller interommatidial angles, leading to increased spatial resolving power (Warrant and McIntyre 1993). Some insect eyes employ physiological strategies such as spatial summation where neurons in the lamina (the first optic ganglion in the insect brain) pool signals from the photoreceptors to improve sensitivity (Kirschfeld 1974; Warrant 1993; Nilsson and Ro 1994; Stöckl et al. 2016). Temporal summation is another physiological strategy whereby the integration time of the photoreceptors is extended to improve photon capture and enhance contrast discrimination, albeit at the cost of reduced temporal resolution (Frederiksen et al. 2008; Warrant 1999). These diverse strategies mean that the visual system of an animal often represents a trade-off between visual capabilities, such as sensitivity and spatial resolution, that are dictated by eye anatomy and physiology, and constrained by factors such as body size, lifestyle and visual ecology (Ogawa et al. 2022; Warrant and McIntyre 1993; Warrant 1999; Narendra et al. 2011; Somanathan et al. 2009b; Palavalli-Nettimi et al. 2019).
Bees are visually guided animals. Individual bees that exit their hive for the first time or from a newly discovered food source, perform exquisitely choreographed learning flights, where they fly out, turn around and face the hive while retreating in a series of consecutive arcs that are roughly centred around the point of interest (e.g. Da Silva et al. 1972; Lehrer 1991; Zeil et al. 1996; Capaldi et al. 2000; Robert et al. 2016, 2018; Reyes et al. 2019; Collett and Hempel de Ibarra 2023). These flights are thought to allow bees to learn visual features around the goal which they rely on subsequent trips (Cartwright and Collett 1987; Zeil and Wittmann 1993; Collett et al. 2013). Foraging bees must also cope with varying degrees of environmental ‘clutter’ and must employ strategies to avoid collisions with stationary and moving obstacles. Bees process the motion of images on their retina (i.e. optic flow) and translate this image motion into object range, thus relying on active vision strategies to perceive their three-dimensional world (Srinivasan et al. 1991; Hrncir et al. 2003; Eckles et al. 2012; Srinivasan 2021; Egelhaaf 2023). Bees use this information along with visuo-motor strategies, to control their flight speeds, stabilise gaze, detect obstacles and perform rapid flight manoeuvres to prevent collisions (e.g. Ings and Chittka 2008; Srinivasan and Zhang 2004; Boeddeker and Hemmi 2010; Baird and Dacke 2012; Burnett et al. 2020; Baird et al. 2021; Sathyakumar 2021; Ravi et al. 2022; Goyal et al. 2022). They use similar strategies to negotiate gaps and move safely between obstacles (e.g. Srinivasan et al. 1991; Baird and Dacke 2016; Ravi et al. 2019, 2020). In addition to being able to discriminate between colours (Giurfa et al. 1996; Hori et al. 2006; Spaethe et al. 2014; Hempel de Ibarra et al. 2014; Koethe et al. 2016), bees also recognise patterns and shapes that enables them to approach specific feeding sites or flowers (Dafni et al. 1997; Dyer et al. 2008, 2016; Hempel de Ibarra and Vorobyev 2009; Lunau et al. 2009; Sánchez and Vandame 2012). Back at their hive, bees use visual cues to defend their hive from intruders (Wittmann et al. 1990; Kelber and Zeil 1990; Koeniger et al. 2017). To carry out such behaviours, bees require a compound eye that has relatively high spatial resolution and good ability to detect contrast differences.
Several aspects of an insect’s visual physiology are correlated with size. Bees exhibit dramatic variation in body size varying from 1.8–38.0 mm in body length (Everaars 1979; Kelber and Somanathan 2019). Smaller bees tend to have smaller compound eyes—and thus fewer ommatidial lenses—which results in reduced spatial resolution and optical sensitivity (Kelber and Somanathan 2019; Ribi et al. 1989; Spaethe and Chittka 2003; Somanathan et al. 2009a, b). In bumblebees, smaller workers have lower spatial resolution (interommatidial angles: 1.2° vertical and 2.9° horizontal), compared to larger workers (0.9° vertical and 2.1° horizontal; Spaethe and Chittka 2003). In other Hymenopterans, such as ants, species with fewer and smaller ommatidial lenses have reduced contrast sensitivity; however, their spatial resolving power is comparable to larger species (Palavalli-Nettimi et al. 2019).
