Bioinformatics analysis identifies Mot2 protein as a potential regulator of autophagy in Saccharomyces cerevisiae
Emmanuella Wesome Avogo, Nicholas A. Burlingame, Swaroopa Badenahalli Narasimhaiah, Elizabeth Delorme-Axford

TL;DR
This study uses bioinformatics to find that the Mot2 protein in yeast may regulate autophagy and cell survival.
Contribution
The study identifies Mot2p as a novel regulator of autophagy and ATG8 in Saccharomyces cerevisiae.
Findings
Mot2p was identified as a potential regulator of autophagy-related genes in yeast.
Mot2p is linked to cell survival and regulation of ATG8.
Bioinformatics analysis revealed Mot2p as a novel factor modulating autophagy.
Abstract
Macroautophagy/autophagy is a conserved mechanism of cellular degradation and recycling. Autophagy is a multi-step process that must be precisely regulated at multiple levels (transcriptional, post-transcriptional, translational, and post-translational). However, there is a gap in our understanding of the molecular details of these mechanisms. Therefore, characterization of factors modulating autophagy and autophagy-related ( ATG ) genes is an important area for investigation. Here, we used a bioinformatics approach to screen 37 yeast ATG genes using the YEASTRACT database to identify potential regulators. Through our selection criteria, we discovered one novel factor—Mot2p. Our findings support that the yeast Mot2 protein is a regulator of cell survival, ATG8 , and potentially, autophagy.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
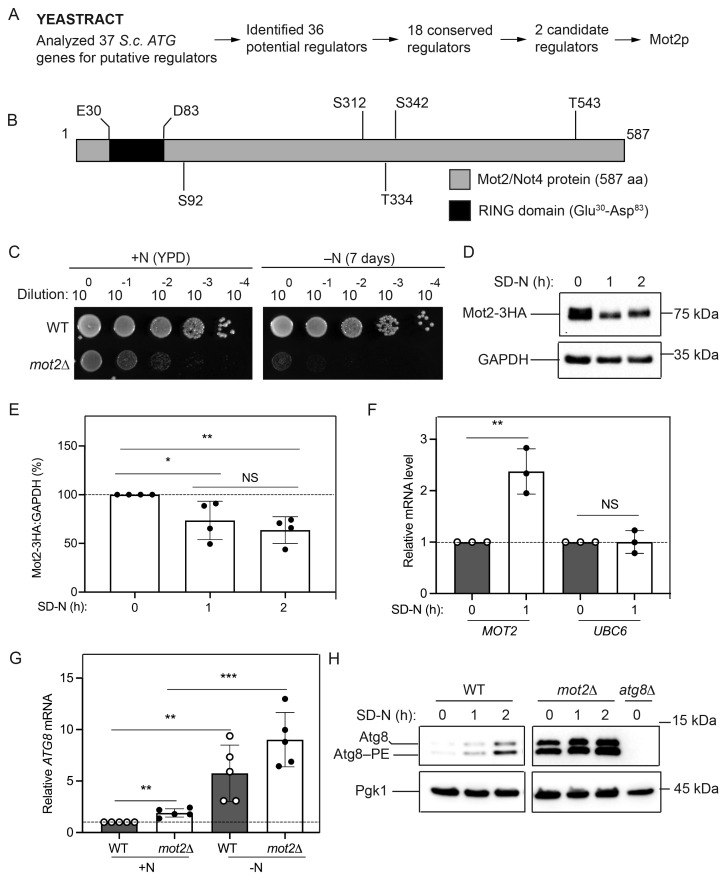 Figure 1
Figure 1|
| ||
|
|
|
|
|
BY4742 |
MATα
|
Horizon Discovery |
|
EWA018 |
SEY6210,
|
This study |
|
|
BY4742,
|
Horizon Discovery |
|
YAB369 |
YTS158,
|
(Delorme-Axford, Abernathy et al. 2018) |
|
YTS158 |
BY4742,
|
(He, Song et al. 2006) |
|
SEY6210 |
MAT
|
(Robinson, Klionsky et al. 1988) |
|
| ||
|
|
|
|
|
|
GAAGGCCATCTTCATTTTTGTC |
(Bernard, Jin et al. 2015) |
|
|
TTCTCCTGAGTAAGTGACATAC |
(Bernard, Jin et al. 2015) |
|
|
ACGAAAACTCCCACCCAACC |
This study |
|
|
CAAAGATACCCGGTGGAGGG |
This study |
|
|
CGCAACTTCAAAGCATCATTGAATCGC |
(Bernard, Jin et al. 2015) |
|
|
GGGGCTTATTAGTGGGAGTAGAGG |
(Bernard, Jin et al. 2015) |
|
|
GATACTTGGAATCCTGGCTGGTCTGTCTC |
(Teste, Duquenne et al. 2009) |
|
|
AAAGGGTCTTCTGTTTCATCACCTGTATTTGC |
(Teste, Duquenne et al. 2009) |
|
| ||
|
|
|
|
|
mouse monoclonal anti-Atg8 (G-10) |
Cat# sc-373963 (Santa Cruz Biotechnology) |
1:1,000 |
|
mouse monoclonal anti-GAPDH (1E6D9) |
Cat# 60004-1-Ig (Proteintech) |
1:20,000 |
|
rabbit polyclonal anti-HA tag |
Cat# 51064-2-AP (Proteintech) |
1:10,000 |
|
