Risk factor for gametocyte carriage and gametocytemia in Plasmodium vivax and Plasmodium falciparum
Minxi Li, Yang Bian, Shishao Ruan, Zifang Wu, Di Zhang, Tongyu Ma, Yaming Wu, Xiao Liu, Duo Wang, Jia Lin, Danni Pan, Wenyan Cui, Lin Wang, Haichao Wei, Xuexing Zhang, Qinghui Wang, Weilin Zeng, Zhaoqing Yang, Yaming Cao, Liwang Cui, Daniel M. Parker, Yan Zhao

TL;DR
This study identifies factors linked to the presence of malaria parasites in a transmissible stage in people in Myanmar, which could help control malaria spread.
Contribution
The study identifies specific risk factors for gametocyte carriage in Plasmodium vivax and Plasmodium falciparum at the China-Myanmar border.
Findings
P. vivax gametocyte carriage is associated with dry season, fever, and prior malaria history.
Polyclonal P. vivax infections have higher gametocyte densities than monoclonal infections.
P. falciparum gametocyte density increases with infection multiplicity and during the dry season.
Abstract
Understanding Plasmodium sexual differentiation is crucial for blocking transmission. This study identified risk factors for gametocyte carriage and gametocytemia in P. vivax and P. falciparum to inform malaria elimination strategies at the China-Myanmar border. Gametocytes and asexual parasites were microscopically detected on thick smears collected from 2011 to 2020 in Laiza Township, Kachin State, Myanmar. Mono-/polyclonality were detected by genotyping at Pvmsp3α/β for P. vivax, and Pfmsp1/2 for P. falciparum. Kulldorff’s retrospective time scan statistics tested for likely clusters of gametocyte-positive cases over time. Chi-square or Fisher’s exact tests compared proportions of gametocyte-positive cases in categorical variables. Generalized linear models assessed risk factors (year, season, demographics, clinical/parasitological features) for gametocyte carriage (logistic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
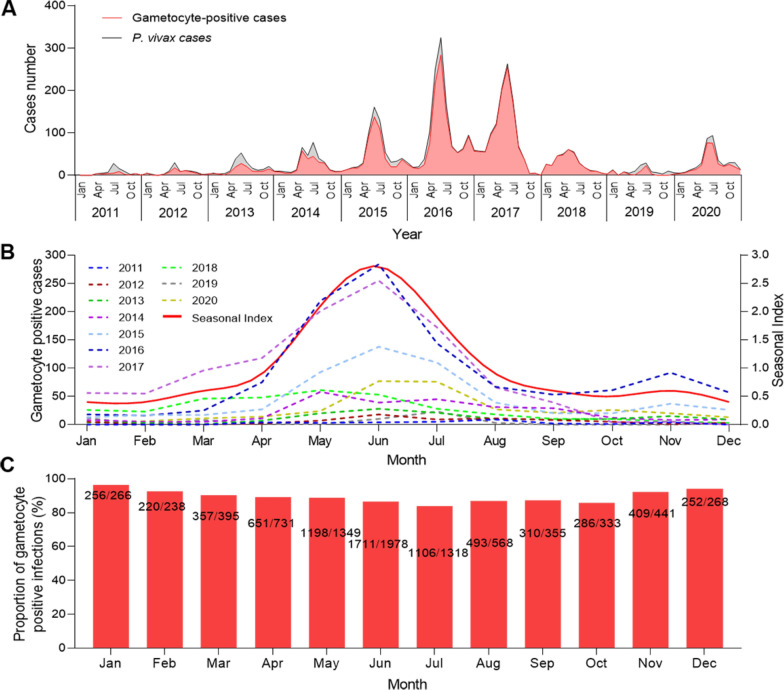 Figure 1
Figure 1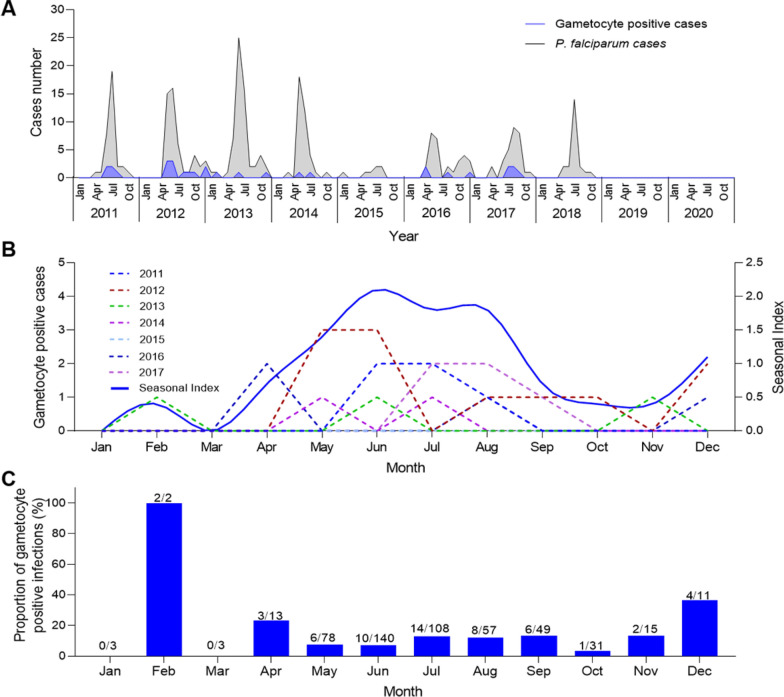 Figure 2
Figure 2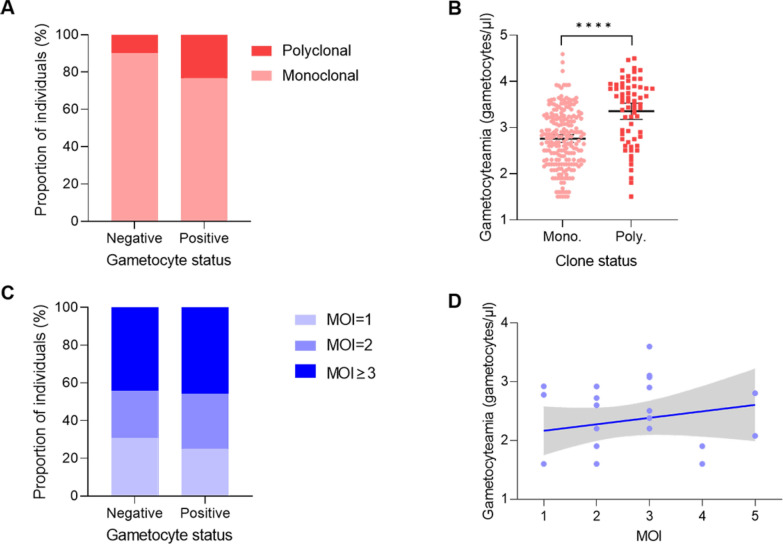 Figure 3
Figure 3 Figure 4
Figure 4- —National Institute of Allergy and Infectious Diseases, National Institutes of Health, USA
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Mosquito-borne diseases and control · Vector-borne infectious diseases
Background
Malaria, an anopheline mosquito-transmitted infectious disease with Plasmodium parasites as the pathogen, poses a serious challenge to global public health [1, 2]. The latest World Malaria Report states that there were an estimated 263 million cases of malaria and 597,000 deaths in 2023, an increase of 11 million cases compared to 2022 [3]. With increasing resistance to antimalarial drugs and insecticides, and the coronavirus disease 2019 (COVID-19) pandemic disrupting many health efforts, the public health gains from malaria efforts have stalled or even reversed. Six countries of the Greater Mekong Subregion (GMS) of Southeast Asia have set targets to eliminate malaria by 2030. However, political turmoil and conflict (especially in Myanmar) have disrupted public health efforts and led to population movement from inland malaria endemic areas to remote border areas, where people live in poverty and public health infrastructure is poor, further exacerbating the problem of sustained malaria transmission along international borders [4]. Although there are no indigenous malaria cases in Chinese mainland, increasing commercial, academic, and personnel exchanges with malaria-endemic regions such as Southeast Asia and Africa raise the risk of reintroduction. This population movement and connectivity between malarious and malaria-receptive areas (such as parts of China where malaria vectors continue to exist) make it possible for malaria to re-spread in China [5].
