A simple fixation for detection of endogenous fluorescent reporters in C. elegans that nearly eliminates intestinal autofluorescence
Ohm H. Patel, Alexandra S. Weisman, Craig P. Hunter

TL;DR
This paper introduces a simple, chemical-free method to reduce intestinal autofluorescence in C. elegans, improving the detection of endogenous fluorescent reporters.
Contribution
A novel fixation method that preserves GFP localization while nearly eliminating intestinal autofluorescence in C. elegans.
Findings
The method preserves non-cytoplasmic GFP localization for months.
Heat fixation nearly eliminates intestinal autofluorescence in C. elegans.
Variability in non-membrane-anchored reporters suggests the need for empirical validation.
Abstract
Gut granules are prominent cytoplasmic organelles that auto fluoresce when excited by much of the visible spectrum confounding fluorescence imaging of endogenous fluorescent reporters. We report a simple, chemical-free fixation method for C. elegans that preserves non-cytoplasmic GFP localization for months while nearly eliminating intestinal autofluorescence. To illustrate the utility and limitations of the method, we present representative images of live and heat-fixed worms expressing a variety of membrane and non-membrane localized GFP reporters expressed in diverse tissues, cells, and cellular organelles. We also describe our experience with various experimental parameters. The observed variability of non-membrane-anchored reporters suggests prudent adopters should empirically interrogate the signal fidelity of their specific reporters upon heat fixation.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
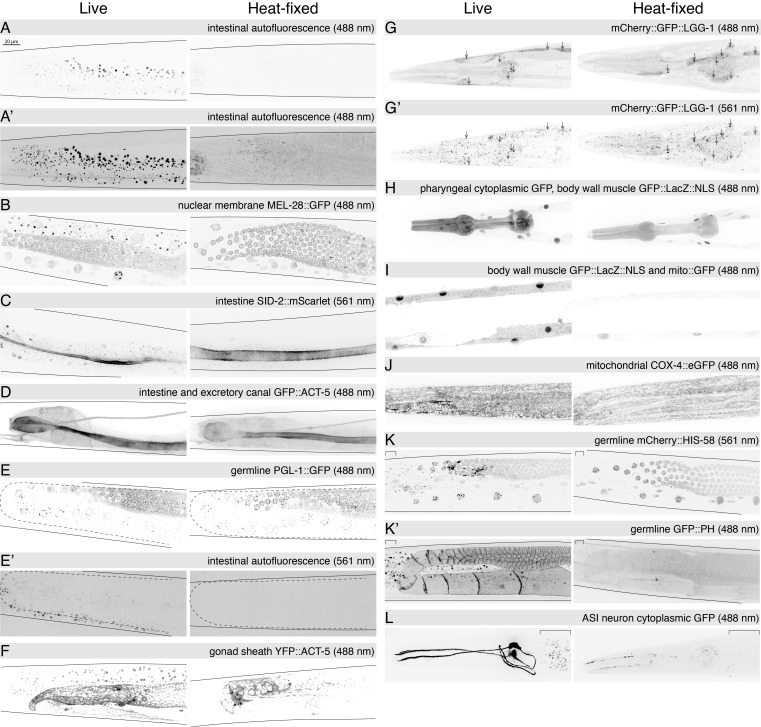 Figure 1
Figure 1|
|
|
|
|
|
|
|
|---|---|---|---|---|---|---|
|
A, A' |
|
wild-type |
|
N/A |
N/A | |
|
B, C |
HC1262 |
|
Hunter Lab |
Gómez-Saldivar et al., 2016 (
|
All;Intestine |
Nuclear membrane, kinetochore and nucleoplasm; Plasma membrane |
|
D |
|
|
CGC |
Szumowski et al., 2015 |
Intestine |
Cytoskeleton |
|
E, E' |
|
|
CGC |
Putnam et al., 2019 |
Germline |
Cytoplasm (P Granules) |
|
F |
|
|
CGC |
Kinchen et al., 2005 |
Somatic gonad sheath |
Cytoskeleton |
|
G, G' |
|
|
CGC |
Chang et al., 2017 |
Multiple somatic |
Autophagosome and Autolysosome |
|
H, I |
|
|
Hunter Lab |
Winston et al., 2002 |
Body wall muscle; Pharyngeal muscle |
Nucleus,mitochondria and cytoplasm; Cytoplasm |
|
J |
|
|
CGC |
Raiders et al., 2018 |
All |
Mitochondria (inner membrane) |
|
K, K' |
|
|
CGC |
Essex et al., 2009 (
|
Germline |
Nucleus; Plasma Membrane |
|
L |
|
|
CGC |
Murakami et al., 2001 |
ASI neuron |
Cytoplasm |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
Topicsbioluminescence and chemiluminescence research
Description