Most research on the visual system of bees has focused on honeybees (Apis), bumblebees (Bombus), carpenter bees (Xylocopa) and sweat bees (Megalopta) (Chakravarthi et al. 2016, 2018; Ribi et al. 1989; Spaethe and Chittka 2003; Greiner et al. 2004; Somanathan et al. 2009b, a; Streinzer and Spaethe 2014). Stingless bees are one of the largest groups of bees (tribe: Melopinini), represented by over 550 species across the world (Grüter 2020). Spatial resolution has only been estimated in two stingless bee species: the Australian Tetragonula carbonaria and the South/South-East Asian Tetragonula iridipennis. Behavioural estimates of spatial resolution between the two species are comparable, at 0.053 cycles deg^−1^ (cpd) in T. carbonaria and 0.054 cpd in T. iridipennis. Anatomical estimates of spatial resolution in T. carbonaria are 0.32 cpd (Dyer et al. 2016), while theoretical estimates in T. iridipennis, are significantly lower at 0.17 cpd. Given the unexpected difference in T. carbonaria (behavioural vs anatomical estimates) and the variation between species (anatomical vs theoretical estimates), here we investigated spatial resolving power and contrast sensitivity of the compound eyes of T. carbonaria and Austroplebeia australis using pattern electroretinograms and compared this to the anatomical characteristics of the eye such as facet numbers and diameters.
Stingless bees have been an important part of indigenous Australian culture for centuries, far outdating modern meliponiculture which has become popular only in the last few decades (Heard and Dollin 2000; Halcroft 2012; Vit et al. 2013). Stingless bees such as T. carbonaria and A. australis are used as an important crop pollinator (Heard 1994, 1999; Heard and Dollin 1998; Vit et al. 2013). These two species are derived from phylogenetically different lineages (Rasmussen and Cameron 2010), that occupy overlapping ranges and exhibit distinct differences in their behaviour. T. carbonaria is a generalist and opportunistic pollen forager that invests more in resource collection, whereas A. australis tend to focus more on resource quality (Leonhardt et al. 2014). Additionally, A. australis foragers spend proportionately less amount of time hovering in front of flowers than T. carbonaria, hence are described as more ‘efficient foragers’ in terms of energy consumption (Halcroft 2012). A. australis are also active in slightly dim light conditions compared to T. carbonaria (Heard and Hendrikz 1993; Halcroft 2012). These two species therefore provide a unique opportunity to compare visual properties in a culturally and economically important species that forage in similar ecologies but differ in their foraging behaviour.
Methods
Study species
We investigated the compound eyes of two species of Australian native stingless bees: Tetragonula carbonaria Smith and Austroplebeia australis Friese (Fig. 1a). Experiments were carried out between January 2022 – February 2023, using worker bees collected from one hived colony of each species maintained at Macquarie University campus, North Ryde, NSW, Australia (33°46′10.24″ S, 151°06′39.55″ E).Fig. 1. External morphology of two Australian native stingless bees. a Gray scale dorsal view of the head of Tetragonula carbonaria and Austroplebeia australis. Average number of facets per eye (mean ± SEM) and mean facet diameters of the medio-frontal regions of the compound eyes are shown. Image of A. australis courtesy of Giovanni Ramón-Cabrera. b Dorsal view of T. carbonaria showing intertegular width measurement, highlighted in yellow
External morphology of stingless bee eyes
We used the intertegular width (ITW), the distance between the wing bases, as an estimate of the body size of A. australis (n = 5) and T. carbonaria (n = 7; Fig. 1b). To determine the number of facets and facet diameters in the compound eyes of A. australis and T. carbonaria, we used an established technique described in detail elsewhere (Ribi et al. 1989; Ramirez-Esquivel et al. 2017a, b). Briefly, we painted the surface of one eye with clear nail polish to create a cast of the facets. The cast was left to dry on the eye, removed and then flattened on a glass slide by making fine incisions. Eye casts were created for five individuals for both species and photographed under a light microscope (Leica DM5000B, Leica Microsystems GmbH, Wetzlar, Germany). For each individual, we counted the number of facets and measured the diameter of 260 randomly selected facets in the medio-frontal region of the compound eyes using ImageJ (version 1.52k) (Schneider et al. 2012). We determined the mean facet diameter of the mediofrontal region (the region with the largest facets) for each individual and from this we calculated the mean facet diameter for each species.