mouse monoclonal anti-Pgk1 (22C5D8) |
Cat# 459250 (Invitrogen) |
1:5,000 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAutophagy in Disease and Therapy
Description
Canonical macroautophagy/autophagy is a highly conserved mechanism of cellular degradation and recycling. Basal autophagy is essential for homeostasis, but is upregulated under stressful conditions (such as nutrient limitation). Currently, >40 autophagy-related ( ATG ) genes have been identified in yeast. This complexity requires strict modulation of the autophagy machinery at multiple levels (transcriptional, post-transcriptional, translational, and post-translational). Despite the need for precision, there is a gap in our understanding of the factors and mechanisms regulating autophagy in the cell. In humans, perturbation of autophagy (i.e., too much or too little) can have deleterious effects on cell health and survival, contributing to disease pathogenesis (Klionsky, Petroni et al. 2021). Therefore, identifying molecular mechanisms modulating ATG gene expression and autophagy is an important area for investigation.
The primary goal of this study was to identify and characterize a novel regulator of ATG genes and autophagy in the yeast Saccharomyces cerevisiae . Using the online database YEASTRACT (Yeast Search for Transcriptional Regulators And Consensus Tracking; www.yeastract.com ) (Teixeira, Monteiro et al. 2006), we screened ATG1
24 , ATG26
27 , ATG29 , ATG31
34 , ATG36 , ATG38
42 to identify potential transcriptional regulators ( Figure 1A ). *ATG25 * (Monastyrska, Kiel et al. 2005), ATG28 (Stasyk, Stasyk et al. 2006), ATG30 (Farré, Manjithaya et al. 2008), ATG35 (Nazarko, Nazarko et al. 2011), and ATG37 (Nazarko, Ozeki et al. 2014) are only present in methylotrophic yeast, and no equivalent has been identified in S. cerevisiae . YEASTRACT analysis of 37 S. cerevisiae *ATG * genes yielded 36 potential transcription factors that were predicted to regulate >50% of the ATG genes examined ( Figure 1A ). Next, using the Saccharomyces Genome Database ( https://www.yeastgenome.org/ ), we found that 18 of the 36 potential regulators are conserved from yeast to humans ( Figure 1A ). Of the remaining 18, we excluded factors that had published association(s) with autophagy. Of these, we were left with 2 candidates—Spt23p and Mot2p. Spt23p has a human homolog—ANKFY1 *. * ANKFY1 has a published association with autophagy (Park, Peng et al. 2016, Wei, Fu et al. 2024), indicating that our approach is a viable method for identifying potential autophagy regulators. Only a single candidate made it past all of the screening criteria—Mot2p.