Tissue fixation is normally achieved by a combination of chemical and environmental conditions that cross-link and or denature proteins with or without extraction of lipids and metabolites (Howat and Wilson, 2014). Here, we show that heat treatment alone can be sufficient to preserve the localization and fluorescence of GFP and mCherry reporters while reducing intestinal autofluorescence (AF). Shown in Figure 1 are examples of various fluorescent protein reporters that are either well or poorly preserved by 95°C fixation (Figure 1). The presence of detectable fluorescence suggests that this brief temperature excursion only partially denatures the fluorescent protein (GFP, YFP, mCherry, mScarlet) or that they refold sufficiently well to be fluorescent. These images also demonstrate that intestinal autofluorescence in both green (488 nm) and red (561 nm) fluorescent channels is significantly reduced. This reduction in AF is most obvious in samples without reporters or where the fluorescent reporter signal is less bright ( Figure 1A- C, E', F, K, K', L). In general, heat fixation preserves the brightness and localization of membrane-bound reporters, including plasma membrane ( Figure 1C ), nuclear membrane ( Figure 1B ), and autophagosomal and autolysosomal membranes ( Figure 1G ). Cytoskeletal ACT-5 protein reporters are also well preserved by heat treatment in the intestine, excretory canal, and germline ( ACT-5 , Figure 1D, F). Cytoplasmic, peripheral membrane, nuclear-localized and mitochondrial-localized protein reporters are poorly preserved ( Figure 1 H-L). Surprisingly, the heat fixation did not disrupt detection of the phase separated liquid condensate P granule marker PGL-1 ::GFP ( Figure 1 E). The heat fixation noticeably altered the morphology of the intestinal lumen from a flattened ribbon to a more symmetrical tube shape ( Figure 1 C, D). The excretory canal also appears to collapse ( Figure 1D ).
Although all images presented in Figure 1 represent same-day fixation and imaging to facilitate direct comparison with the live-imaged samples, fixed animals can be stored at 4°C with addition of sodium azide, to discourage microbial growth, for at least six months with only slightly detectable changes in signal strength. Inclusion of levamisole in the fixation condition can cause morphological artifacts (e.g. elongated nuclei). Most strains were tested in multiple independent replicate experiments ( Figure 1, legend). We have not explored post-heat-fix treatments to increase permeability for addition of non-permeable dyes or antibodies. The heat fixation causes the worms to be near-linear, a benefit for studies that involve measuring worm size.
Methods
C. elegans * maintenance as described in (Brenner, 1974). All strains were maintained on normal growth medium plates seeded with OP50 at 20°C.
