Storage stability of non-encapsulated pneumococci in saliva is dependent on null-capsule clade, with strains carrying aliC and aliD showing a competitive disadvantage during culture enrichment
Claire S. Laxton, Orchid M. Allicock, Chikondi Peno, Tzu-Yi Lin, Alidia M. Koelewijn, Femke L. Toekiran, Luna Aguilar, Anna York, Anne L. Wyllie

TL;DR
Non-encapsulated pneumococci in saliva show varying stability depending on their genetic makeup, with some strains being less detectable under certain storage conditions.
Contribution
The study reveals that non-encapsulated pneumococci strains with aliC and aliD are less stable during culture enrichment, affecting their detection in saliva-based studies.
Findings
NCC2 strains (with aliC and aliD) showed lower and less stable detection compared to NCC1 strains during culture enrichment.
Extraction-free detection methods provided more stable results for NESp in saliva, especially for NCC2 strains.
NCC2 strains may be underrepresented in carriage studies due to reduced resilience during culture-based detection.
Abstract
Non-encapsulated Streptococcus pneumoniae (NESp) represent up to 19% of circulating pneumococci and exhibit high rates of genetic exchange and antimicrobial resistance. Saliva is increasingly used as a pneumococcal carriage study specimen, and we recently developed a qPCR assay to enhance carriage surveillance and characterization of NESp in saliva. Previous work has established that pneumococci remain viable in unsupplemented saliva for extended periods under various conditions. However, these findings may not be applicable to NESp. Therefore, to ensure the robustness of NESp detection in saliva-based carriage studies, we evaluated the impact of transport and storage conditions of saliva samples on NESp detection. Six NESp strains from two clinically relevant NESp null-capsule clades (NCCs), NCC1 (carrying pspK) and NCC2 (carrying aliC and aliD), were spiked into pneumococcus…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
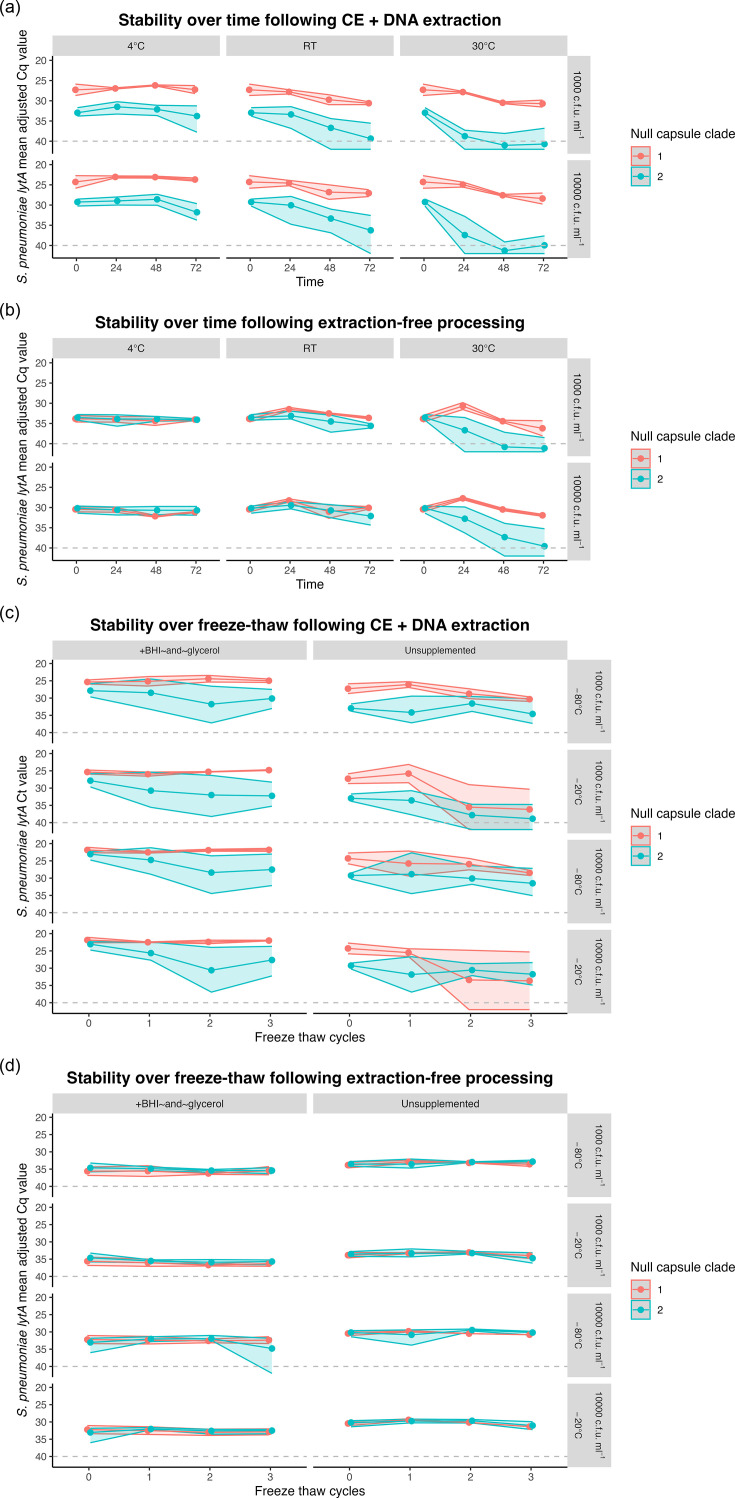 Fig. 1
Fig. 1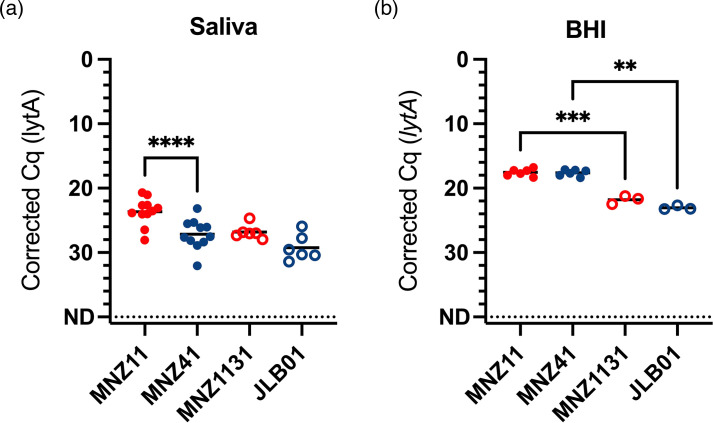 Fig. 2
Fig. 2| Strain | NCC, MLST | Source | Whole-genome sequence accession |
|---|---|---|---|
