Salt chemotaxis and its plasticity in hermaphroditic nematodes
Akane Matsumura, Yuzuha Komachiya, Ayaka Sugiyama, Mayuko Tanuma, Hayao Ohno

TL;DR
This study explores salt chemotaxis in several nematode species, finding that most show similar abilities to C. elegans, suggesting they could be useful for future research.
Contribution
The study identifies multiple nematode species with salt chemotaxis plasticity, expanding potential model organisms for chemosensation research.
Findings
Most nematode species tested showed salt chemotaxis plasticity similar to C. elegans.
The results highlight the biological importance of salt chemotaxis across species.
These species could serve as alternative models for future chemosensation studies.
Abstract
Salt chemotaxis in the nematode Caenorhabditis elegans has been used as a model to study chemosensation, behavior, and learning and memory. To investigate whether other nematode species could serve as alternative models, we examined salt chemotaxis plasticity in five androdioecious nematode species— Caenorhabditis briggsae , Caenorhabditis tropicalis , Oscheius myriophilus , Oscheius tipulae , and Pristionchus pacificus —all isolated as wild type. Most strains exhibited salt chemotaxis plasticity similar to that of C. elegans , underscoring the biological importance of this ability and supporting their potential use in future research on salt chemotaxis.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
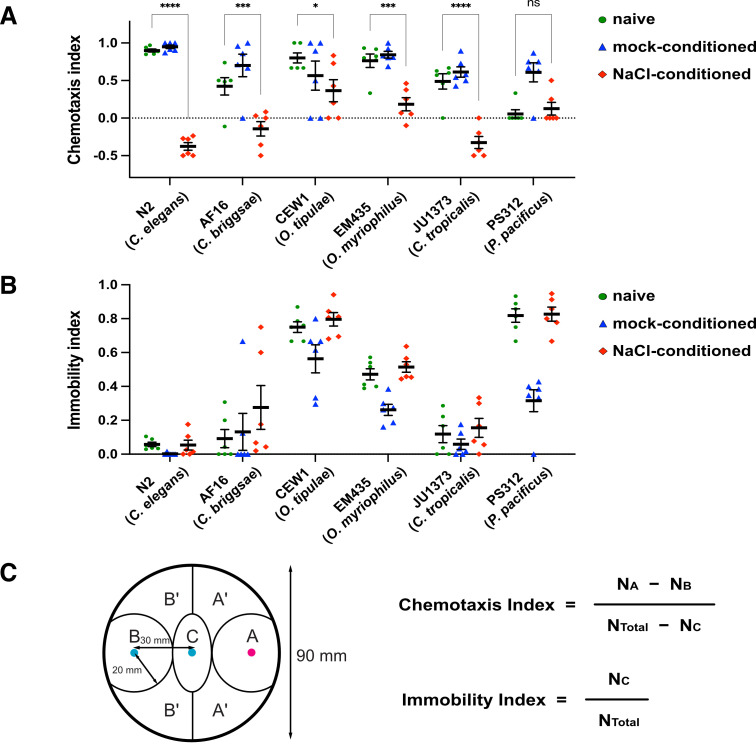 Figure 1
Figure 1|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCephalopods and Marine Biology · Genetics, Aging, and Longevity in Model Organisms
Description
The nematode * Caenorhabditis elegans * is attracted to salt concentrations associated with prior feeding experience and avoids salt concentrations associated with starvation (Luo et al., 2014; Kunitomo et al., 2013). The standard nematode growth medium (NGM), widely used to culture * C. elegans * , contains approximately 50 mM NaCl. Consequently, * C. elegans * worms cultured on NGM plates show attraction to NaCl when placed on agar plates containing a low concentration of NaCl. However, when exposed to NaCl under starvation conditions, they learn to avoid it. This behavioral strategy serves as an experimental system to investigate the molecular mechanisms underlying associative learning (e.g., Tomioka et al., 2006). In this study, we examined salt chemotaxis behavior and its plasticity in nematode species other than * C. elegans * , assessing their potential as models for studying learning and memory.
For research on salt chemotaxis learning, an ideal model nematode should meet the following criteria: (1) it can be cultured using standard methods established for * C. elegans * —enabling laboratories with existing * C. elegans * culture systems to use identical media and reagents; (2) it is androdioecious, allowing easy propagation and crossing, as well as the straightforward isolation of homozygous mutants through self-fertilization; (3) wild isolates are readily accessible through repositories such as the * Caenorhabditis * Genetics Center (CGC); (4) it exhibits salt chemotaxis and associated behavioral plasticity comparable to * C. elegans * , such that experimental conditions optimized for * C. elegans * can be directly applied without extensive re-optimization; and (5) it demonstrates normal locomotion on assay plates, allowing discrimination between defects in chemotaxis behavior (Che mutants) and locomotor abnormalities (Unc mutants). To satisfy criteria (1) through (3), we selected five androdioecious species: * Caenorhabditis briggsae * AF16 , * Caenorhabditis tropicalis * JU1373 , Oscheius myriophilus EM435 , * Oscheius tipulae * CEW1 , and * Pristionchus pacificus * PS312 . We assessed both naive salt chemotaxis and the plasticity of salt chemotaxis following starvation conditioning in these strains. As a control, we used the * C. elegans * N2 strain.
