Alterations of slitrk5a induce OCD-like “checking behaviors” in zebrafish
Benjamin A. Sempowski, Kathryn R. Woolford, Jenna Bouhussein, Barbara Lom

TL;DR
This study shows that changes in the slitrk5a gene in zebrafish lead to OCD-like repetitive behaviors, suggesting a link between this gene and obsessive-compulsive disorder.
Contribution
The study is the first to demonstrate OCD-like behaviors in zebrafish due to slitrk5a gene alterations, supporting its role in neuropsychiatric conditions.
Findings
slitrk5a gene alterations in zebrafish caused increased repetitive checking behaviors.
The changes did not affect embryonic development or anxiety-like behaviors.
This finding supports a potential role of SLITRK5 in OCD-like behaviors.
Abstract
The six transmembrane SLITRK proteins differentially regulate important aspects of neuronal development and function. Variants in SLITRK5 have been associated with complex neuropsychiatric conditions including obsessive compulsive disorder (OCD) and Slitrk5 knockout mice exhibit overgrooming and anxiety-like behaviors. This study generated slitrk5a zebrafish mutants using CRISPR-Cas9. Alterations in slitrk5a did not affect F 0 gross embryonic development or anxiety-like behaviors, however, a repetitive, checking behavior was significantly increased in the novel approach test (NAT). This observation in zebrafish supports an emerging association of SLITRK5 sequence alterations with OCD-like repetitive behaviors.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
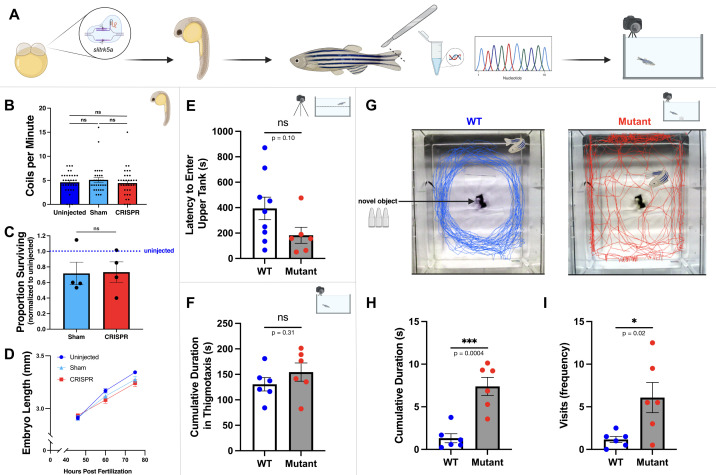 Figure 1
Figure 1|
|
|
|
|
Agarose |
Fisher Scientific |
BP165 |
|
Alt-R™ CRISPR-Cas9 crRNA |
Integrated DNA Technologies |
5’ - GGTTCCTCAAGGAGACCTCC - 3’ |
|
Alt-R™ S.p. Cas9 Nuclease V3 |
Integrated DNA Technologies |
1081058 |
|
Alt-R™ CRISPR-Cas9 tracrRNA |
Integrated DNA Technologies |
1072532 |
|
Aquafeed Z-cubes |
Clear H2O |
60-01-0250 |
|
Brine shrimp eggs |
Bio-Marine |
Artemia cysts |
|
DNA Clean & Concentrator-100 Kit |
Zymo |
D4029 |
|
DNeasy Blood & Tissue Kit |
Qiagen |
69504 |
|
Duplex buffer |
Integrated DNA Technologies |
11-01-03-01 |
|
Methylene blue |
Thermo Scientific |
414240250 |
|
OneTaq 2X Master Mix |
New England Biolabs |
M0482S |
|
Sequencing |
Eurofins Genomics |
n/a |
|
Software - behavior analysis |
Noldus |
EthoVision XT 16 |
|
Software - graphing and statistics |
GraphPad |
Prism 10.4.1 |
|
Software - image analysis |
Public domain |
ImageJ |
|
Software - sequence analysis |
Dotmatics |
SnapGene |
|
Stereomicroscope |
Nikon |
SMZ1270 |
|
Tricaine methanesulfonate |
Western Chemical |
Tricaine-S |
|
Primer - PCR forward |
5’ - GGTAGTCCGGCTCTATTTGAAG - 3’ | |
|
Primer - PCR reverse |
5’ - GGCTTGTTTGTGGTGGTAATG - 3’ | |
|
Primer - sequencing |
5’ - TTGGCATCGTACCATAAAGCATAG - 3’ | |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsObsessive-Compulsive Spectrum Disorders · Genetics and Neurodevelopmental Disorders · Williams Syndrome Research
Description