The human-to-mosquito transmission of malaria parasites completely depends on the sexual gametocyte stage [6]. Plasmodium falciparum gametocytes appear in peripheral blood approximately 7–15 days after the initial erythrocytic cycle, whereas those of other species (e.g. Plasmodium vivax) appear much earlier, within 1–3 days [7–9]. Unlike other species, P. vivax can form hypnozoites in hepatocytes, and its gametocytes develop well before the parasites enter the bloodstream. It has been suggested that the gametocytes of P. vivax are derived from merozoites in the liver stage, which could explain why patients infected with P. vivax can transmit without showing clinical symptoms [10–12]. The factors associated with gametocyte carriage are complex, and the mechanisms are not yet fully understood. Human genetic background, environmental factors (including climatic and meteorological factors), asexual parasitemia, allelic differences in Plasmodium strain-specific genes, and host fever or hematopoiesis may all affect sexual differentiation, sexual orientation, gametocyte maturation and viability, and ultimately transmission to mosquitoes [13, 14].
Multiplicity of infection (MOI), the number of different parasite genotypes that co-exist in a given infection, is considered a meaningful metric for describing the intensity of malaria transmission (with MOI normally thought to be higher in settings with higher transmission) [15]. In P. falciparum infections, merozoite surface protein 1 (Pfmsp1) and merozoite surface protein 2 (Pfmsp2) are commonly used as polymorphic markers to distinguish genetic variation in the parasite. Differentiating parasite genotypes in a specific infection from 1 (monoclonal infection) to 10 (polyclonal infection) can be distinguished using this method in endemic settings [16, 17]. For P. vivax, merozoite surface protein 3α (Pvmsp-3α) and merozoite surface protein 3β (Pvmsp-3β) genotyping are often used to identify the genetic diversity of the population. A positive correlation has been observed between MOI and gametocyte carriage, which in turn is associated with increased mosquito infection rates [18, 19].
Heterogeneity in the transmission of malaria has been documented at multiple scales. For instance, geographic clusters with high malaria burdens are frequently observed in many malarious regions, and these clusters can act as reservoirs for feeding persistent or recurrent malaria cases in other regions through human and mosquito movement. At more micro-scales, heterogeneities have been documented in individual susceptibility to malaria infection, exposure to mosquito vectors, and infectiousness [20–23]. In this study, we implemented passive case surveillance at a treatment facility in Laiza Township, Kachin State, Myanmar. Using data from this malarious border region we investigated parasite and host-related factors associated with gametocyte carriage and explored the extent to which specific Plasmodium variants are linked to gametocyte carriage. The results of these analyses are relevant for understanding factors influencing the natural formation of malaria gametocytes in infections, for understanding heterogeneities in potential infectiousness, and potentially for developing localized control measures aimed at preventing malaria transmission along the China-Myanmar border areas.
Methods
Study site and sample collection
The research site is located in Laiza Township (24°45′N, 97°33′E, elevation 263 m), northeast of Myanmar, adjacent to Nabang Township, Yingjiang County, Yunnan Province, China. Since 2011, the malaria investigation team of the Southeast Asia International Center of Excellence for Malaria Research (ICEMR) established a local malaria monitoring station and implemented passive case detection (PCD) in Laiza Township. In 2010, following military conflict between the local and central governments of Myanmar, some internally displaced people settled about 5 km from the Laiza town, establishing border refugee camps. From 2011 to 2017, we obtained information on malaria cases from Laiza Hospital and other clinics, including internally displaced person (IDP) camps. Since 2018, due to limited staffing resources, case data have been collected exclusively from Laiza hospital. To ensure consistency in data sources, malaria case trends from Laiza Hospital were analyzed over the period from 2011 to 2020. All available samples, including those from clinics serving the IDP camps, were used to analyze factors associated with gametocyte carriage and gametocytemia.