Fig. 1 shows salt preference ( Fig. 1A ) and immobility ( Fig. 1B ) measured after cultivation on NGM (naive), after starvation conditioning without NaCl (mock conditioning), and after starvation conditioning with NaCl (NaCl conditioning). All wild-type strains examined showed plasticity in salt chemotaxis ( Fig. 1A ), suggesting the biological significance of this ability. However, some strains displayed behaviors different from * C. elegans * N2 . For example, CEW1 displayed a weaker avoidance of NaCl following NaCl conditioning. PS312 exhibited a tendency to aggregate and remain immobile on the assay plates under both naive and NaCl-conditioning conditions, resulting in a lack of salt chemotaxis. For research purposes, JU1373 among * Caenorhabditis * species and EM435 among non- * Caenorhabditis * species appear suitable as model strains because they demonstrate salt chemotaxis similar to N2 and remain active on assay plates. Nonetheless, the other strains could also be effectively studied for salt chemotaxis by tailoring experimental conditions to each strain. Research on the nervous systems of nematode species beyond * C. elegans * is beginning to progress (e.g., Toker et al., 2025), and investigations into P. pacificus have started to characterize genes and neurons involved in salt chemotaxis (Mackie et al., 2025). Utilizing salt chemotaxis learning in non- * C. elegans * species may advance our understanding of the mechanisms underlying learning and memory.
Methods
The nematode strains were cultivated at 20°C on NGM plates (Brenner, 1974) seeded with E. coli HB101 as the bacterial food source. Salt chemotaxis learning assay was performed as described (Tomioka et al., 2006). A 9 cm agar assay plate, ~2 mm thick and composed of 5 mM KPO 4 (pH 6.0), 1 mM CaCl 2 , 1 mM MgSO 4 , and 2% agar was used. To establish a salt gradient, an agar plug, which was 5 mm in diameter and 6 mm in height and contained 100 mM NaCl, was placed near the edge of the plate and left overnight. Just prior to placing the worms, 1 µL of 0.5 M sodium azide was spotted both at the location of the salt gradient peak and at the opposite side of the plate. For learning assays, 50–150 young adult worms cultivated on NGM plates were collected and washed three times with CTX buffer (5 mM KPO 4 [pH 6.0], 1 mM CaCl 2 , 1 mM MgSO 4 , 0.05% gelatin). The worms were then transferred to a conditioning buffer of identical composition, either containing 20 mM NaCl (NaCl conditioning) or lacking NaCl (mock conditioning), and incubated with rotation at 22°C for one hour. Following conditioning, the worms were placed at the center of the assay plate and incubated at 22°C for 30 minutes. The chemotaxis index was calculated as (N A – N B ) / (N Total – N C ), where N A represents the number of worms found within 20 mm of the salt gradient peak, N B is the number within 20 mm of the control spot, N Total is the total number of worms on the plate, and N C is the number within the central elliptical region (20 mm short axis by 40 mm long axis) ( Fig. 1C ). The immobility index was defined as the ratio N C / N Total ( Fig. 1C ).
Reagents
Strains used in this study:
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Brenner S 197451 The genetics of Caenorhabditis elegans.Genetics 7710016-6731719410.1093/genetics/77.1.714366476 PMC 1213120 · doi ↗ · pubmed ↗
- 2Kunitomo H Sato H Iwata R Satoh Y Ohno H Yamada K Iino Y 2013 Concentration memory-dependent synaptic plasticity of a taste circuit regulates salt concentration chemotaxis in Caenorhabditis elegans.Nat Commun 42210221010.1038/ncomms 321023887678 · doi ↗ · pubmed ↗
- 3Luo L Wen Q Ren J Hendricks M Gershow M Qin Y Greenwood J Soucy ER Klein M Smith-Parker HK Calvo AC Colón-Ramos DA Samuel AD Zhang Y 201464 Dynamic encoding of perception, memory, and movement in a C. elegans chemotaxis circuit.Neuron 8250896-62731115112810.1016/j.neuron.2014.05.01024908490 PMC 4082684 · doi ↗ · pubmed ↗
- 4Mackie M Le VV Carstensen HR Kushnir NR Castro DL Dimov IM Quach KT Cook SJ Hobert O Chalasani SH Hong RL 2025630 Evolution of lateralized gustation in nematodes.Elife 1410.7554/e Life.10379640586702 PMC 12208668 · doi ↗ · pubmed ↗
- 5Toker IA Ripoll-Sánchez L Geiger LT Sussfeld A Saini KS Beets I Vértes PE Schafer WR Ben-David E Hobert O 2025523 Divergence in neuronal signaling pathways despite conserved neuronal identity among Caenorhabditis species.Curr Biol 35120960-982229272945.e 710.1016/j.cub.2025.05.03640412379 PMC 7617994 · doi ↗ · pubmed ↗
- 6Tomioka M Adachi T Suzuki H Kunitomo H Schafer WR Iino Y 200697 The insulin/PI 3-kinase pathway regulates salt chemotaxis learning in Caenorhabditis elegans.Neuron 5150896-627361362510.1016/j.neuron.2006.07.02416950159 · doi ↗ · pubmed ↗