The SLITRKs are a family of six transmembrane proteins that play critical roles in central nervous system development and function (Aruga and Mikoshiba, 2003; Won et al., 2019; Puranik and Song, 2024). Structurally, SLITRKs exhibit homology with both the SLIT and TRK protein families that have well-established roles in neuronal development and function. Slit ligands guide axons during early development by signaling through robo transmembrane receptors (Brose and Tessier-Lavigne, 2000; Chedotal, 2007; Blockus and Chedotal, 2016). Transmembrane trk receptors transmit signals from neurotrophin ligands to influence neuronal survival and development (Huang and Reichardt, 2003; Deinhardt and Chao, 2014). SLITRK extracellular domains are characterized by leucine-rich repeats (LRRs) characteristic of molecules that guide synapse formation and stability (de Wit et al., 2011; Ko, 2012; Schroeder and de Wit, 2018) and by intracellular domains that include phosphorylation sites characteristic of trk receptors. SLITRKs are expressed in developing and adult vertebrate neural tissues at synaptic contact sites and have been associated with a variety of human neuropsychiatric conditions such as schizophrenia, obsessive compulsive disorder (OCD), and Tourette syndrome (Proenca et al., 2011; Monteiro and Feng, 2016).
*SLITRK5, * one member of the SLITRK family, plays diverse roles in regulating central nervous system processes including neurite outgrowth, synapse formation, dendritic branching, and signal transmission (Yim et al., 2013; Um et al., 2014; Song et al., 2015; Kang et al., 2016; Puranik and Song, 2024). Slitrk5 is expressed in regions of the developing central nervous system during periods of morphogenesis and synaptogenesis and is highly expressed in the adult CA1 region of the hippocampus, occipital and frontal lobes of the brain, spinal cord, and medulla, indicating a plausible association with multiple areas of neuron growth and central nervous system processing (Aruga et al., 2003; Beaubien and Cloutier, 2009; Meyer, 2014; Round et al., 2014). Variants and epigenetic modifications of human SLITRK5 have been linked to behavioral and neurodevelopment conditions including obsessive-compulsive disorder (OCD), Tourette syndrome, attention deficit/hyperactivity disorder (ADHD), conduct disorder (CD), and schizophrenia (Proenca et al., 2011; Song et al., 2017; Salesse et al., 2020; Halvorsen et al., 2021; Chiocchetti et al., 2022; Puranik and Song, 2024). Additionally, Slitrk5 knockout (KO) mice exhibit severe anxiety and OCD-like behaviors such as excessive and harmful self-grooming (Shmelkov et al., 2010). Physiologically, elevated activity in the orbitofrontal cortex has also been observed, consistent with functional imaging findings in humans with OCD, implicating dysregulation of corticostriatal circuitry (Rauch et al., 2007; Ting and Feng, 2011). Increased SLITRK5 is also associated with epilepsy in a rat model (Liu et al., 2023).
The zebrafish ( Danio rerio ) is a cost-effective, genetically accessible, and easily manipulable model system to study nervous system development and neuropsychiatric conditions (Fetcho and Liu, 1998; Stewart et al., 2015; Fontana et al., 2018; Nelson and Granato, 2022). Slitrk5a expression in the developing zebrafish, first detected at 48 hours post fertilization (hpf), is observed in the retina, midbrain, medulla oblongata, valvula cerebelli, pituitary gland, and spinal cord suggesting slitrk5a could contribute to neurogenesis, morphogenesis, and/or synaptogenesis (Round et al., 2014). This study specifically investigated slitrk5a in the zebrafish central nervous system with downstream implications on behaviors associated with neuropsychiatric conditions such as obsessive compulsive disorder (OCD).