Patients with fever (defined as axillary temperature ≥ 37.5 °C) or suspected malaria patients with a history of fever within the past 24 h were included in the study. Demographic characteristics (ethnicity, age, sex, occupation, education level and travel history) and clinical data (body temperature, fever history, malaria history, previous treatment, use of mosquito nets, etc.) were collected using structured questionnaires. In order to determine the species and density of malaria parasites, peripheral blood was obtained through standard finger puncture methods to prepare thick and thin blood smears and dry blood spots which were stored at − 20 ℃ and used for later molecular detection experiments. All participants signed consent forms and were interviewed prior to sampling. This study was reviewed and approved by the Institutional Review Committee of the University of South Florida and China Medical University.
Microscopic analyses
For malaria diagnosis, thick and thin smears were prepared from finger-prick blood samples collected from patients, stained with Giemsa, and examined under a light microscope (LM). Initial diagnosis was performed at local hospitals or clinics by field microscopists. Patients with confirmed malaria were treated by hospital and clinic staff according to the local malaria treatment guidelines (described in [24]). All blood smears were subsequently transported to a nearby project field laboratory and re-examined by two experienced microscopists to identify Plasmodium species and quantify parasite densities of both asexual and sexual stages. Slides were double-read under microscopy, with asexual parasite densities first assessed against 200 white blood cells (WBCs). When sexual forms (gametocytes) were observed, their density was determined against 500 WBCs. A third microscopist was consulted in cases of discrepancies, defined as follows: (i) difference in Plasmodium species identification; (ii) positive versus negative results; and (iii) if the higher count divided by the lower count was ≥ 2. In such cases, the two closest values among the three readings were used to calculate the final result. Parasite density (parasites/μl of blood) was calculated by dividing the number of parasites counted by the number of WBCs observed and multiplying the result by an assumed standard of 8000 WBCs/μl. If no parasites were observed after examining 100 high-power fields at 1000 × magnification [(100 × objective) × (10 × ocular)], the result was recorded as negative [25]. The final parasite density was calculated as the arithmetic mean of the two or three readings, and per 200 WBC for asexual stage parasites or per 500 WBC for sexual stage parasites. The microscopists involved in this study were certified by the Myanmar National Malaria Control Program and had ≥ 5 years of experience in malaria diagnosis in endemic settings.
Molecular analyses
Genomic DNA (gDNA) was extracted from DBSs using the QIAamp DNA Minikit (Qiagen, Hilden, Germany) following manufacturer’s instructions. Genotyping methods for Pvmsp-3α and Pvmsp-3β have been previously described [26]. For the initial amplification, 1.0 μl of gDNA was added to a 10 μl reaction mixture containing 1.0 μl of 10 × PCR buffer, 0.8 μl of dNTPs, 0.2 μl of rTaq polymerase (Takara, Beijing, China), 0.2 μl of each primer (10 pmol/L), and distilled H₂O to reach the final volume. In nested PCR, 1.0 μl of the primary amplification product was used as the template in a total reaction volume of 10 μl. The PCR products were separated by electrophoresis on a 1.2% agarose gel and analyzed accordingly. Allelic polymorphism of the Pvmsp-3α and Pvmsp-3β loci were assessed using established polymerase chain reaction-restriction fragment length polymorphism (PCR–RFLP) techniques. Briefly, 4 μl of each PCR product was digested with restriction enzymes for one hour at 37 °C. The enzymes HhaI and PstI (Takara, Beijing, China) were used to digest Pvmsp-3α and Pvmsp-3β amplicons, respectively, in separate reactions with a total volume of 20 μl. The resulting restriction fragments were resolved on a 1.2% agarose gel and visualized accordingly. Mixed-strain infections were inferred when multiple-sized PCR products appeared during initial amplification or when the combined size of RFLP-restricted DNA fragments exceeded that of the uncut PCR product.
Nested PCR was employed to genotype the Pfmsp1 block 2 (K1, MAD20 and RO33) and Pfmsp2 block 3 (3D7 and FC27) allelic families. Briefly, the first round of PCR was performed using 1 µl of gDNA extract. The cycling conditions for the primary PCR round included an initial denaturation step of 2 min at 94 °C, followed by 30 cycles of 30 s at 94 °C, 1 min at 45 °C and 2 min at 72 °C, and a final extension step of 5 min at 72 °C. The nested PCR was performed using 1 μl of the primary PCR product as template. For the nested Pfmsp1 and Pfmsp2 rounds, the cycling conditions consisted of an initial denaturation step of 2 min at 94 °C followed by 35 cycles of 30 s at 94 °C, 1 min at 58 °C and 2 min at 72 °C, and a final extension step of 5 min at 72 °C. PCR amplicons were visualized following agarose gel electrophoresis using a GIS gel imaging system (UVP, Upland, USA). MOI was calculated separately for each locus, and the highest MOI per clinical isolate was used, as previously described in detail [16, 17].
Temporal cluster analysis
We used Kulldorff’s retrospective temporal scan statistics to identify potential clusters of vivax gametocyte positive cases over time. Briefly, the process involves scanning temporal windows and comparing the observed and expected number of cases within each window to those outside the window. A likelihood ratio test was used to assess differences in malaria risk within and outside the scanning window, under the null hypothesis of equal risk over time. The window with the maximum log likelihood ratio (LLR) was considered the most likely cluster with the highest malaria risk [27]. Temporal clustering analysis was conducted for each year from 2011 to 2020, with a temporal aggregation of one month. Additionally, the data were also analyzed for the entire study period (2011–2020). Sscan statistics were computed using SaTScan™ version 9.3 (Kulldorff M and Information Management Services, Inc., https://www.satscan.org/).
Seasonal fluctuation analysis
The seasonal index for each month was calculated by dividing the 10-year average number of cases in that month by the overall monthly mean across the entire study period, to assess seasonal variation. No substantial seasonal fluctuation would be expected if the monthly seasonal indices were all close to 1 [28].
