New record of Anopheles superpictus (Diptera: Culicidae) in central Italy (Tuscany): resting adults and evidence of natural breeding sites
Ilaria Bernardini, Francesco Severini, Michela Menegon, Claudia Mangiapelo, Riccardo Bianchi, Gioia Bongiorno, Marco Di Luca, Daniela Boccolini

TL;DR
Anopheles superpictus, a malaria-transmitting mosquito, was newly recorded in central Italy, with evidence of natural breeding sites, highlighting its potential role in malaria resurgence.
Contribution
First documentation of Anopheles superpictus in central Italy and confirmation of its natural breeding sites in the region.
Findings
Anopheles superpictus was the most abundant mosquito species collected in the Maremma Plain, comprising 46% of specimens.
Larvae of Anopheles superpictus were found in seven of twelve surveyed sites along the Albegna riverbed.
The species was confirmed to occur stably in the area through repeated surveys in 2023 and 2024.
Abstract
Anopheles superpictus (subgenus Cellia) plays an efficient role in malaria transmission in countries of the Mediterranean basin, Eastern Europe, and the Caucasus region, where it has been involved in the transmission of both Plasmodium falciparum and P. vivax. In Italy, this species was historically considered a secondary malaria vector, primarily recorded in the South including Sicily, along small rivers. It was less frequently observed in central Italy, only rarely reported in the North. Between 2022 and 2024, as part of routine investigations on residual anophelism, An. superpictus specimens were collected for the first time in the southern Maremma Plain (Magliano in Toscana, Grosseto, Tuscany). Adult mosquitoes were collected in animal shelters and tool premises of two farms using traps and manual aspiration methods. Larval sampling was performed by exploring potential breeding…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
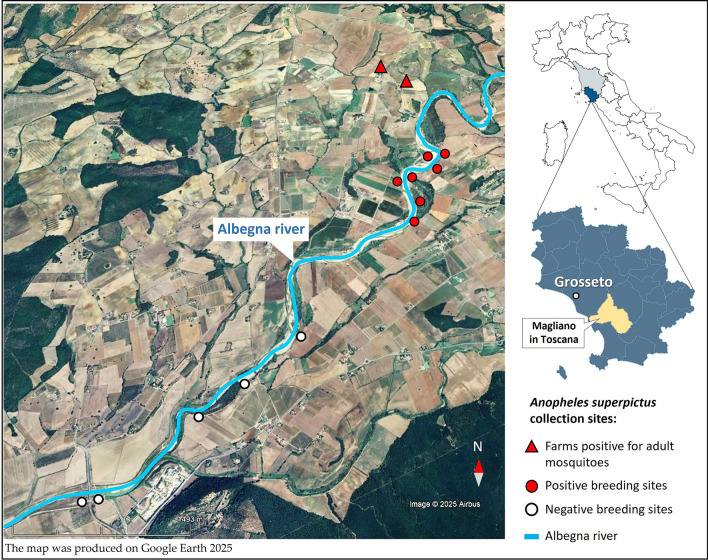 Figure 1
Figure 1 Figure 2
Figure 2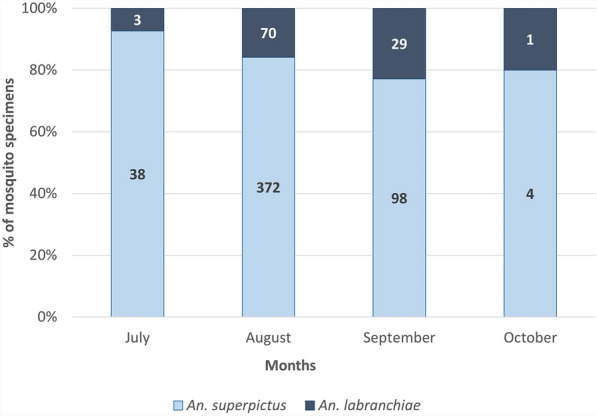 Figure 3
Figure 3 Figure 4
Figure 4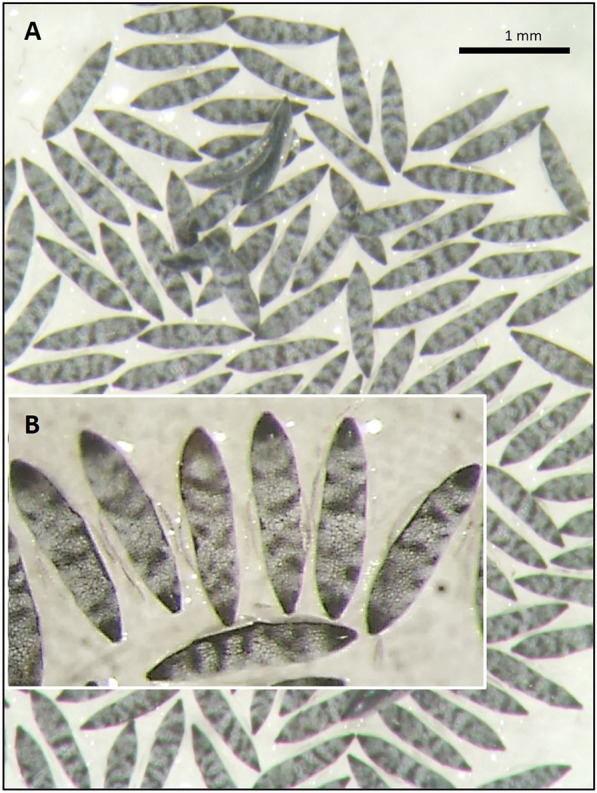 Figure 5
Figure 5 Figure 6
Figure 6- —"Definition of the integrated vector management strategy and its application in the Municipal Arboviral Plans within the Rete Città Sane"
- —EU funding within the NextGeneration EU-MUR PNRR Extended Partnership initiative on Emerging Infectious Diseases
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMosquito-borne diseases and control · Malaria Research and Control · Insect Resistance and Genetics