| MNZ41 | NCC2b, 6153 | Carriage isolate, South Korea [ | |
| MNZ11 | NCC1, 6151 | Carriage isolate, South Korea [ | |
| MNZ85 | NCC2a, 2315 | Carriage isolate, South Korea [ | ASJF000030000 [ |
| C144.66 | NCC1, 9570 | Adenoiditis isolate, USA [ | |
| D37 | NCC2, NF | Carriage isolate, Israel [ | |
| D48 | NCC2, NF | Carriage isolate, Israel [ | |
| MNZ1131 | NCC1, 6151 | MNZ11 | – |
| JLB01 | NCC2b, 6153 | MNZ41 | – |
| Processing method | Strain | Storage conditions* (% ND) | Freeze-thaw† (% ND) | Total (% ND) | |
|---|---|---|---|---|---|
| Unsupplemented | +BHI and glycerol | ||||
| CE and DNA extraction |
| 0/48 (0.0%) | 4/32 (12.5%) | 0/32 (0.0%) | 4/112 (3.6%) |
|
| 18/96 (18.8%) | 3/64 (4.7%) | 0/64 (0.0%) | 21/224 (9.4%) | |
| Extraction-free |
| 0/48 (0.0%) | 0/28 (0.0%) | 0/32 (0.0%) | 0/108 (0.0%) |
|
| 10/96 (10.4%) | 0/59 (0.0%) | 1/64 (1.6%) | 11/219 (5.0%) | |
| Total (% ND) | 28/288 (9.7%) | 7/183 (3.8%) | 1/192 (0.5%) | 36/663 (5.4%) | |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPneumonia and Respiratory Infections · Inhalation and Respiratory Drug Delivery · Protein purification and stability
Introduction
The upper respiratory tract commensal bacterium, Streptococcus pneumoniae (pneumococcus), remains the leading global cause of morbidity and mortality from lower respiratory infections. The introduction of pneumococcal conjugate vaccines (PCVs) has led to a substantial reduction in pneumococcal disease caused by up to 21 of the 100+ pneumococcal serotypes described to date [12]. Concomitantly, PCV use has driven shifts in pneumococcal population dynamics, leading to increased prevalence of non-vaccine-targeted serotypes and non-encapsulated Streptococcus pneumoniae (NESp), which lack a polysaccharide capsule [36]. Carriage prevalence of NESp accounts for 3–19% of circulating pneumococcal strains [78]. This is of particular concern due to their heightened propensity for genetic exchange, including of antibiotic resistance elements, leading to high rates of multidrug resistance among NESp strains [917]. It is therefore important that active surveillance of the pneumococcal landscape includes both typable and non-typable pneumococcal strains.