Statistical analysis
Chi-square tests or Fisher’s exact tests were used to compare proportions of gametocyte-positive cases across categorical variables. Generalized linear models (GLM) were used to investigate the effect of season, gender, age, ethnicity, malaria history, travel history, temperature, and parasite density on gametocyte carriage using a binomial distribution (i.e., a logistic regression). The same covariates were used in a GLM with a Gaussian distribution to identify predictors of gametocytemia (continuous outcome, with gametocytes per WBC). Adjusted odds ratios (aOR) were used to quantify the magnitude of association between the factors of interest and gametocyte carriage in the logistic regression. The Wilcoxon rank sum test was used for non-parametric analyses. Standard methods were used to compute 95% confidence intervals (CI), and P values were interpreted as two-tailed. P < 0.05 indicates a statistically significant difference. All statistical analysis were performed using SPSS Statistics version 24.0 (SPSS Inc., Chicago, USA) and R software version 4.3.1 (R Core Team, R Foundation for Statistical Computing, Vienna, Austria).
Results
From January 2011 to January 2020, 46,547 questionnaires were collected from hospitals, clinics, and refugee camps. Among these, 8519 P. vivax and 542 P. falciparum cases were confirmed via microscopic examination. After excluding cases with incomplete information, a total of 8240 P. vivax and 510 P. falciparum cases were included in the risk factor analysis, with gametocyte positivity rates of 88.0% and 11.0%, respectively. Mixed infections of P. vivax and P. falciparum were excluded from the analysis due to the extremely low number of cases. Notably, Laiza Hospital was the only site that continuously collected samples throughout the entire study. From this hospital, 4580 P. vivax and 268 P. falciparum cases were identified, with gametocyte positivity rates of 86.1% and 11.2%, respectively. These cases were specifically included to analyze the trend of gametocyte positivity over time.
Temporal patterns in gametocyte carriage
First, we identified a major outbreak of P. vivax during the 10-year study period which occurred between 2015 and 2017. Notably, there was a rebound in 2020 compared with the cases in 2019 (Fig. 1A). Additionally, the trend of gametocyte-positive P. vivax cases was consistent with the overall fluctuations in malaria cases from 2011 to 2020 (Fig. 1A). Gametocyte-positive cases exhibited a distinct seasonal pattern, with the majority occurring during the rainy season (May to October), and a peak observed from May to July (Fig. 1B). The seasonal gametocyte index reached its highest level in June [seasonal index (SI) = 2.8]. The proportion of gametocyte-positive infections ranged from 83.8% in July to 96.2% in January (Fig. 1C).Fig. 1. Trend and dynamics of gametocyte carriage of Plasmodium vivax in Laiza township at the China-Myanmar border during 2011–2020. A Temporal case counts of total P. vivax infections (black line) and gametocyte-positive cases (red line). B Monthly gametocyte positive cases in different years (left y-axis) and average seasonal index (right y-axis). C Monthly proportion of gametocyte positive cases
There were relatively few cases of P. falciparum at the China-Myanmar border, with an overall downward trend in recent years. Notably, no cases of P. falciparum have been reported since 2018 (Fig. 2A). The seasonal trend showed that most cases still occurred during the rainy season, peaking from May to July (Fig. 2B). The seasonal index of gametocyte-positive cases peaked in June (SI = 2.1). Additionally, the proportion of gametocyte-positive infections was significantly higher during the dry season (November to April) than during the rainy season (May to October) (P = 0.0042, Chi-square test) (Fig. 2C).Fig. 2. Trend and dynamics of gametocyte carriage of Plasmodium falciparum in Laiza township at the China-Myanmar border during 2011–2020. A Temporal case counts of total P. falciparum infections (black line) and gametocyte-positive cases (blue line). B Monthly gametocyte positive cases in different years (left y-axis) and average seasonal index (right y-axis). C Monthly proportion of gametocyte positive cases. No gametocyte-positive cases were detected by microscopy after 2017, and P. falciparum transmission ceased after 2018
The duration of temporal clusters of gametocyte-positive cases progressively shortened over the study period, from April–August 2011 to June–July 2020 (Table 1). Temporal cluster analysis revealed that gametocyte carriage mainly occurred during the rainy season, particularly in June to July. The specific months with higher temporal clustering exhibited minor variations across different years. A particularly significant temporal cluster of gametocyte carriage was observed between May 2015 and August 2017 in the study area (LLR = 1629.69, P = 0.001) (Table 1). During this period, a total of 2613 gametocyte-positive cases were recorded, and the relative risk of malaria infection was 6.43 compared to other periods (Table 1). Table 1. Clusters of vivax gametocyte positive cases, detected using Kulldorff’s purely temporal clustering approach, in Laiza 2011–2020YearCluster time frame^a^ObservedExpectedRRLLRP-valueAll years 20112011/4 to 2011/82513.414.958.730.001 20122012/5 to 2012/95328.844.6117.520.001 20132013/5 to 2013/76934.533.0119.880.001 20142014/5 to 2014/9202108.994.8369.140.001 20152015/5 to 2015/7341140.394.68160.730.001 20162016/5 to 2016/7647279.274.15272.790.001 20172017/3 to 2017/7842447.265.19304.710.001 20182018/3 to 2018/6208110.643.3759.590.001 20192019/6 to 2019/73310.365.6721.490.001 20202020/6 to 2020/715353.174.6183.560.00110 years 2011–20202015/5 to 2017/82613922.266.431629.690.001RR relative risk; LLR log likelihood ratio^a^Clusters are identified by comparing the observed number of cases within each window to the expected number under a random distribution, while scanning across varying time intervals
Characteristics of gametocyte positivity among detected P. vivax and P. falciparum cases