To generate mutant zebrafish with altered slitrk5a gene sequences, CRISPR-Cas9 reagents were microinjected into embryonic zebrafish at the single-cell stage ( Fig. 1A ; Hwang et al., 2013; Sorlien et al., 2018). The embryos were examined in early development then grown to adulthood when they could be genotyped to determine if the slitrk5a sequence had been altered. CRISPR-injected, sham-injected control, and uninjected control embryos at 24 hpf displayed similar (p>0.05) average tail coiling events per minute indicating that CRISPR reagents did not alter early motor activity ( Fig. 1B ). At 36 hpf, CRISPR-injected and sham-injected control survived at similar rates (p=0.89) suggesting that CRISPR reagents did not alter survival rates ( Fig. 1C ). The reduced survival rates of injected (sham- and CRISPR-injected) embryos compared to uninjected control embryos are not unexpected given that microinjection necessarily damages the chorion. Embryo lengths ( Fig. 1D ) were also similar (p>0.05) in all comparisons of uninjected (n=41), sham-injected (n=33), and CRISPR-injected (n=30) embryos at 48, 60, and 75 hpf, indicating that gross morphological growth rates were unaffected by microinjection, CRISPR reagents, or the resulting genetic alterations. CRISPR-Cas9 editing of a target gene was not expected to occur successfully in all cells of all embryos. Consequently, changes in slitrk5a sequence could cause growth alterations that are not discernible in an F 0 population prior to genotyping.
Out of a cohort of approximately 50 CRISPR-injected embryos, 30 viable adult fish resulted, six of which were identified as having alterations in the slitrk5a gene sequence on chromosome 15. Three behavioral assays were used to compare adult zebrafish behavior in wild type (WT) and slitrk5a mutant F 0 fish reared from the same clutch of embryos. The novel tank test (NTT) was administered ( Fig. 1E ) to estimate levels of anxiety in zebrafish through their latency to enter the upper half of the new tank (Kysil et al., 2017; Duarte et al., 2019). Typically, fish immediately swim to the bottom of a new tank, begin to explore as they acclimate, and eventually enter the upper portions of the tank, swimming toward the surface. The latency for the fish to swim toward the surface of the water can be used as an estimate for anxiety (Kysil et al., 2017). No significant difference (p=0.10) was observed between latencies for WT (n=9) and mutant fish (n=6) to enter the upper half of the novel tank, suggesting that altering the slitrk5a sequence did not affect this measure of anxiety.
An open field (OF) assay (Johnson and Hamilton, 2017) was used to assess general swimming behavior and thigmotaxis, the tendency of the fish to swim close to the walls of the tank, an anxiety-related behavior. No significant difference was observed in thigmotaxis ( Fig. 1F ) between the WT controls (n=6) and slitrk5a mutant adult fish (n=6) during open field observation (p=0.23), suggesting alterations in the slitrk5a sequence also did not alter this anxiety-related behavior. Moreover, the OF assay revealed no significant differences between the mutant fish and WT controls in average swimming velocity (mutant: 8.8 + 0.96 cm/s; WT: 7.9 + 0.70 cm/s; p=0.45) or total distance swum (mutant: 2510 + 268 cm; WT: 2303 + 203 cm; p=0.55) indicating that mutations in the slitrk5a sequence did not alter general swimming ability or behavior.
Finally, the novel approach test (NAT) was used to assess anxiety-like and OCD-like behaviors. In the NAT, fish are placed into a tank with a novel object to model fear as the fish may perceive the new object as potentially harmful (Johnson and Hamilton, 2017; Hamilton et al., 2017). The path of WT fish demonstrated a consistent peripheral circling patterns with few interactions with the central novel object whereas the paths of slitrk5a mutant fish demonstrated repeated approaches to the novel object, illustrating an OCD-like repetitive checking behavior ( Fig. 1G ). A significant increase in the overall number of visits to the novel object was observed ( Fig. 1H ; p=0.0004) as well as a significant increase in the amount of time spent around the novel object in slitrk5a mutant fish compared to WT fish ( Fig. 1I ; p=0.02). These data suggest that alterations in slitrk5a induced repetitive, OCD-like behaviors in zebrafish. In the NAT, slitrk5a mutant zebrafish interacted significantly more with the novel object; this checking behavior is consistent with the loss of SLITRK5 in mice that led to an OCD-like repetitive behavior of overgrooming (Schmelkov et al., 2010). Additionally, this observation parallels human behavior as some individuals with OCD repetitively check stimuli or objects that could potentially pose a threat even after confirming that no threat exists (APA, 2013).