Background
Malaria is a disease that still poses globally important challenges. In endemic countries, it continues to represent one of the more threatening public health concerns [1]. In non-endemic settings, it is the most frequently imported parasitic disease, and the persistence of putative vectors [2] underlines the need to implement and strengthen surveillance systems, particularly across European countries bordering the Mediterranean basin [3]. In Italy, Plasmodium falciparum transmission was virtually interrupted by 1948, just one year after the launch of the National Eradication Campaign. Nonetheless, sporadic P. vivax cases continued to be reported until 1962, and the country was officially declared malaria free in 1970 [4]. Former malaria vectors, such as Anopheles labranchiae Falleroni, 1926, An. sacharovi Favr, 1903, both members of the An. maculipennis complex (Anopheles) Meigen 1818, and An. superpictus (Cellia) Grassi 1899 are still present in scattered foci across the country. Anopheles labranchiae has been recorded along central-southern coastal areas, including Sicily and Sardinia [5–7]. Anopheles sacharovi historically restricted to parts of the Adriatic and Tyrrhenian coastlines and Sardinia, was presumed extinct since the late 1960s but was recently rediscovered in the Apulia region (Lecce province), where it had never been recorded during the malaria-endemic period [8]. The third taxon, i.e. An. superpictus, was historically considered a secondary malaria vector in Italy [9–11]. However, across its broad Palearctic distribution range, this species has instead played an efficient role as a malaria vector [12–14]. In the southeastern Mediterranean basin, including North Africa and West Asia, as well as in Eastern Europe and the Caucasian area of the Western Palearctic Region [15–17], it has been involved in the transmission of both agents of human malaria, P. falciparum and P. vivax [18–22]. The competence of An. superpictus for P. vivax transmission has also been supported by an experimental infection study [23]. In contrast, its susceptibility to P. falciparum has never been directly tested. However, several authors have hypothesized that this species, belonging to the subgenus Cellia, which includes some of the most efficient African malaria vectors, could be permissive to Afrotropical strains of P. falciparum [9–12], unlike the refractoriness observed in vector species of the An. maculipennis complex [24, 25].