Animal preparation for pattern electroretinography
To investigate spatial vision in A. australis and T. carbonaria, we performed pattern electroretinograms (pERG). Bees were first anaesthetized by cooling them in an ice box for 5–7 min before mounting them. To secure the bees, they were carefully manoeuvred into a 3 mm diameter pipette tip until the head and a small portion of the thorax was exposed. The pipette mount along with the bee was fixed horizontally onto a plastic stage with the bee’s dorsal side facing upwards using removable adhesive (Blu-Tack, Bostik Indonesia Inc., Banten, Indonesia). The head of the bee was oriented approximately 45° from the mount to ensure that the compound eyes faced the stimulus, and the mandibles were fixed using beeswax to avoid movements. We removed the antennae in T. carbonaria but kept them in A. australis as this was the only way to ensure animals remained alive during the experiments. The animals were mounted within a Faraday cage wherein electrophysiological recordings were carried out. An active electrode of 0.25 mm diameter platinum wire with a sharp tip immersed in conductive gel (Livingstone International Pty Ltd., New South Wales, Australia) was positioned above the medio-frontal region of the compound eyes. A silver/silver chloride wire was inserted into the thorax of the bees, which served as an indifferent electrode, that was 0.1 mm diameter in T. carbonaria and 0.127 mm diameter in A. australis.
To reduce any effects of circadian rhythms on eye physiology, the experiments were conducted during the activity time of the species i.e., between 2 and 8 h post-sunrise.
Pattern electroretinography (pERG)
The pERG technique was used to measure the spatial resolving power and contrast sensitivity of the compound eyes of A. australis and T. carbonaria. Detailed methods are described earlier (Ogawa et al. 2019; Palavalli-Nettimi et al. 2019; Ryan et al. 2020, Penmetcha et al. 2021; Ogawa et al. 2023a, b). Briefly, the experiments were carried out in a dark room within a Faraday cage. The bees were dark adapted for 10 min prior to the first recording. We found this to be the maximum time that the bees could be dark-adapted for and remain alive for the duration of the experiment. The bees were then adapted to a uniform grey stimulus that had the same mean irradiance as the grating stimuli for 5 min.
The pERGs were amplified with a gain of × 1000 and bandpass filtered between 1 Hz and 1 kHz with a differential amplifier (DAM50, World Precision Instruments Inc., FL, USA). Voltages were digitised at 20,000 Hz using a National Instruments data acquisition device (USB-6353, National Instruments, Austin, TX, USA) controlled via custom software written in Microsoft Visual Studio (2013, Microsoft Corporation, Redmond, WA, US) which was also used to control the stimulus presentation.
Visual stimuli were projected by a digital light processing (DLP) projector (W1210ST, BenQ corporation, Taipei, Taiwan) onto a white screen (51 cm width × 81 cm height) at a distance of 30 cm from the animal. The luminance of the white screen for the projector was 1.75 × 10^−4^ W/cm^2^ measured using a calibrated radiometer (ILT1700, International Light Technologies, Peabody, MA, USA). We used vertical contrast-reversing sinusoidal gratings of 11 spatial frequencies (0.6, 0.5, 0.45, 0.4, 0.35, 0.3, 0.25, 0.2, 0.15, 0.1 and 0.05 cpd) and up to eight Michelson’s contrasts (95%, 85%, 75%, 50%, 25%, 12.5%, 6% and 3%) for each spatial frequency in descending order. Every second spatial frequency was presented in descending order, followed by the alternative spatial frequency in ascending order to assess if signal strength degraded over time. As a control, the non-visual electrical signal (background noise) was recorded at two spatial frequencies (0.05 and 0.1 cpd) at 95% contrast with a blackboard to shield the animal from the visual stimuli before and after running the experimental series. The maximum signal out of four control runs was used as the noise threshold.