The creation of F _0 _ slitrk5a mutant zebrafish demonstrates feasibility for CRISPR-Cas9 gene targeting to create mutant fish in which developmental and behavioral analyses can be conducted to examine the roles of slitrk5a in neuronal development and behavior. That a behavioral phenotype of significantly enhanced checking was observed in a small number of likely mosaically edited F 0 fish in this preliminary study warrants investment in future studies to create a homozygous loss-of-function slitrk5a mutant line of zebrafish for analysis of development and behavior.
Methods
**Zebrafish: ** This study followed protocols approved by the Davidson College Animal Care and Use Committee. Adults were housed in a recirculating aquatic system kept at a 28 °C with a 14:10 hour light:dark cycle. Adult fish were fed twice daily with brine shrimp hatched in-house and Aquafeed Z Gel Cubes. Embryos and early larvae were raised in 10 cm Petri dishes with system water supplemented with 0.0005% methylene blue as a gentle antibiotic (Nüsslein-Volhard and Dahm, 2002). Debris and dead embryos were removed daily and the water was replaced. Larvae and juvenile fish were reared in 750 mL plastic beakers with fish system water plus 0.0005% methylene blue until they were able to feed on brine shrimp (typically ~14 days). Water changes were completed every other day and fish were closely monitored until they had grown sufficiently to be moved into recirculating colony system tanks.
** Generation of slitrk5a Mutants Using CRISPR-Cas9: ** A CRISPR crRNA was designed to target the beginning of exon one of the zebrafish *slitrk5a * gene using Integrated DNA Technologies’ (IDT) design portal to maximize specificity and minimize off-target effects. Upon receipt, the Alt-R S.p. Cas9 Nuclease V3 was diluted to 57 μM in Cas9 buffer (20 mM Tris-HCl, 600 mM KCl, 20% glycerol) as previously described (Sorlien et al., 2018; Wu et al., 2018). To form the gRNAs, equal amounts of crRNA and trans-activating (tracr) RNA were mixed, diluted to 57 μM in duplex buffer and annealed by heating to 95 °C for five minutes then cooled on ice. To generate the ribonucleoprotein complex (RNP), equal volumes of annealed gRNAs and Cas9 solutions were mixed, incubated at 37 °C for five minutes then cooled on ice, generating a 28.5 μM RNP solution. Approximately 1 nL of RNP was microinjected into zebrafish embryos at the single-cell stage. The resulting embryos were closely monitored throughout development and reared to adults.
**Early Developmental Analysis: ** Wild-type (AB background) zebrafish were injected with CRISPR-Cas9 reagents targeting *slitrk5a * as described above (CRISPR treatment). In parallel, a cohort of embryos from the same clutch were similarly microinjected with ~1 nL of 0.1% phenol red (sham control) and a cohort of embryos were left untreated (uninjected control). To assess for potential developmental differences, larvae were imaged with a Nikon SMZ1270 stereomicroscope equipped with a digital camera. Early embryonic motor activity was measured by counting the number of spontaneous tail coiling events per minute while embryos were in the chorion at 24 hpf with the observer unaware of the treatment group (de Oliveira et al., 2021; von Hellfield et al., 2023). Tail coiling events were manually scored as visible contractions of the animal's tail during a one minute observation. As a general indication of development, body lengths (anterior-posterior) of embryos at 48, 60, and 75 hpf were measured without knowledge of the treatment group using ImageJ. Additionally, 36 hpf survival rates across the three groups of embryos were measured for four separate experiments.
**gDNA Isolation and Genotyping of Adult Zebrafish: ** At approximately three months of age, samples were obtained of each adult’s caudal fin as previously described (Westerfield, 2007). Genomic DNA (gDNA) was extracted from the tissue using the Qiagen DNeasy Blood & Tissue Kit following manufacturer specifications. Isolated gDNA was then used to amplify the genomic region around *slitrk5a * via standard polymerase chain reaction (PCR) *. * PCR products were purified using the Zymo DNA Clean & Concentrate Kit and then sequenced with Sanger sequencing via Eurofins Genomics. Sequences were aligned to the zebrafish *slitrk5a * reference sequence using SnapGene. Sequencing alignments were analyzed for mutations indicative of non-homologous end-joining (NHEJ) DNA repair initiated by CRISPR-guided Cas9 double-stranded (DS) breaks in the region targeted by the gRNA. A non-targeted portion of the slitrk5a gene adjacent to the target site remained intact with nearly 100% homology to the reference sequence. Mutant individuals with alterations in the slitrk5a sequence were then housed separately from WT adults with intact slitrk5a sequences within the colony where their caudal fins regenerated before behavioral testing.