NESp are divided into two groups: group I, which retains non-functional capsular genes, and group II, which completely lacks capsular genes in the cps locus [18]. Group II NESp are further classified into NCCs based on the presence of specific genetic elements within the cps locus, including pspK (NCC1), aliC and aliD (NCC2, referred to as Swiss_NT by SeroBA 1.0 [19]), only aliD (historically NCC3) or transposable elements alone (NCC4) [7,2022]. The genes present in NCC1 and NCC2 strains, pspK, aliC and aliD, are virulence factors which can compensate for the lack of polysaccharide capsule. AliC and AliD regulate the expression of choline-binding protein AC (CbpAC), which helps reduce C3b deposition on the bacterial surface, thereby enhancing resistance to classical complement-mediated clearance [2324]. PspK increases biofilm formation and bacterial adherence to epithelial cells. In turn, this reduces secretory IgA-mediated nasopharyngeal clearance and enhances transmission during influenza co-infection [122025].
Extensive research has been conducted on encapsulated pneumococci due to the critical role of the capsule in strain virulence. While the absence of capsule generally reduces virulence, NCC1 and NCC2 strains have been associated with conjunctivitis outbreaks, otitis media and occasional cases of invasive pneumococcal diseases [7,14, 2629]. Pneumococcal carriage studies often neglect to further classify serologically non-typable strains; thus, the prevalence of NCC1 and NCC2 NESp is most likely underreported [571330]. We recently sought to remedy this by developing a multiplex-qPCR method to simplify the classification of group II NESp and increase the inclusion of NESp monitoring in future carriage studies [31].
While nasopharyngeal swabs remain the gold-standard sample for pneumococcal carriage detection [32], swab collection is invasive and resource-intensive, requiring specialized materials and trained healthcare professionals. Saliva is a more accessible sample type than swabs [33] and is increasingly being used for pneumococcal carriage detection [3439]. As such, the use of saliva for community-based studies continues to be validated and optimized. For example, we and others have demonstrated the high storage stability of respiratory pathogens, including encapsulated pneumococci, in unsupplemented saliva samples [4043]. However, the stability of NESp, particularly the potentially more virulent NCC1 and NCC2 strains, has not yet been similarly investigated. Therefore, we examined the stability of the detection of NESp in saliva, over different temperatures, timespans and freeze-thaw cycles, to better inform the experimental design of future saliva-based pneumococcal carriage studies and improve the accuracy of NESp carriage estimates.
Methods
Saliva samples
Participants were asked to provide whole-mouth unstimulated saliva by passively drooling saliva into a sterile 25 ml polypropylene tube at least 30 min after eating, drinking or brushing their teeth. Samples were transferred at room temperature to the laboratory within 30 min of collection for temporary storage at 4 °C, and aliquots were made and stored at −80 °C within 12 h.
Each sample was thoroughly screened for the absence of pneumococci by incubating an aliquot of each sample at 37 °C and tested at 24, 48 and 72 h for the genes encoding the major pneumococcal autolysin LytA (lytA) [44] and the pneumococcal iron uptake ABC transporter lipoprotein PiaB (piaB) [4546], using methods described previously [31]. Saliva samples which tested negative for both piaB and lytA were thawed, pooled, aliquoted and either placed on ice for immediate use or stored until needed at −80 °C.