In Italy, An. superpictus exhibits a phenology typical of late summer mosquitoes, with adult populations usually peaking between August and September. Females are partially endophagic and exophilic, displaying opportunistic feeding behaviour preferentially shifted to large mammals when available. During the resting phase, they prefer warm indoor shelters and outbuildings [18]. As for larval habitats, the species generally breeds in residual pools formed in gravelly riverbeds when water flow decreases significantly in late summer and autumn. Larvae are typically found in small, slow-flowing pools on rocky riverbeds with scattered vegetation. The limited availability of such specific breeding sites, along with climatic variability and anthropogenic environmental changes, has significantly influenced the distribution and abundance of the species [9, 12, 18, 26, 27].
In the past, An. superpictus was collected in southern Italy including Sicily, predominantly along small river courses. It was less frequently found in central regions, only rarely recorded in the North [4, 16]. Currently, its range of distribution appears to be significantly restricted. In recent years, only sporadic records of few adult specimens have been documented in southern regions, such as Calabria, Basilicata and Apulia [11, 28, 29]. However, despite its apparent decline in abundance and range, An. superpictus should still be taken into consideration as a potential malaria vector, contributing to increase the receptivity of southern Italian localities [11, 29].
Considering the ongoing climatic and environmental changes, along with increased human mobility, entomological surveillance remains crucial for assessing the potential risk of malaria reintroduction in vulnerable areas. Data from field surveys contribute to strengthening the basic knowledge required to improve national prevention strategies. Within this framework, and as part of routine efforts to update residual anophelism data, entomological investigations were carried out between 2022 and 2024 in central Italy, specifically in the southern Maremma Plain (Grosseto province, Tuscany), a historically endemic region for malaria before its eradication. This study aims to describe the survey activities carried out and presents the findings obtained.
Methods
Study area
The study area is located in the southernmost part of the Maremma Plain countryside, specifically in the municipality of Magliano in Toscana (Grosseto province), approximately 20 kms from the Tyrrhenian Sea. The territory consists of modest hilly reliefs, none of which exceed 300–350 m above sea level (a.s.l.), along with some valleys traversed by the main river Albegna and its tributary streams. The Albegna riverbed is wide, gravelly and with riparian vegetation along the riverbank. It maintains a good level of cleanliness, flowing through a naturalistically significant environment included in the Maremma Park (SIR-SIC-ZPS code: 121 “Medio Corso del Fiume Albegna”—code Natura 2000 IT51A0021 and SIR-sir code B22 “Torrente Trasubbie” cod. IT5190103). Human impact on the area is limited, with contiguous arable fields and pastures. Typical settlement patterns consist of scattered houses and/or farms. In 2022, the residential population registered for the Municipality was 13.1 inhabitants per square kilometre. In this area, the climate is typically Mediterranean. During 2022, July and August were the hottest months. Maximum temperatures ranged from 32.5 °C in July to 21.6 °C in October; the average relative humidity was around 66%. In total 13 rainy days were recorded, with cumulative precipitation reaching 253.2 mm, of which 222.6 mm fallen in September (https://www.sir.toscana.it/consistenza-rete).