The Mot2/Not4 (modulator of transcription) protein was first characterized as a transcription factor that represses basal transcription of certain mating-specific genes, but was postulated to have a more global effect on gene expression (Cade and Errede 1994). Mot2p is also a phosphoprotein with five identified phosphorylation sites–Ser92, Ser312, Thr334, Ser342, and Thr 543 (Lau, Mulder et al. 2010) ( Figure 1B ). The Mot2 protein has a RING domain ( Figure 1B ) and functions as an E3 ligase to modulate cell stress responses (Albert, Hanzawa et al. 2002, Mulder, Inagaki et al. 2007). Mot2p was later identified as a subunit of the yeast Ccr4-Not complex (Liu, Badarinarayana et al. 1998). The Ccr4-Not complex regulates transcription (Liu, Badarinarayana et al. 1998, Badarinarayana, Chiang et al. 2000, Denis, Chiang et al. 2001), mRNA turnover (Badarinarayana, Chiang et al. 2000), and translational repression (Preissler, Reuther et al. 2015). Yin and colleagues investigated Ccr4-Not complex members Ccr4p and Pop2p, demonstrating bidirectional roles in autophagy regulation (Yin, Zhang et al. 2023). However, to the best of our knowledge, no one has examined the role of Mot2p in autophagy.
Nitrogen starvation is a robust trigger for autophagy in yeast (Delorme-Axford, Guimaraes et al. 2015). Autophagy-deficient cells display reduced viability under prolonged nitrogen starvation conditions (Tsukada and Ohsumi 1993, Bernard, Jin et al. 2015). Excessive autophagy results in a similar phenotype (Hu, McQuiston et al. 2015, Delorme-Axford, Wen et al. 2023). To examine the importance of the Mot2 protein under prolonged nitrogen starvation condition, we monitored the cell survival phenotype of cells lacking MOT2 ( Figure 1C ). Loss of MOT2 decreased cell survival under nutrient-rich conditions ( Figure 1C ), consistent with what has been observed by others (Lau, Mulder et al. 2010, Preissler, Reuther et al. 2015). Additionally, mot2 Δ cells do not survive prolonged nitrogen starvation (7 days; Figure 1C ), suggesting that extended starvation in the absence of MOT2 is detrimental.
Positive regulators are typically upregulated in response to autophagy inducing conditions (Xie, Nair et al. 2008, Yao, Delorme-Axford et al. 2015). Conversely, negative regulators are typically inactivated following autophagy (He and Klionsky 2009). To determine how nitrogen starvation impacts Mot2 protein levels, MOT2 was chromosomally tagged at its C terminus with the hemagglutinin (3HA) epitope. Endogenous Mot2-3HA fusion protein levels were assessed by a time course of nitrogen starvation (0, 1, and 2 h) and examined by western blot analysis ( Figure 1D, E). Mot2-3HA fusion protein levels significantly decreased with nitrogen starvation in WT cells (>35% by 2 h; Figure 1D, E). In contrast, when cells are starved for nitrogen, MOT2 mRNA levels are upregulated (~2-fold) in wild-type (WT) cells ( Figure 1F ), suggesting that MOT2 /Mot2p may be differentially regulated under autophagy-inducing conditions. Notably, we observed a similar phenomenon with the yeast metabolic transcription factor Stb5p (Delorme-Axford, Wen et al. 2023). As a control, we also examined *UBC6 * (a gene with no known connection to autophagy) expression levels during nitrogen starvation; no significant differences in *UBC6 * levels were noted ( Figure 1F ).
During autophagy induction in yeast, ATG8 levels increase (Bartholomew, Suzuki et al. 2012). Our YEASTRACT screen identified *ATG8 * as a predicted target of Mot2p; there are multiple consensus sites along the *ATG8 * promoter that lie within regions -302 to -163 upstream of the ATG +1 start site. Therefore, we investigated whether Mot2p could modulate ATG8 mRNA expression ( Figure 1G ). During nutrient-rich conditions, *ATG8 * levels were significantly higher (~2-fold) in mot2 Δ cells compared to WT ( Figure 1G ). As expected, we observed that *ATG8 * levels were upregulated when WT cells were starved for nitrogen (~6-fold; Figure 1G ). In mot2 Δ cells, ATG8 levels were enhanced (~9-fold) when cells were starved for nitrogen compared to nutrient-rich conditions (~2-fold; Figure 1G ). When cells were starved for nitrogen, we noted that ATG8 levels were elevated in the mot2 Δ strain compared to WT, although the difference was not statistically significant ( Figure 1G ).