Bacterial isolates
Pneumococcal isolates (Table 1) representing the major NCCs (NCC1, 2a and 2b) were obtained from Lance Keller and Larry McDaniel (University of Mississippi Medical Centre, USA), Moon H Nahm (University of Alabama, USA) and Ron Dagan (Ben-Gurion University, Israel) [294748].
Isolates were plated as a lawn onto tryptic soy agar II plus 5% (v/v) defibrinated sheep blood (blood plates) and incubated at 37 °C with 5% CO_2_ overnight. The lawn was harvested into 1 ml brain heart infusion (BHI) medium using a cotton swab and used to inoculate 45 ml BHI medium. Cultures were grown at 37 °C with 5% CO_2_ to an OD at 620 nm (OD_620_) of ~0.6 absorbance units. Cultures were harvested by centrifugation at 4,000 g, and pellets were resuspended in 5–10 ml BHI medium supplemented with 10% (v/v) glycerol and stored at −80 °C. The bacterial concentration (c.f.u. per millilitre) of each sample was determined by colony counting of serially diluted samples cultured on blood plates and incubated at 37 °C with 5% CO_2_ overnight. Preliminary work from this study (data not shown) and other studies has found that due to aggregate formation, vigorous vortexing is required between serial dilution of NESp [49]. The recommended approach, to dilute into individual 1.5 ml microcentrifuge tubes with vortexing for 5–10 s between dilutions, was adopted here to ensure accurate colony counting.
Experimental design
For each isolate listed in Table 1, concentrated stocks were diluted in BHI and then spiked into saliva at final concentrations of 1,000 and 10,000 c.f.u. ml^−1^ (final concentration of BHI<3% v/v), as described previously [40]. Spiked saliva samples were incubated at 4 °C, room temperature (~20 °C) and 30 °C. At 24 h, 48 h and 72 h, the samples were vortexed, and 100 µl was removed from each for culture enrichment (CE) (plated immediately, timepoint 0) to test for strain viability as described previously [3140]. A further 50 µl of each sample was aliquoted and stored at −80 °C for extraction-free pneumococcal detection [43].
To determine the effect of freeze-thawing on the stability of pneumococcal detection, saliva was spiked with pneumococci to final concentrations of 1,000 and 10,000 c.f.u. ml^−1^ as above (unsupplemented) or supplemented with BHI with 50% glycerol (final concentrations of 30% BHI v/v and 15% glycerol v/v). Aliquots were stored for a minimum of 3 h at either −20 °C or −80 °C to allow complete freezing, before being thawed at room temperature. Once thawed, the samples were vortexed, and 100 µl was removed from each sample for CE before the remainder was returned to −20 °C or −80 °C. This process was repeated twice more. Where sample volume was insufficient for processing due to technical issues, the datapoint was excluded.
To further examine the recovery of strains immediately following spiking, representative NCC1 (MNZ11) and NCC2 (MNZ41) strains were spiked at 15,000 c.f.u. ml^−1^ into five different lytA- and piaB-negative saliva samples collected and prepared as above. Each individual saliva sample was spiked in a matched pairs fashion. Stains were spiked into 1 ml BHI in the same way in triplicate. Following spiking, samples were vortexed for 10 s and then plated for CE within 10 min.
Saliva sample processing by CE-DNA extraction or extraction-free methods
Culture-enriched saliva samples were thawed at room temperature, and DNA was extracted from 200 µl of each sample using the MagMAX Viral/Pathogen Nucleic Acid Isolation Kit (MVP I) using a KingFisher Apex instrument (Thermo Fisher Scientific), with modifications [39]. The method is referred to here as CE-DNA extraction.
Spiked saliva samples were additionally processed using an extraction-free method, without CE, as described previously [43]. This method involves a lysis step with proteinase K, heat inactivation at 95 °C for 10 min and immediate transfer of the treated sample (which we will refer to as lysate) for pneumococcal detection via qPCR.