Heat Fixation
Materials: 95°C water, 1.7 ml polypropylene tube, heat block, S. Medium
Procedure:
Microscopy
For each strain we mounted and imaged live and heat-fixed day 1 adult hermaphrodite worms prepared on the same day from the same culture. We used levamisole (3 mM) to immobilize live worms for microscopy. Fixed and live worms were mounted on 10% grooved agar pads (Rivera Gomez and Schvarzstein, 2018), trimmed to be smaller than the overlying coverslip, and sealed with beeswax. Identical acquisition parameters (laser power, gain, z-slice intervals) were used for live and heat-fixed animals within each experiment. All displayed images (z-stacks) were collected with a Nikon Spinning Disk Confocal (CSU-W1) microscope using a Plan-Apochromat 60x/1.42 Oil objective, laser lines (filters) 488 nm (525/36 nm) for GFP/YFP and 561 nm (605/52 nm) for mCherry/mScarlet at 1-30% laser power, and slice-intervals of 1.0 or 0.2 µm. In some replicate experiments images were acquired using a LD C-Apochromat 40x/1.1 Water objective on a Zeiss LSM980. Collected images were manipulated (orientation, cropping, brightness and contrast) using FIJI/Image J (version 2.16.0/1.54) (Schindelin et al., 2012). Brightness and contrast display settings were identically adjusted for each pair, images are presented with an unsaturated linear display unless otherwise noted in the figure text. Maximum projection images from z-slices corresponding to the thickness of the worm (or one-quarter thickness in panel I) were processed in FIJI/ImageJ and the figure assembled using Adobe Illustrator.
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Brenner S 197451 The genetics of Caenorhabditis elegans.Genetics 7710016-6731719410.1093/genetics/77.1.714366476 PMC 1213120 · doi ↗ · pubmed ↗
- 2Chang Jessica T Kumsta Caroline Hellman Andrew B Adams Linnea M Hansen Malene 201774 Spatiotemporal regulation of autophagy during Caenorhabditis elegans aginge Life 62050-084X 10.7554/elife.18459 PMC 549674028675140 · doi ↗ · pubmed ↗
- 3Essex Anthony Dammermann Alexander Lewellyn Lindsay Oegema Karen Desai Arshad 2009215 Systematic Analysis in Caenorhabditis elegans Reveals that the Spindle Checkpoint Is Composed of Two Largely Independent Branches Molecular Biology of the Cell 2041059-15241252126710.1091/mbc.e 08-10-104719109417 PMC 2642744 · doi ↗ · pubmed ↗
- 4Gómez-Saldivar G Fernandez A Hirano Y Mauro M Lai A Ayuso C Haraguchi T Hiraoka Y Piano F Askjaer P 2016624 Identification of Conserved MEL-28/ELYS Domains with Essential Roles in Nuclear Assembly and Chromosome Segregation.P Lo S Genet 1261553-7390 e 1006131 e 100613110.1371/journal.pgen.100613127341616 PMC 4920428 · doi ↗ · pubmed ↗
- 5Howat William J. Wilson Beverley A. 2014111 Tissue fixation and the effect of molecular fixatives on downstream staining procedures Methods 7011046-2023121910.1016/j.ymeth.2014.01.02224561827 PMC 4240801 · doi ↗ · pubmed ↗
- 6Kinchen Jason M. Cabello Juan Klingele Doris Wong Kelvin Feichtinger Richard Schnabel Heinke Schnabel Ralf Hengartner Michael O. 200531 Two pathways converge at CED-10 to mediate actin rearrangement and corpse removal in C. elegans Nature 43470290028-0836939910.1038/nature 0326315744306 · doi ↗ · pubmed ↗
- 7Murakami Mayumi Koga Makoto Ohshima Yasumi 2001111 DAF-7/TGF-β expression required for the normal larval development in C. elegans is controlled by a presumed guanylyl cyclase DAF-11Mechanisms of Development 10910925-4773273510.1016/s 0925-4773(01)00507-x 11677050 · doi ↗ · pubmed ↗
- 8Nikonorova Inna A. Wang Juan Cope Alexander L. Tilton Peter E. Power Kaiden M. Walsh Jonathon D. Akella Jyothi S. Krauchunas Amber R. Shah Premal Barr Maureen M. 202251 Isolation, profiling, and tracking of extracellular vesicle cargo in Caenorhabditis elegans Current Biology 3290960-982219241936.e 610.1016/j.cub.2022.03.00535334227 PMC 9491618 · doi ↗ · pubmed ↗