All samples, including those collected from IDP camp clinics, were analyzed to identify factors associated with gametocyte carriage and gametocytemia. A total of 8240 patients with P. vivax infection were included in the study, of whom 7249 tested positive for gametocytes (Table 2). Regarding seasonal distribution, the proportion of vivax malaria cases with concurrent gametocyte carriage was significantly higher in the dry season than in the rainy season (91.7% vs 86.5%, P < 0.001). Among patients without a history of malaria, 86.4% were gametocyte-positive, which was significantly lower than the 98.1% observed among those with a reported history of malaria infection (P < 0.001). Regarding parasite density, the prevalence of gametocytes carriage was 91.4% in P. vivax infections with parasite densities > 500 parasites/μl, significantly higher than the 78.2% observed in infections with densities < 500 parasites/μl (P < 0.001) (Table 2). No statistically significant differences in gametocyte positivity were observed across categories of gender (P = 0.407), age (P = 0.088), ethnicity (P = 0.457), occupation (P = 0.062), education (P = 0.115), travel history (P = 0.295), and fever status (P = 0.962) (Table 2). Table 2. Characteristics of gametocyte carrier cases in Plasmodium vivax and P. falciparumCharacteristicsP. vivax**P. falciparumTotalGamet. + , n (%)P-valueTotalGamet. + , n (%)P-valueAll8240724951056 Season < 0.0010.004 Dry23392145 (91.7)4711 (23.4) Rainy (May to October)59015104 (86.5)46345 (9.7) Sex0.4070.165 Male49144311 (87.8)38046 (12.1) Female33262938 (88.3)13010 (7.7) Age (years)0.0880.002 < 5776691 (89.0)195 (26.3) 5–1427852473 (88.8)9216 (17.4) 15–2427822444 (87.9)21525 (11.6) 25 + 18971641 (86.5)18410 (5.4) Ethnicity0.4570.761 Non-Kachin267231 (86.5)302 (6.7) Kachin79737018 (88.0)48054 (11.3) Occupation0.0620.047 Indoor worker^a^13081134 (86.7)767 (9.2) Manual labor^b^345301 (87.2)-- Farmer1093943 (86.3)1087 (6.5) Soldier806729 (90.4)15417 (11.0) Student38043363 (88.4)14117 (12.1) Other^c^884779 (88.1)238 (29.4) Education0.1150.338 Primary or less37663312 (87.9)17724 (13.6) Middle/high school43253814 (88.2)31930 (9.4) University above149123 (83.2)142 (14.3) Malaria history^d^** < 0.001**** < 0.001** No71446174 (86.4)46423 (5.0) Yes10961075 (98.1)4633 (71.7) Travel history0.2951.000 No7199 (87.9)49254 (11.0) Yes50 (92.6)182 (11.1) Fever0.9620.083 No26222306 (87.9)11718 (15.4) Yes56184943 (88.0)39338 (9.7) Parasite density(parasites/μl)** < 0.001**0.743 ≤ 50021101649 (78.2)909 (10.0) > 50061305600 (91.4)42047 (11.2)^a^Indoor workers include office workers, housewives/housekeepers, and teachers^b^Manual labor includes factory/construction workers, lumberjack, plantation workers, temporary job/labor, hunters, miners, herdsman and gardeners/bush clearing^c^Other include non-school child and business personGamet GametocyteBold indicates a P value less than 0.05
Among the 510 patients infected with P. falciparum, 56 were positive for gametocytes. Similar to P. vivax infections, significant seasonal variation in gametocyte carriage was observed (P = 0.004), along with a significant difference between individuals with and without a history of malaria infection (P < 0.001) (Table 2). The highest proportion of gametocyte carriers was observed in children under 5 years old (26.3%), followed by those aged 5‒14 years (17.4%). Gametocyte carriage rates differed significantly across age groups (P = 0.002). Regarding occupation, the highest gametocyte carriage rate was found in non-school age children and business persons (lumped together into a category named “Other”), at 29.4%, followed by students and soldiers (12.1% and 11.0% respectively); these differences were also statistically significant (P = 0.047) (Table 2). No statistically significant differences in gametocyte positivity were found based on gender (P = 0.165), ethnicity (P = 0.761), education (P = 0.338), travel history (P = 1.000), or fever status (P = 0.083) (Table 2).
Results from logistic regression for risk factors for gametocyte carriage among vivax and falciparum malaria positive individuals
The results of the multivariable analysis revealed several statistically significant factors that influence gametocyte carriage in P. vivax infections. Firstly, seasonal factors were significantly associated with lower odds of gametocyte positivity during the rainy season compared to the dry season (aOR = 0.74, 95% CI: 0.62–0.89, P = 0.0013) (Table 3). Furthermore, patients with a history of malaria in the past 12 months had higher odds of gametocyte positivity compared to those without a previous malaria history (aOR = 17.54, 95% CI: 11.20–28.90, P < 0.001) (Table 3). Similarly, the density of Plasmodium parasites in the patients also had a statistically significant effect. Patients with high parasite density had higher odds of gametocyte positivity compared to those with low parasite density (aOR = 2.61, 95% CI: 2.31–2.95, P < 0.001) (Table 3). Compared to children under 5 years old, the odds of gametocyte carriage were lower in the 15–24 year age group (aOR = 0.75, 95% CI: 0.56–0.99, P = 0.0466) (Table 3). Body temperature (i.e., fever) was also associated with gametocyte carriage. Compared to patients with normal body temperature, those with fever had higher odds of gametocyte infection (aOR = 1.36, 95% CI: 1.15–1.61, P < 0.001) (Table 3). Additionally, there was a variation in P. vivax gametocyte carriage across different years (aOR = 2.01, 95% CI: 1.90–2.12, P < 0.001) (Table 3). Table 3. Results from logistic regression for risk factors for gametocyte carriage in Plasmodium vivax and P. falciparum infectionsVariablesP. vivax**P. falciparumaOR (95% CI)P-valueaOR (95% CI)P-valueSeason Dry11 Rainy (May to October)0.74 (0.62–0.89)0.00130.39 (0.14–1.20)0.0881Sex Male11 Female1.00 (0.86–1.17)0.99660.69 (0.25–1.70)0.4315Age (years) < 511 5–140.96 (0.72–1.27)0.77880.50 (0.10–2.79)0.4051 15–240.75 (0.56–0.99)0.04660.22 (0.05–1.19)0.0678 25 + 0.87 (0.65–1.17)0.36060.16 (0.03–0.85)0.0257Ethnicity Non-Kachin11 Kachin0.95 (0.62–1.43)0.803316.94 (0.96–674.02)0.0885Malaria history^a^ No11 Yes17.54 (11.20–28.90) < 0.00166.76 (27.17–180.31) < 0.001Travel history No11 Yes1.89 (0.62–7.24)0.29804.16 (0.33–25.45)0.1786Fever No11 Yes1.36 (1.15–1.61) < 0.0010.62 (0.24–1.61)0.3160Parasite density (Log, parasites/μl)2.61 (2.31–2.95) < 0.0010.85 (0.73–0.99)0.0358 Year2.01 (1.90–2.12) < 0.001**1.16 (0.94–1.44)0.1601aOR adjusted odds ratios, CI confidence interval^a^ Malaria reported in the past 12 monthsBold indicates a P value less than 0.05
In P. falciparum infections, individuals over 25 years old were less likely to carry gametocytes than children under 5 years old (aOR = 0.16, 95% CI: 0.03–0.85, P = 0.0257) (Table 3). Similar to P. vivax, P. falciparum malaria patients with a history of malaria had significantly higher odds of gametocyte positivity compared to those without previous malaria history (aOR = 66.76, 95% CI: 27.17–180.31, P < 0.001) (Table 3). Interestingly, parasite density was identified as a protective factor against gametocyte carriage. Patients with high falciparum parasite density had lower odds of gametocyte positivity compared to those with low parasite density (aOR = 0.85, 95% CI: 0.73–0.99, P = 0.0358) (Table 3).