Atg8 protein exists as two species in the cell – a non-lipidated soluble form and a lipidated phosphatidylethanolamine (PE)-conjugated species (Cheong and Klionsky 2008). Furthermore, Atg8 protein levels increase when autophagy is induced (Huang, Scott et al. 2000, Xie, Nair et al. 2008). Thus, assessing the total amount of Atg8 protein and its lipidation status is an indicator of autophagy (Delorme-Axford, Abernathy et al. 2018). Given that we identified Mot2p as a potential regulator of *ATG * genes ( Figure 1A ), and cells lacking MOT2 do not survive prolonged nitrogen starvation ( Figure 1C ), we examined whether loss of MOT2 had an effect on autophagy and Atg8 protein levels in mot2 Δ cells ( Figure 1H ). We found that cells lacking MOT2 had an increased amount of Atg8 protein relative to the WT under nutrient-rich and starved conditions ( Figure 1H ), supporting the idea that Mot2p negatively regulates Atg8 protein levels, and potentially, autophagy.
Here we present data supporting a role for Mot2p as a regulator of cell survival, ATG8 , and potentially, autophagy. These findings add to the repertoire of studies that have utilized YEASTRACT (Teixeira, Monteiro et al. 2006) to identify regulators of ATG genes and autophagy (Yao, Delorme-Axford et al. 2015, Delorme-Axford, Abernathy et al. 2018, Delorme-Axford, Wen et al. 2023). These results serve as a basis for future work aimed at elucidating the role of Mot2p in the regulation of ATG8 /Atg8p and autophagy.
Methods
** *Yeast Strains, Media, and Cell Culture: * ** Yeast cells were grown in YPD (1% yeast extract, 2% peptone, and 2% glucose) medium from Gibco (A1374501). To induce autophagy, cells were grown to mid-log phase in YPD, then shifted to nitrogen starvation medium (SD-N; 0.17% yeast nitrogen base without ammonium sulfate or amino acids and 2% glucose) for the specified time points. Chromosome tagging with 3HA was performed using established methods (Longtine, McKenzie et al. 1998).
** *Yeast Growth Assay: * ** Yeast growth assays were performed as previously described (Yin, Zhang et al. 2023) with the following modifications. Yeast cells were cultured in YPD to mid-log phase and then shifted to SD-N for the time points indicated. An aliquot of cells (1 OD 600 unit) was removed from each culture and serially diluted. Each dilution (5 μl) was spotted on YPD plates. Cells were grown at 30°C for 2 days before being imaged with a ChemiDocTouch imaging system (Bio-Rad).
** RNA and Real-Time Quantitative PCR (RT-qPCR): ** Yeast cells were cultured in YPD to mid-log phase and then shifted to SD-N (1 h) for autophagy induction. Cells (1 OD 600 unit) were collected at the indicated time point, and the pellets were frozen in liquid nitrogen. Total RNA was extracted using the NucleoSpin RNA extraction kit (Clontech, 740955.250). Reverse transcription was carried out using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems/Thermo Fisher Scientific, 4368814). For each sample, 1 µg RNA was used for cDNA synthesis. RT-qPCR was performed using the Power SYBR Green PCR Master Mix (Applied Biosystems/Thermo Fisher Scientific, 4367659) in a CFX Opus 96 (Bio-Rad, 12011319) real-time PCR machine. For all RT-qPCR experiments, melt curves were run after the PCR cycles to verify primer specificity. Relative gene expression was calculated using the 2 ^−ΔΔCT^ method (Livak and Schmittgen 2001), normalized to *SLD3 * levels.