The sinusoidal gratings were reversed at a temporal frequency of 2 Hz. For each combination of the stimuli, 10 repetitions of the response for 5 s each were averaged in the time domain. The averaged responses were then analysed using a Fast Fourier Transform, FFT. An F-test was used to assess whether the response signal at the second harmonic (4 Hz) of the FFT response spectrum differed significantly from 10 neighbouring frequencies, five on either side, for each presented stimulus. Spatial resolving power and contrast threshold were obtained by interpolating from the last point above the noise threshold whose FFT amplitude at 4 Hz was also significantly greater than the 10 surrounding frequencies, and the first point below the noise threshold. Contrast sensitivity is defined as the inverse of contrast threshold.
Statistical analyses
A linear model in R (R Core Team 2018) was used to test if the maximum contrast sensitivities and spatial resolving powers were different between the two species. Subsequently, we used a linear mixed-effects model to investigate whether the spatial frequency of the pERG stimulus and the species affected contrast sensitivity functions. Spatial frequency of the stimulus and the species were the fixed effects while the animal identity was a random effect. We carried out a pairwise comparison to identify the spatial frequencies at which the contrast sensitivities of the two species were different using the emmeans package in R (R Core Team 2018), with a Bonferroni correction applied.
All linear mixed effect models were carried out in the lme4 package (Bates et al. 2015) of R (R Core Team 2018) using lmer with the restricted maximum likelihood (REML) estimation method. The Akaike Information Criterion function in R (R Core Team 2018) was used to find the best fit for all regression models and a value of 10 was used as the cut-off. The significance of the fixed effect terms was examined using the t-test with Satterthwaite approximation for degree of freedom (lmerTest package). In all instances mentioned above, the model assumptions such as linearity were tested by plotting the residuals against the fitted values of the model.
Results
External morphology of A. australis and T. carbonaria eyes
Foragers of A. australis were slightly larger with an ITW of 1.27 ± 0.02 mm (mean ± SEM), compared to T. carbonaria which was 1.15 ± 0.02 mm (Fig. 1b; Table 1). Facet numbers varied between the two species, with approximately 5% more facets in A. australis (3075.8 ± 21.78; mean ± SEM) compared to T. carbonaria (2937.8 ± 53.33). Facet diameters also varied between the two species with slightly larger facets in A. australis (range: 13.2–22.51 µm) compared to T. carbonaria (11.04–19.69 µm; Figs. 1a, 2; Table 1). Table 1. External morphology and visual properties of Australian stingless bees and European honeybeeMeasurement (mean ± SEM)SpeciesA. australisT. carbonariaA. mellifera (n = 5)Intertegular width (mm)1.27 ± 0.02(n = 5)1.15 ± 0.02(n = 7)3.2*Mean number of facets3075.8 ± 21.78(n = 5)2937.8 ± 53.33(n = 5)5311 ± 186^†^(n = 5)Mean facet diameter (µm)13.2–22.51(minimum–maximum)11.04–19.69(minimum–maximum)18.8 ± 0.6^†^(30 facets/individual)Spatial resolution (cpd)0.53 ± 0.03(n = 5)0.53 ± 0.02(n = 5)0.54 ± 0.02^†^(n = 5)Maximum contrast sensitivity (at 0.05 cpd)13.07 ± 1.17 (7.65%)(n = 5)5.99 ± 1.09 (16.68%)(n = 5)16.92 ± 3.0^†^ (6%)(n = 5)Data for A. mellifera from *Somanathan et al. 2009b and ^†^Ryan et al. 2020Fig. 2. Facet diameters of two Australian native stingless bees. Boxplots shows the diameter of all the measured facets in T. carbonaria and A. australis. Plus ( +) symbol indicate outliers
Spatial resolving power and contrast sensitivity of A. australis and T. carbonaria