**Novel Tank Test (NTT): ** The novel tank test (NTT) was administered as previously described to record the latency for isolated adult fish to enter the upper half of a new tank (Kysil et al., 2017; Duarte et al., 2019). Zebrafish were removed from colony tanks, individually placed into 2.0 L transport tanks, and allowed to acclimate for one hour. Acclimatized fish were then individually placed into a 9.5 L tank (32.5 x 21 x 17.5 cm) and video recorded for 10 minutes using EthoVision XT tracking software. If a fish did not enter the upper half of the tank after ten minutes, they were removed and returned to their individual 2.0 L transport tank. The initial latency for the fish to enter to the upper half of the tank was analyzed using EthoVision XT software. Each individual fish was observed in the NTT assay three times.
**Open Field (OF) Observation and the Novel Approach Test (NAT): ** Open field (OF) observations and the novel approach test (NAT) were performed as previously described (Hamilton et al., 2017; Johnson and Hamilton, 2017). Fish were removed from colony tanks and individually placed into 2.0 L transport tanks, with enrichment and allowed to acclimate for one hour. Fish were then placed individually into a 21 L tank (37.5 x 31 x 17.5 cm) and video recorded for five minutes. This five minute acclimation period served as the open field (OF) observation. Subsequently, a novel object (three yellow, plastic 1.5 mL tubes glued to a 60 mm plastic petri dish lid) was placed in the center of the tank and observation continued for an additional five minutes. Internal EthoVision XT analysis was performed to calculate the dependent variables in this study. Thigmotaxis was defined as the peripheral region of the tank within 6 cm of the edges on all four sides. Visits to the novel object were defined as the fish approaching the central object within 5 cm. Each individual fish was observed on the OF/NAT assay twice.
**Statistical Analysis: ** Statistical analysis was performed using GraphPad Prism 10. Survival comparisons used paired t-tests. Motor activity and embryo length used ANOVA multivariate analysis. Comparisons for all three behavioral analyses (NAT, NTT, OF) used unpaired t-tests. A p-value of ≤0.05 was considered significant (*) in all analyses.
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aruga J Mikoshiba K 200391 Identification and characterization of Slitrk, a novel neuronal transmembrane protein family controlling neurite outgrowth.Mol Cell Neurosci 2411044-743111712910.1016/s 1044-7431(03)00129-514550773 · doi ↗ · pubmed ↗
- 2Aruga J Yokota N Mikoshiba K 2003102 Human SLITRK family genes: genomic organization and expression profiling in normal brain and brain tumor tissue.Gene 3150378-1119879410.1016/s 0378-1119(03)00715-714557068 · doi ↗ · pubmed ↗
- 3Beaubien F Cloutier JF 2009121 Differential expression of Slitrk family members in the mouse nervous system.Dev Dyn 238121058-83883285329610.1002/dvdy.2216019924824 · doi ↗ · pubmed ↗
- 4Blockus H Chédotal A 201691 Slit-Robo signaling.Development 143170950-19913037304410.1242/dev.13282927578174 · doi ↗ · pubmed ↗
- 5Brose K Tessier-Lavigne M 200021 Slit proteins: key regulators of axon guidance, axonal branching, and cell migration.Curr Opin Neurobiol 1010959-43889510210.1016/s 0959-4388(99)00066-510679444 · doi ↗ · pubmed ↗
- 6Chédotal A 2007 Slits and their receptors.Adv Exp Med Biol 6210065-2598658010.1007/978-0-387-76715-4_518269211 · doi ↗ · pubmed ↗
- 7Chiocchetti AG Yousaf A Waltes R Bernhard A Martinelli A Ackermann K Haslinger D Rotter B Krezdorn N Konrad K Kohls G Vetro A Hervas A Fernández-Rivas A Freitag CM 2022128 The methylome in females with adolescent Conduct Disorder: Neural pathomechanisms and environmental risk factors.P Lo S One 171e 0261691 e 026169110.1371/journal.pone.026169135089926 PMC 8797262 · doi ↗ · pubmed ↗
- 8de Wit J Hong W Luo L Ghosh A 201175 Role of leucine-rich repeat proteins in the development and function of neural circuits.Annu Rev Cell Dev Biol 271081-070669772910.1146/annurev-cellbio-092910-15411121740233 · doi ↗ · pubmed ↗