** SDS-PAGE and Western Blots: ** SDS-PAGE and western blots were performed as previously described (Cheong and Klionsky 2008, Delorme-Axford, Tasmi et al. 2023). Western blots were visualized using an Azure 600 (Azure Biosystems) or an iBright (Thermo Fisher Scientific) imaging system. Densitometry for western blots was performed using ImageJ ( https://imagej.net/ij/ ).
** Statistical analysis: ** The two-tailed unpaired *t * test was used to determine statistical significance with GraphPad Prism (GraphPad Software, USA). For Figure 1, p values are as follows: * p <0.05; ** p <0.01; *** p <0.001; NS indicates not significant. A p value < 0.05 was considered significant.
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Albert Thomas K. Hanzawa Hiroyuki Legtenberg Yvonne I.A. de Ruwe Marjolein J. van den Heuvel Fiona A.J. Collart Martine A. Boelens Rolf Timmers H.Th.Marc 200221 Identification of a ubiquitin-protein ligase subunit within the CCR 4-NOT transcription repressor complex The EMBO Journal 2130261-418935536410.1093/emboj/21.3.35511823428 PMC 125831 · doi ↗ · pubmed ↗
- 2Badarinarayana Vasudeo Chiang Yueh-Chin Denis Clyde L 200071 Functional Interaction of CCR 4-NOT Proteins With TATAA-Binding Protein (TBP) and Its Associated Factors in Yeast Genetics 15531943-26311045105410.1093/genetics/155.3.104510880468 PMC 1461164 · doi ↗ · pubmed ↗
- 3Bartholomew Clinton R. Suzuki Tsukasa Du Zhou Backues Steven K. Jin Meiyan Lynch-Day Melinda A. Umekawa Midori Kamath Avani Zhao Mantong Xie Zhiping Inoki Ken Klionsky Daniel J. 2012625 Ume 6 transcription factor is part of a signaling cascade that regulates autophagy Proceedings of the National Academy of Sciences 109280027-8424112061121010.1073/pnas.1200313109 PMC 339650622733735 · doi ↗ · pubmed ↗
- 4Bernard Amélie Jin Meiyan González-Rodríguez Patricia Füllgrabe Jens Delorme-Axford Elizabeth Backues Steven K. Joseph Bertrand Klionsky Daniel J. 201531 Rph 1/KDM 4 Mediates Nutrient-Limitation Signaling that Leads to the Transcriptional Induction of Autophagy Current Biology 2550960-982254655510.1016/j.cub.2014.12.04925660547 PMC 4348152 · doi ↗ · pubmed ↗
- 5Cade Rebecca M. Errede Beverly 199451 MOT 2 Encodes a Negative Regulator of Gene Expression That Affects Basal Expression of Pheromone-Responsive Genes in Saccharomyces cerevisiae Molecular and Cellular Biology 1451098-55493139314910.1128/mcb.14.5.3139-3149.19948164669 PMC 358681 · doi ↗ · pubmed ↗
- 6Cheong Heesun Klionsky Daniel J. 2008 Chapter 1 Biochemical Methods to Monitor Autophagy‐Related Processes in Yeast Methods in Enzymology 0076-687912610.1016/s 0076-6879(08)03201-119185709 · doi ↗ · pubmed ↗
- 7Delorme-Axford Elizabeth Abernathy Emma Lennemann Nicholas J. Bernard Amélie Ariosa Aileen Coyne Carolyn B. Kirkegaard Karla Klionsky Daniel J. 2018321 The exoribonuclease Xrn 1 is a post-transcriptional negative regulator of autophagy Autophagy 1451554-862789891210.1080/15548627.2018.144164829465287 PMC 6070002 · doi ↗ · pubmed ↗
- 8Delorme-Axford Elizabeth Guimaraes Rodrigo Soares Reggiori Fulvio Klionsky Daniel J. 201531 The yeast Saccharomyces cerevisiae: An overview of methods to study autophagy progression Methods 751046-202331210.1016/j.ymeth.2014.12.00825526918 PMC 4355233 · doi ↗ · pubmed ↗