The spatial resolving power of the two species did not differ (A. australis: 0.53 ± 0.03 cpd; mean ± SEM; T. carbonaria: 0.53 ± 0.02; Fig. 3b; Table 1; Linear model: F(1,8) = 0.002, p = 0.96). As the spatial frequencies of the stimulus increased, the contrast sensitivity decreased in both species (Fig. 3a; Table 2). The maximum contrast sensitivity attainted at the lowest spatial frequency (0.05 cpd) was significantly higher in A. australis at 13.07 ± 1.17 (7.65%; mean ± SEM) than in T. carbonaria at 5.99 ± 1.09 (16.68%; Fig. 3a; Table 1; Linear model: F(1,8) = 11.42; p = 0.009). In A. australis, the contrast sensitivity was significantly higher at all spatial frequencies except at 0.26 and 0.31 cpd (Fig. 3a; Table 2; T. carbonaria: 0.23 cpd, A. australis: 0.23 cpd; p = 0.006).Fig. 3. Contrast sensitivity (a) and spatial resolving power (b) of the compound eyes of the two Australian native stingless bees, T. carbonaria and A. australis (n = 5 for both species). In a each coloured data point is the mean contrast sensitivity of all individuals of a particular species at the corresponding spatial frequency. The error bars show 95% confidence intervals. Individual data points are shown in grey. Data points for A. australis were shifted to the right of the recorded spatial frequency to improve visualisation. All contrast sensitivity values plotted on a log scale. In b each coloured data point is the mean spatial resolving power of all individuals of a particular species at 95% contrast. Error bars (black lines) show standard error (SEM). Individual data points are shown in greyTable 2Summary of linear mixed-effects model to test the relationship between the contrast sensitivity function, spatial frequency of the grating stimulus and the species testedParameterEstimateStandard errorDegrees of freedomt-Valuep-ValueIntercept−0.090.0453.59−2.470.02Spatial frequency−0.660.0583.09−14.64 < 2 \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${e}^{-16}$$\end{document} A. australis−0.060.0547.11−1.160.25Spatial frequency:* A. australis*−0.260.0683.26−4.226.31 \documentclass[12pt]{minimal} \usepackage{amsmath} \usepackage{wasysym} \usepackage{amsfonts} \usepackage{amssymb} \usepackage{amsbsy} \usepackage{mathrsfs} \usepackage{upgreek} \setlength{\oddsidemargin}{-69pt} \begin{document}$${e}^{-5}$$\end{document} Model: contrast sensitivity ~ spatial frequency * species + (1 | individual identity). The t-tests for fixed effects use Satterthwaite approximations to degrees of freedom
Discussion
We investigated the spatial properties of the compound eyes in two Australian native stingless bees. A. australis were larger in body size and had more and larger facets compared to T. carbonaria. The spatial resolving power of the two species did not differ significantly; however, the larger A. australis, had higher contrast sensitivity compared to the smaller T. carbonaria.
Spatial resolving power of compound eyes of bees
Compared to the two species of stingless bees studied here, the honeybee A. mellifera has almost 1.5 times more facets. However, the spatial resolving power of both the stingless bee species (0.53 cpd; Fig. 3b, Table 1) is comparable to A. mellifera (0.54 cpd; Table 1; Ryan et al. 2020). The methods used to study stingless bees and honeybees were identical, which makes these results directly comparable. By comparison, in ants, where the same pERG technique was used, spatial resolving power decreased with the number of facets, although this relationship was not linear. The smallest ants that had only 8% of the facets compared to the large-eyed ants, still had 80% of the spatial resolution compared to the largest animals (Palavalli-Nettimi et al. 2019). A similar trend appears among the bees in this study. Bees, owing to their flying mode of locomotion have larger eyes and more facets per eye compared to worker ants that lead a pedestrian lifestyle. It is interesting that species with such differences in their external sensory array and lifestyles have similar spatial resolution. The variation in eye anatomy of bees is important, as it influences their contrast sensitivity which is maximised to suit their lifestyle and body size limitations.