Sampling sites
Adult mosquito collections were carried out in two farms (42°35′21.9″ N; 11°22′09.8″ E), where domestic animals, particularly backyard poultry and stabled cattle are reared. Both sites are located below 300 m a.s.l. and lie near the course of the Albegna river (Fig. 1). For larva collections twelve potential breeding sites, situated within a radius of 1–5 kms from the farms, and along a slow-flowing section of the river characterized by shallow gravelly pools and aquatic vegetation, were investigated (Figs. 1 and 2).Fig. 1. Map of the study area (Magliano in Toscana municipality, Grosseto province, Tuscany). Triangles represent the two farms where Anopheles superpictus adult samples were collected. Red dots indicate the positive larval breeding sites along the Albegna river, while white dots represent the negative onesFig. 2Larval breeding sites of Anopheles superpictus along the Albegna riverbed (Magliano in Toscana, Grosseto, Tuscany). A general view of the site. B close-up of a residual pool surrounded by low vegetation, showing in the right-hand box a detail of larval aggregation
Mosquito collection
In 2022, adult mosquitoes were collected from July to October by using BG-PRO (Biogents^®^) and CDC light traps, placed at five different sites inside and outside the animal stables, operating from sunset to dawn. In addition, in September 2022, supplementary active sampling was conducted using electric aspirators to collect resting adults inside animal shelters and nearby tool premises. Larval sampling was also carried out along the Albegna riverside using standard 500 ml dippers, with 10–20 dips per site [30]. In September 2023 and 2024, only two snapshot surveys were conducted mid-monthly, using the same adult collection methods.
All adult specimens were stored at − 20 °C before their processing. Larvae were reared to the fourth instar for morphological identification, and then preserved in 70% ethanol.
Specimen processing
Both larvae and adults were identified using dichotomous morphological keys of the Italian mosquito species [31–33]. Observations of morphological characters of the larvae and the adult specimens were performed in Phosphate-Buffered Saline solution (PBS) and at dry conditions respectively, under a stereomicroscope (Leica Stereozoom S9i). For species identification within the An. maculipennis complex specimens, gravid females were induced to lay eggs and the exochorion banding pattern was observed [34].
Furthermore, a subsample of An. superpictus were molecularly analysed for species confirmation by sequencing both the cytochrome c oxidase subunit 1 (cox1) gene and the ribosomal DNA internal transcribed spacer region 2 (ITS2). An 889-base pair fragment of the cox1 gene was amplified by combining the forward primer LCO1490 [35] and the reverse primer (5′AAAAATTTTAATTCCAGTTGGAACAGC 3′) described in Kumar et al. [36]. The ITS2 region was amplified using the 5.8S primer forward (5′-TGT GAA CTG CAG GAC ACA TG-3′) [37] and Hyr reverse primer (5′-GGG GTW GTC ACA CAT AAC TTG AGG-3) [38].
Anopheles maculipennis s.l. specimens were identified only by ITS2 analyses, according to Marinucci et al. [39] protocol.
All PCR products were sequenced at Eurofins Genomics (Ebersberg, Germany) using the aforementioned primers. Resulting sequences were assembled using DS Gene v1.5 software (Accelrys Inc. 2003) and analysed using NCBI's Basic Local Alignment Search (BLAST) [40].
Show data analysis
For the 2022 survey data, differences in mosquito species were assessed using the Shannon–Wiener diversity index (H), calculated as: H = − ∑[(pi) × ln(pi)], where:
∑: sum
pi: proportion of individuals of i-th species in the whole community
ln: natural logarithm
pi = n/N (n*:* individual of a given species; N: total number of individuals in the community)
An H value of 0 indicates that only one species is present in the habitat; increasing H values correspond to greater species diversity.
The presence and abundance of anopheline species were compared using the non-parametric Mann–Whitney test (U-test). Differences were considered statistically significant at P < 0.05.
All statistical analyses were performed using R^®^ software (version 3.5.0).