Risk factors for Plasmodium gametocyte density among gametocyte carriers
The results of the multivariable analysis for gametocyte density identified several statistically significant factors influencing gametocyte density in P. vivax infections. Similar to the risk factors for gametocyte carriage, seasonal factors, age, and parasite density had a significant impact, with a lower gametocyte density during the rainy season compared to the dry season (aOR = 0.70, 95% CI: 0.66–0.75, P < 0.001) (Table 4). Furthermore, compared to children under 5 years old, individuals over 25 years old had lower gametocyte density (aOR = 0.84, 95% CI: 0.76–0.93, P = 0.0012) (Table 4). Patients with high parasite density exhibited higher gametocyte density compared to those with low parasite density (aOR = 1.09, 95% CI: 1.06–1.11, P < 0.001) (Table 4). Similarly, travel history also had a statistically significant effect; individuals with a travel history had more than twice the gametocyte density (aOR = 2.07, 95% CI: 1.48–2.91, P < 0.001) (Table 4). However, having a previous episode of malaria within the last 12 months was associated with lower gametocyte density compared to individuals without a history of malaria (aOR = 0.53, 95% CI: 0.49–0.57, P < 0.001) (Table 4). Body temperature was also a protective factor for gametocyte density in P. vivax. Patients with fever had lower gametocyte density compared to those with normal body temperature (aOR = 0.86, 95% CI: 0.81–0.92, P < 0.001) (Table 4). There was also a difference in gametocyte density by sex. Compared to males, females had lower gametocyte density (aOR = 0.88, 95% CI: 0.83–0.93, P < 0.001) (Table 4). Furthermore, gametocyte density decreased annually (aOR = 0.78, 95% CI: 0.77–0.79, P < 0.001) (Table 4). Table 4. Results from multivariable Gaussian regression for risk factors for density in Plasmodium vivax and P. falciparum infectionsVariablesP. vivax**P. falciparumaOR (95% CI)P-valueaOR (95% CI)P-valueSeason Dry11 Rainy (May to October)0.70 (0.66–0.75) < 0.0010.14 (0.03–0.60)0.0115Sex Male11 Female0.88 (0.83–0.93) < 0.0011.73 (0.40–7.54)0.4695Ethnicity Non-Kachin11 Kachin1.05 (0.90–1.23)0.54350.18 (0.00–38.69)0.5372Age (years) < 511 5–140.93 (0.84–1.02)0.13142.05 (0.29–14.2)0.4732 15–240.95 (0.86–1.05)0.30370.94 (0.15–5.88)0.9494 25 + 0.84 (0.76–0.93)0.00120.76 (0.10–5.76)0.7907Malaria history^a^ No11 Yes0.53 (0.49–0.57) < 0.0010.84 (0.25–2.83)0.7860Travel history No11 Yes2.07 (1.48–2.91) < 0.0010.53 (0.01–25.67)0.7493Fever No11 Yes0.86 (0.81–0.92) < 0.0011.28 (0.38–4.28)0.6948Parasite density (Log, parasites/μl)1.09 (1.06–1.11) < 0.0011.01 (0.81–1.25)0.9349 Year0.78 (0.77–0.79) < 0.001**1.08 (0.79–1.48)0.6121aOR adjusted odds ratios, CI confidence interval^a^Malaria reported in the past 12 monthsBold indicates a P value less than 0.05
In P. falciparum gametocyte carriers, gametocyte density was lower in the rainy season compared to the dry season (aOR = 0.14, 95% CI: 0.03–0.60, P = 0.0115) (Table 4). Unlike P. vivax, no significant differences were observed in body temperature (aOR = 1.28, 95% CI: 0.38–4.28, P = 0.6948), sex (aOR = 1.73, 95% CI: 0.40–7.54, P = 0.4695), malaria history (aOR = 0.84, 95% CI: 0.25–2.83, P = 0.7860), travel history (aOR = 0.53, 95% CI: 0.01–25,67 P = 0.7493), and asexual parasite density (aOR = 1.01, 95% CI: 0.81–1.25, P = 0.9349) (Table 4).
Effect of genotyping on Plasmodium gametocyte carriage and gametocytemia
Of the 339 randomly selected P. vivax samples, 331 were successfully genotyped. Three main Pvmsp-3α genotypes were identified based on their PCR product size: type A (1900–2000 bp), type B (1400–1500 bp), and type C (1100–1300 bp). RFLP analysis revealed a total of 19 cases of mixed infection. The PCR products of Pvmsp-3β showed two main types of size polymorphism based on PCR product size: type A (1700–2200 bp) and type B (1400–1500 bp). RFLP fragments digested by PstI showed a total of 59 cases of mixed infection. Combining Pvmsp-3α and Pvmsp-3β genes, PCR–RFLP analysis detected a total of 70 mixed infection cases. In P. vivax populations, the proportion of polyclonal infection in samples with gametocyte carriers was significantly higher than in samples without gametocytes (P = 0.0388, Fig. 3A). The mean P. vivax gametocyte density in polyclonal infections was significantly higher than in monoclonal infections (P < 0.0001, Fig. 3B).Fig. 3. Relationship between gametocyte carriage or gametocytemia (gametocytes/µl of blood) and MOI value in Plasmodium vivax (A and B) and P. falciparum (C and D). Gametocytemia on the y-axis is on a log10 scale. MOI Multiplicity of infection
Although 510 P. falciparum malaria samples were examined by microscopy, only 166 DBSs were collected from these clinical cases. Among these, 24 samples were diagnosed as gametocyte-positive by light microscopy. All DBS samples of P. falciparum were successfully genotyped, with the MAD20 allele gene family being the most common Pfmsp1 gene variation in infected patients, while the 3D7 allele gene family in Pfmsp2 was more prevalent than the FC27 family. The MOI range was 1–5, and the proportion of infections with MOI ≥ 2 tended to be higher in samples with gametocyte carriers than in samples without gametocytes (P > 0.05, Fig. 3C). P. falciparum gametocyte density increased with the MOI value (P > 0.05, Fig. 3D).