Bees of the genus Tetragonula are the only stingless bees in which spatial resolution has been estimated using theoretical techniques (e.g., µCT, tracking pseudopupil changes) and behavioural methods (single target detection ability) (Kelber and Somanathan 2019). The theoretical estimate of spatial resolution of T. iridipennis is 0.17 cpd, and the behavioural estimate is 0.04 cpd (chromatic contrast only) and 0.054 cpd (chromatic + achromatic contrast). The theoretical estimate of spatial resolution of T. carbonaria is 0.3 cpd (e.g., pseudopupil changes), and the behavioural estimate is 0.053 cpd (both chromatic only and chromatic + achromatic contrast). Behavioural and theoretical estimates of spatial acuity typically do not match in A. mellifera (Ryan et al. 2020). The discrepancies in the estimates of spatial resolving power from pseudopupil illumination and the pERG are potentially due to the differences in the technique. Pseudopupil tracking method measures the axial directions of ommatidia whereas pERG relies on extracellular signals from the lamina where both temporal and spatial summation strategies may occur. The behavioural estimates of spatial acuity in T. carbonaria suggest that bees detect a 1 cm diameter flower only when they are 6 cm from the target. Although, in natural outdoor experiments, bees detect thin silk strands of spider webs at this distance (Sathyakumar 2021), which suggests a higher behavioural spatial acuity is required. It would be thus useful to assess their spatial resolution with behaviours such as object detection or avoidance, which require high spatial resolution.
Contrast sensitivity of compound eyes of bees
Contrast sensitivity is the ability to discriminate patterns as their brightness contrast decreases (Land 1997). pERG measurements showed that at the lowest spatial frequency of 0.05 cpd the maximum contrast sensitivity was 13.07 (7.65% contrast) in A. australis and 5.99 (16.68%) in T. carbonaria (Fig. 3a, Table 1). In addition to having higher contrast sensitivity, A. australis bees also have more and larger facets (Figs. 1a, 2, Table 1). At present, we lack anatomical data on these stingless bee species. If the rhabdoms of A. australis are wider and the focal length is shorter compared to T. carbonaria, it would account for their increased optical sensitivity and improved contrast sensitivity. Indeed, day-active bees (Tetragonula iridipennis) and ants (Temnothorax rugatulus), with short focal lengths have higher than expected optical sensitivity that suggests they may also have higher contrast sensitivity (Ramirez-Esquivel et al. 2017a; Jezeera et al. 2021). So, why do A. australis require higher contrast sensitivity compared to T. carbonaria? While there are no studies that have specifically addressed this, there are a few possibilities based on behavioural observations. A. australis are active at low light intensities of up to 5.8 W m^−2^ (Halcroft 2012) compared to T. carbonaria that stop activity at 15 W m^−2^ (Heard and Hendrikz 1993). Other Hymenoptera such as ants that forage in low light also tend to have increased contrast sensitivity (Ogawa et al. 2019). Interestingly, A. australis exhibited similar contrast sensitivity to A. mellifera despite having significantly fewer facets and being considerable smaller in body size (Ryan et al. 2020). This suggests that a high contrast sensitivity is relatively important for A. australis, and perhaps other visual tasks such as colour discrimination or temporal resolution may be more important for T. carbonaria. A higher contrast sensitivity could enable A. australis to discriminate between many different shades and colours of petals. Analyses of the collected pollen suggests that A. australis gather pollen from a relatively narrow flower colour gamut compared to T. carbonaria (Leonhardt et al. 2014). It remains to be tested whether A. australis detect spatial patterns more efficiently compared to T. carbonaria.
In conclusion, Australian native stingless bee A. australis have slightly larger body size, have more and larger facets and higher contrast sensitivity in their compound eyes compared to T. carbonaria. These differences did not affect their spatial resolving power, which was relatively high and comparable to the much larger European honeybee, indicating the significance of spatial resolution in these pollinators. Now that we know the physiological capacities of the compound eyes of stingless bees, it sets the scene to investigate the behavioural limits of visually guided behaviours in these important pollinators.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Halcroft MT (2012) Investigations into the biology, behaviour and phylogeny of a potential crop pollinator: the Australian stingless bee, Austroplebeia australis. University of Western Sydney, Ph D thesis
- 2Ogawa Y, Lochlan J, Ryan LA, Robson SKA, Hart NS, Narendra A (2023 b) The properties of the compound eye of the Weaver ant, Oecophylla smaragdina. J Comp Physiol A 209:489–49810.1007/s 00359-023-01629-7PMC 1035413737055584 · doi ↗ · pubmed ↗