Results
Longitudinal surveys conducted from July to October 2022 resulted in the collection by traps of 1,106 mosquitoes, peaking abundance in August (n = 646) and September (n = 394). Specimens belonged to four genera: Aedes, Anopheles, Culex and Culiseta and nine species were identified through both morphological and molecular analyses (Table 1). The Shannon–Wiener diversity index (H) was calculated as 1.38, indicating a limited level of mosquito species diversity in the study area. Table 1. Mosquito species collected using BG-PRO and CDC-light traps, from July to October 2022, in Magliano in Toscana municipality (Grosseto , Tuscany)Month of collectionsMosquito speciesTotal per monthAedes albopictusAedes caspiusAedes detritusAnopheles labranchiaeAnopheles superpictusCuliseta annulataCuliseta longiareolataCulex pipiens s.lCulex theileriJuly010338006048August10181703721121602646September3832029982181698394October01114056018Total (percentage)48 (4.3)52 (4.7)2 (0.2)103 (9.3)512 (46.3)3 (0.3)35 (3.2)341 (30.8)10 (0.9)1106
Overall, among the Culicinae subfamily, Culex pipiens s.l. is the most abundant, accounting for 30.8% (260 females, 81 males,), followed by Aedes caspius (47 females, five males), Ae. albopictus (26 females, 22 males), Culiseta longiareolata (13 females, 22 males), Cx. theileri (seven females, three males), Cs. annulata (three females), Ae. detritus (two females).
Species belonging to the Anophelinae subfamily accounted for 56% of all specimens collected. Anopheles superpictus was the most abundant species, representing 46.3% of the total sample with 487 females, and 25 males, reaching its highest number in August (n = 372) and sharply declining in October (n = 4) (Table 1). Anopheles labranchiae was also recorded, with 9.3% of the total sample, including 97 females, and six males. The monthly percentage of the two anopheline species is shown in Fig. 3. The Mann–Whitney test (U test) compares the abundances of An. superpictus and An. labranchiae yielded U = 101 (P = 0.0003), indicating a statistically significant predominance of the former species.Fig. 3. Monthly percentage distribution of Anopheles superpictus and An. labranchiae collected using BG-PRO and CDC light traps from July to October 2022 in two farms located in the municipality of Magliano in Toscana (Grosseto, Tuscany). Histogram columns represent the percentage distribution, with total specimen counts shown above each bar
The sex ratio of all collected mosquitoes was mostly biased toward females, particularly among Anopheles species, likely due to the reduced dispersal capacity of males from breeding sites.
Focusing on manual anopheline captures in September, a total of 30 engorged and gravid females were collected in a tool premise nearby a livestock shed, including 20 An. superpictus and 10 An. labranchiae specimens; consistent with trap collection results, An. superpictus was predominant, accounting for 67% of the specimens, compared to 33% for An. labranchiae.
Among the twelve breeding sites investigated in the Albegna riverbed area, selected based on accessibility and proximity to farms, seven (58%) tested positive for An. superpictus larvae. Overall, 50 specimens were collected, with mean densities ranging from 0.1 to 0.2 larvae per dip (Fig. 2). All adults and larvae of the An. superpictus were morphologically identified based on species-specific characters, as illustrated in Fig. 4.Fig. 4. Morphological characteristics of Anopheles superpictus collected in the municipality of Magliano in Toscana (Grosseto, Tuscany). A IV instar larva. A.1: inner clypeal setae; A.2: long mesothoracic pleural setae, one simple, the other one with minute lateral branches, diagnostic for the species; A.3: long metathoracic pleural setae, both with lateral branches. B Adult female. B.1: Head with white scales at the apex of the maxillary palpi; B.2: colour pattern of the costal vein (subgenus Cellia); B.3: colour pattern of the anal vein; B.4: dark and pale spots on the cubital vein
As support of morphological identification, molecular analyses were carried out on 4% of the adult An. superpictus specimens (n = 20 out of 512). All cox1 and ITS2 sequences obtained from the 20 specimens were identical, with no observed nucleotide variation. One representative sequence for each marker was deposited in GenBank. The accession numbers of the newly generated sequences acquired in this study are PQ570580 (ITS2) and PQ569434 (cox1). Moreover, BLAST analysis of cox1 and ITS2 obtained sequences showed a > 99% nucleotide homology with An. superpictus available sequences in GenBank. In particular, the cox1 consensus fragment of 889 nucleotides exhibited 99.10% identity with a sequence from a specimen collected in Greece (MT993498). When considering only the cox1 region (658 nucleotides), that would be obtained by amplification with the most utilized LCO1480-HCO2198 primers [35], identities ranged from 99.09 to 98.94% when compared to several sequences derived from samples collected in Tajikistan (JX255703-JX255708). Additionally, ITS2 sequences exhibited 100% identity with those obtained from An. superpictus from several WHO European and Eastern Mediterranean countries, including Greece (FJ526598), Uzbekistan (AY515163), Afghanistan (EU482199), Iran (KF483835) and Pakistan (DQ487149).