Discussion
Efforts to control malaria along international borders are crucial for achieving malaria elimination. Despite strengthened malaria control measures, such as the widespread use of long-lasting insecticidal nets (LLINs) and indoor residual spraying (IRS), a significant malaria outbreak occurred between 2015 and 2017, and a rebound occurred in 2020 along the China-Myanmar border. Notably, the outbreak of COVID-19 in 2019 likely disrupted effective malaria control measures by diverting resources and attention from malaria-related activities, potentially contributing to an increase in malaria cases [29, 30]. Gametocytes play a critical role in the life cycle of malaria parasites and serve as a key link in the transmission of malaria from humans to mosquitoes. A deeper understanding of gametocyte carriage dynamics and their transmission implications can inform more effective, targeted interventions to reduce the infectious reservoir and achieve malaria elimination.
During the 10-year study (2011–2020), gametocyte carriage occurred more frequently during the rainy season, following the same general pattern as symptomatic malaria infections (although the proportion of gametocyte-positive individuals was highest during the dry season, when the overall number of cases was lower). This finding is consistent with prior reports indicating that the prevalence of gametocytes in other transmission settings generally correlates with the prevalence of asexual parasites [31, 32]. Consistent with earlier studies, gametocytes were observed more frequently in P. vivax infections than in P. falciparum infections [33]. This may be attributed to the distinct biological characteristics of these two parasite species. P. vivax gametocytes mature faster and can be detected in the bloodstream within 3 to 5 days after the initial microscopic identification of asexually replicating parasites. In contrast, P. falciparum gametocytes exhibit markedly different morphological features, require 8 to 10 days to mature, and accumulate and sequester during their development in the bone marrow parenchyma before emerging into the blood circulation [34, 35]. Nevertheless, gametocyte carriage and density can be influenced by various other factors, including individual or host characteristics, geographical epidemiological transmission patterns, human host factors, parasite biology, and intervention strategies [36–41].
Environmental factors, including climate, temperature, humidity, and seasonality, may influence the prevalence of gametocytes. In certain tropical regions with endemic malaria, transmission predominantly occurs during the rainy season, which typically lasts several months, whereas malaria transmission ceases during the prolonged dry season [42]. Conversely, some studies in areas with high malaria transmission intensity have indicated that seasonality has minimal or no impact on the proportion of gametocyte-positive infections [43]. In Thailand, research has demonstrated that the prevalence of gametocyte in both P. vivax and P. falciparum infections is significantly higher during the dry season compared to the rainy season, showing a negative correlation with rainfall levels [44, 45]. Consistent with these findings, this study observed a significant increase in the proportion of gametocyte positivity for both P. vivax and P. falciparum during the dry season relative to the rainy season. Although the number of mosquitoes is often lower in the dry season compared to the rainy season, mosquito activity becomes more concentrated and efficient, thereby enhancing the likelihood of malaria transmission [46, 47]. Relatedly, a second peak of malaria cases also occurs in some parts of the GMS, although the seasonal peak during the rainy season is most consistent in the malarious regions of this area [48].
Our results indicate that older age in children and adults serves as a protective factor against gametocyte carriage in both P. vivax and P. falciparum, correlating with lower gametocytemia levels in P. vivax. Similar patterns of gametocyte carriage have been reported in other studies, where gametocyte prevalence was higher among the younger age groups [36, 43, 49, 50]. With increasing age, factors such as the maturation of the immune system, the formation of immune memory, enhanced antibody levels, and immune tolerance developed through long-term infections may gradually improve the host’s ability to control malaria parasites [36]. The high prevalence of gametocyte carriage among the younger age group (< 5 years) highlights their potential role in sustaining malaria transmission in the region. Children have been identified as critical contributors to the malaria infectious reservoir in various settings [36]. However, some studies report that adults can exhibit significantly higher gametocytemia compared to children under the age of 12 years [36, 51]. This may also be attributed to their behavior, body size and relative exposure to mosquito bites [22].
Patients with a history of malaria infection constitute a substantial risk factor for Plasmodium gametocyte carriage in this study. This observation is consistent with a 19-year longitudinal analysis of a malaria cohort, which revealed that previous episodes of clinical malaria contribute to the development of gametocyte carriage [52]. Previous malaria infections significantly increase the likelihood of gametocyte carriage due to the complex interplay of biological, immunological, and epidemiological factors. In P. vivax, many episodes arise from hypnozoite stages in the liver that remain dormant following previous infections via an unknown mechanism. The presence of hypnozoites leads to relapses, causing recurrent waves of asexual parasites and gametocytes [53]. Even in individuals with partial immunity, these relapses contribute to sustained gametocyte carriage. In P. falciparum, chronic infections and immune pressure can lead to low-level, ongoing gametocyte production [54]. An epidemiological study conducted along the western border of Thailand indicates that recrudescent infections increase the risk of persistent gametocyte carriage in P. falciparum by 2.3 times [55]. Furthermore, our study indicates that a history of malaria infection may paradoxically lead to a reduction in gametocyte density in P. vivax. Antibodies targeting malaria parasites are produced in individuals with a history of malaria infection, including those that respond to gametocyte-infected erythrocyte surface antigens during the human blood cycle, potentially reducing gametocytemia in these patients [56–58].