Specimens included in the An. maculipennis s.l. were identified as An. labranchiae through egg pattern observations (Fig. 5). The species was also confirmed by ITS2 sequencing as the only member of the complex detected in the area. The GenBank accession number of An. labranchiae ITS2 sequence is PV798893.Fig. 5. Oviposition of Anopheles labranchiae collected in Magliano in Toscana municipality (Grosseto, Tuscany). A: General view of the egg-banding pattern, showing the egg exochorion with wedge-shaped dark marks on a pale background. B: Magnified view of the eggs, showing short and narrow floats
In September 2023 and 2024, two snapshot surveys were conducted using CDC light traps to confirm the continued presence of An. superpictus in the area. During these surveys, 15 and 32 females of this species were collected, along with three and five An. labranchiae specimens.
Discussion
This study, carried out in the period 2022–2024, documented new field records of An. superpictus in sites located in Magliano in Toscana municipality (Grosseto province), at the southern border of Maremma Plain. Notably, no historical or recent data on anopheline mosquitoes were previously available for this area. Moreover, the last records of the An. superpictus in inland localities in Grosseto province date back nearly a century, during anti-malaria control plans implemented at that time [41–43].
In the 2022 longitudinal surveys (July–October), An. superpictus was recorded at consistently high densities. It emerged as the most abundant mosquito species captured by both traps and manual collections. The application of the Shannon–Wiener diversity index provides a quantitative measure supporting the observed species pattern in the study area (Table 1). However, the low H value, which confirms the dominance of An. superpictus, should be interpreted with caution due to the limited spatial and temporal coverage of the sampling.
Larval sampling further enabled the characterization of its natural breeding sites, an achievement unreported in recent decades. The ecological characteristics of the study area appear particularly suitable for the development of An. superpictus. The gravelly bed of the Albegna river, the absence of dense vertical vegetation, and the presence of shallow water bodies, offering larvae protection from larvivorous fish, correspond closely with the known habitat preferences of the species (Fig. 2).
The snapshot surveys conducted in September 2023 and 2024 confirmed the continued presence of the species, although the number of specimens collected was markedly lower than in the same period of 2022. This reduction may partly reflect the limitations of single-time-point sampling methods. However, meteorological factors likely played a role as well.
An. superpictus populations tend to fluctuate in response to river water levels: years with frequent summer showers or prolonged droughts may hinder the formation of suitable breeding sites, leading to reduced mosquito abundance. Likewise, the species’ seasonal trend is variable. As river levels decline during mid-summer and isolated pools occur, optimal breeding conditions emerge, allowing population growth that typically peaks between September and October [18]. From an epidemiological standpoint, such year-to-year fluctuations in vector density may influence the receptivity of an area. In settings where malaria parasites circulate at low levels, increased densities of An. superpictus may enable the species to spread into nearby areas previously considered malaria-free, potentially resulting in malaria outbreaks [12, 18, 44].
In 2022, An. superpictus reached its peak abundance in August and September, the two wettest months of the summer. Total rainfall during this period amounted to 232.2 mm, in stark contrast to 57.8 mm in 2023 and 70.8 mm in 2024. Nevertheless, the snapshot surveys confirmed that even in years with scarce precipitation, when residual pools are reduced, the species remains present in the study area, albeit at lower densities.
In both surveyed farms, adult specimens of An. superpictus were found in sympatry with An. labranchiae. Together, these two anopheline species accounted for more than half of the mosquito fauna collected in the area. However, An. labranchiae was recorded at significantly lower densities compared to An. superpictus.