Moreover, we found that fever exerts a positive influence on gametocyte prevalence but is negatively associated with gametocytemia in P. vivax infections*.* Consistently, McKenzie et al. reported that patients with gametocytes had higher fever and parasitemia than those without gametocytes in P. vivax infections, suggesting that the presence of gametocytes may exacerbate febrile responses [33, 59]. In contrast, a study in Gambian indicated that children with fever (> 37.4 ℃) or high parasite densities (> 100,000 parasites/ml) were less likely to carry gametocytes [60]. This discrepancy may reflect regional differences in malaria transmission dynamics or could be related to variations in patient immunity, such as inflammatory responses. Additionally, travel history was associated with increased gametocyte density in P. vivax infections in this study. Some studies have reported that individuals with a history of travel to malaria-endemic areas have a higher risk of malaria infection, particularly in regions with relatively high transmission intensity [61], although no direct correlation with gametocyte density has been established.
Regarding parasitic factors, it is crucial to note that P. falciparum and P. vivax exhibit different patterns of gametocyte development and transmission. In P. falciparum, gametocytes typically appear 10–14 days post-infection, whereas in P. vivax, gametocytes emerge concurrently with asexual stages [62]. Furthermore, parasite density and the stage of infection can significantly influence gametocyte carriage.For example, asexual parasitemia is associated with gametocyte carriage in both P. falciparum and P. vivax [63–66], and higher asexual parasitemia levels are linked to an increased risk of gametocytemia [67]. Our findings indicate that asexual parasite density serves as a risk factor for both gametocyte carriage and density in P. vivax, but acts as a protective factor against gametocyte production in P. falciparum.
Previous studies have demonstrated that mixed-genotype infections lead to increased transmission compared to single-clone infections in rodent Plasmodium infections [68]. Additionally, it has been reported that MOI can lead to prolonged or persistent gametocyte production [19, 69]. It has been proposed that polyclonal P. falciparum infections last longer than monoclonal infections, and can increase the likelihood of gametocyte production [70], although some studies have found no correlation between genetic diversity and gametocyte prevalence [71, 72]. In this study, we found that P. falciparum-infected patients with gametocytes exhibited relatively high polyclonality, suggesting that intraspecific competition within Plasmodium infections may facilitate gametocyte development in this population. We also found that polyclonal infections are significantly correlated with gametocyte carriage and density in P. vivax. These polyclonal infections may persist in individuals, enhancing gametocyte carriage and providing increased opportunities for certain parasite variants to evade the host immune system’s antiparasitic response while dedicating resources to gametocyte formation [70], thereby promoting both the development and persistence of gametocytes.
Despite the significant findings of this study, several limitations should be acknowledged. First, while molecular tools (e.g., qRT-PCR targeting pvs25/pfs25) offer substantially greater sensitivity for gametocyte detection, logistical constraints in this resource-limited setting necessitated reliance on LM [73, 74]. The evidence demonstrates that gametocyte carriage is nearly universal in both P. vivax and P. falciparum infections when assessed by molecular methods, with frequently submicroscopic yet remaining infectious to mosquitoes [75]. Therefore, our LM-derived estimates of gametocyte prevalence and density likely represent a lower bound, with true carriage rates potentially exceeding those reported. Importantly, LM remains clinically relevant as it effectively identifies the most infectious individuals—those with gametocytemia levels strongly predictive of mosquito infectivity [12]. Second, although our genotyping approach reliable identifies predominant clones, it exhibits reduced sensitivity in detecting polyclonal infection and minority clones compared to recent amplicon deep sequencing methods [76, 77]. Additionally, while our PCR–RFLP based method offers estimates of clonal diversity, employing a maximum likelihood approach with COIL using the initially targeted SNPs would allow for more precise estimation of the probable number of clones in each P. vivax infection [78, 79]. The sample size of P. falciparum was relatively small, which may limit the generalizability of the results. However, our findings here do reflect the ongoing reduction of P. falciparum transmission along the China-Myanmar border. Third, accurate treatment information was not collected for many patients with a history of malaria infection, as specific information on treatment regimens and durations was unavailable. Previous studies have shown that treatment with conventional antimalarial drugs such as chloroquine, amodiaquine, sulfadoxine-pyrimethamine, or their combinations is associated with persistence of P. falciparum gametocytemia for weeks to months [80]. Additionally, observations from malariotherapy patients suggest that co-infection with P. malariae may increase P. falciparum gametocyte production [81]. A recent short communication indicates that co-infection can either enhance or decrease gametocyte carriage, depending on the specific interactions between the species involved [82]. Due to the limited number of microscopic gametocyte-positive cases for P. falciparum, the effect of co-infection on gametocyte production was not analyzed. Finally, this study focused on symptomatic patients and would undoubtedly have missed other infections in the broader communities from which these patients originate (including symptomatic patients who do not seek treatment and individuals who are asymptomatic and therefore have no suspected reason to seek care). While clinical cases remain a priority for treatment and surveillance, their contribution to overall transmission may be eclipsed by asymptomatic reservoirs in elimination settings, as asymptomatic infections frequently harbor persistent infectious gametocytes [83, 84]. Our analysis should therefore not necessarily be considered to be representative of the entire parasite reservoir in this setting or elsewhere. Nevertheless, this study provides valuable insights into potential factors affecting gametocyte carriage and density, contributing to a deeper understanding of the risk factors related to malaria gametocyte along the China-Myanmar border.
Conclusions
This study highlights the complex interplay of biological, environmental, and epidemiological factors that influence malaria gametocyte carriage and density along the China-Myanmar border. Younger age, a history of malaria infection, travel, and polyclonal infections emerged as key contributors to increased gametocyte prevalence, particularly in P. vivax infections. Gametocyte carriage and density were higher in the dry season than in the rainy season for both species (P. falciparum and P. vivax). These findings underscore the importance of targeted strategies that consider host, parasite, and environmental dynamics to effectively reduce the infectious reservoir and advance malaria elimination efforts in border regions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1WHO. World malaria report 2024. World Health Organization. https://www.who.int/teams/global-malaria-programme/reports/world-malaria-report-2024. Accessed 11 Dec 2024.
- 2WHO. More malaria cases and deaths in 2020 linked to COVID-19 disruptions. World Health Organization. 2021. https://www.who.int/news/item/06-12-2021-more-malaria-cases-and-deaths-in-2020-linked-to-covid-19-disruptions. Accessed 6 Dec 2021.