Larval habitats for An. labranchiae could not be identified in the study area, as no larvae were found during field sampling. This former malaria vector, frequently recorded at notable densities in several areas of Grosseto province [25, 42, 45, 46], typically exploits a wide variety of larval habitats. They can range from coastal salt marshes to artificial water bodies created by human activities, such as rice paddies and irrigation canals, demonstrating the remarkable ecological plasticity of the species [46, 47]. Although the Albegna riverbed offers ideal breeding conditions for An. superpictus, certain sections of the river may also provide suitable environments for An. labranchiae, even if this could not be confirmed through larval sampling.
To support species identification, molecular analyses were also conducted on An. superpictus specimens collected in Magliano in Toscana. Sequence analysis of the cox1 and ITS2 genes revealed a high degree of similarity with multiple haplotypes available in GenBank, originating from populations across several European and Eastern Mediterranean countries. In some of these countries, the species is recognized as a competent malaria vector [19–22, 44]. Furthermore, from a phylogenetic perspective, several studies have identified An. superpictus as a species complex, including at least three closely related taxa, each potentially playing a different role in malaria transmission [14, 48, 49]. Nevertheless, the taxonomic status within this complex has so far remained unresolved [13, 15].
Conclusions
This study, although limited by surveys conducted on a small local scale and on a seasonal basis, contributes substantially to the current knowledge of the presence and distribution of An. superpictus in Italy, providing new and consistent field records of a species rarely reported in recent decades. It also adds valuable evidence to entomological data at the European level [17].
Moreover, the findings may serve as a starting point for future research. Broader investigations across central Italy, combined with population genetic analysis, will be essential to clarify the current epidemiological relevance of the species.
Investigations in Magliano in Toscana revealed that An. superpictus could reach high densities in the study area and, together with the stable presence of An. labranchiae, these findings highlight the importance of regular, targeted monitoring to assess changes in species abundance and distribution over time. These two former malaria vectors may gradually disperse throughout the territory, depending on the availability of suitable larval habitats, potentially leading to dynamic changes in distribution and an evolving receptivity in vulnerable areas.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1World Health Organization. World malaria report 2024. Geneva: WHO; 2024. https://www.who.int/teams/global-malaria-programme/reports/world-malaria-report-2024. Accessed 26 Jun 2025.
- 2European centre for disease prevention and control. Malaria—annual epidemiological report for 2022. Stockholm: ECDC; 2024. https://www.ecdc.europa.eu/en/publications-data/malaria-annual-epidemiological-report-2022. Accessed 26 Jun 2025.
- 3European Centre for Disease Prevention and Control. Anopheles maculipennis s.l.—current known distribution: October 2023. Stockholm: ECDC; 2023. https://www.ecdc.europa.eu/en/publications-data/anopheles-maculipennis-sl-current-known-distribution-october-2023. Accessed 26 Jun 2025.
- 4World Health Organization. Mosquitoes of the genus Anopheles in countries of the WHO European Region having faced a recent resurgence of malaria: regional research project, 2003–2007. Copenhagen: WHO Regional Office for Europe; 2008. Report No.: E 922010. https://iris.who.int/handle/10665/107914.
- 5European Centre for Disease Prevention and Control. Anopheles superpictus – current known distribution: October 2023. Stockholm: ECDC; 2023. https://www.ecdc.europa.eu/en/publications-data/anopheles-superpictus-current-known-distribution-october-2023.
- 6World Health Organization. Malaria vectors and approaches to their control in malaria affected countries of the WHO European Region: proceedings of a regional meeting on vector biology and control; 2001 May 3–5; Almaty, Kazakhstan. Copenhagen: WHO Regional Office for Europe; 2001. Report No.: EUR/01/5027499. https://iris.who.int/handle/10665/107428.
- 7European Centre for Disease Prevention and Control. Guidelines for the surveillance of native mosquitoes in Europe. Stockholm: ECDC; 2014. 111 p. https://www.ecdc.europa.eu/en/publications-data/guidelines-surveillance-native-mosquitoes-europe.