Detection of pneumococcal carriage
Each sample (2.5 µl of either DNA template or extraction-free lysate) was tested using the same dualplex qPCR targeting piaB and lytA as described above. Genomic DNA extracted from a pneumococcus serotype 19A strain (lytA- and piaB-positive) was included in every plate as a positive control, using at least two standard concentrations (0.0001–1 ng µl^−1^). Assays were run on a CFX96 Touch instrument (Bio-Rad) under the following conditions: 95 °C for 3 min followed by 45 cycles of 98 °C for 15 s and 60 °C for 30 s. Samples were considered positive for pneumococci when the quantification cycle (Cq) values for both genes were ≤40 and within 2 Cq of each other, and negative controls were undetectable [50]. Since NESp strains MNZ41, C144.66 and D48 are naturally piaB-negative, only lytA positivity was considered for samples spiked with these strains [31]. Plate-to-plate variation was corrected for by multiplying each sample Cq by an adjustment factor as described previously [31].
Statistical analyses
Linear regression using R (version 3.5.2) was conducted to evaluate the impacts of time and temperature (time) or freeze-thaw cycle and supplementation (freeze-thaw) on the detection of pneumococci from spiked saliva samples using the lytA-adjusted Cq. Interaction terms were used to evaluate whether the effects of time and temperature (time), freeze-thaw cycle and supplementation (freeze-thaw) and starting concentration (both) varied by strain. The ΔCq value represents the change in the mean Cq value from freshly spiked saliva under each condition (categorical). P values of less than 0.05 were considered significant. Tabulated model results are available in the Supplementary Material.
To further study viability following CE-DNA extraction of NCC1 vs NCC2 strains immediately after spiking into saliva or BHI, a mixed-effects analysis was employed using GraphPad Prism (version 10.3.0) with Tukey’s multiple comparisons, utilizing a restricted maximum likelihood algorithm due to differently sized groups. The experiment was conducted in a repeated measures fashion, and the ΔCq value represents the change in the mean value between each strain. P values of less than 0.05 were considered significant.
Results
Detection of culture-enriched NESp depends on storage conditions, supplementation and NCC
When tested following CE-DNA extraction, the stability of detection of NESp over time was dependent upon temperature (Fig. 1a). There was no significant loss in detection over 72 h when stored at 4 °C (ΔCq=1.01, P=0.083, 95% CI: −0.14 to 2.16). However, detection dropped significantly after 48 h when stored at RT (ΔCq=3.44, P<0.001, 95% CI: 1.50–5.38), and after 24 h when stored at 30 °C (ΔCq=4.86, P<0.001, 95% CI: 2.76–6.97). In addition, the stability of detection over time and temperature depended on NCC. When stratified by NCC, detection of NCC2 strains, but not NCC1, dropped significantly after 72 h when stored at 4 °C (ΔCq=1.68, P=0.035, 95% CI: 0.12–3.24), after 48 h when stored at RT (ΔCq=3.92, P=0.006, 95% CI: 1.20–6.64), and for less than 24 h when stored at 30 °C (ΔCq=7.00, P<0.001, 95% CI: 4.38–9.60).
Detection of NCC1 NESp over time, temperature and freeze-thaw is more stable than that of NCC2 when spiked into saliva. Panels (a) and (b) show the detection of NESp over time at different temperatures (4 °C, RT=room temperature and 30 °C) following either culture enrichment and DNA extraction (CE+DNA) or extraction-free methods, respectively. Pooled saliva was initially spiked at either 1,000 or 10,000 c.f.u. ml−1, as indicated by the right-hand label. Panels (c) and (d) show the detection of NESp over three freeze-thaw cycles following either CE+DNA or extraction-free methods, respectively. Pooled saliva was initially spiked into unsupplemented saliva or saliva supplemented with BHI plus glycerol at either 1,000 or 10,000 c.f.u. ml−1 and cycled through either −20 °C or −80 °C, as indicated by the right-hand label. Data are shown as the mean adjusted Cq for lytA for each of NCC1 (n=2, red) or NCC2 (n=4, blue) with the range shown as a shaded ribbon.
We next assessed the effect of freeze-thaw cycles on the detection stability of NESp (Fig. 1c). For samples that were processed by CE-DNA extraction, detection stability of NESp across freeze-thaw cycles was dependent on storage temperature. There was a significant reduction in detection by the second freeze-thaw cycle for samples stored at −20 °C (ΔCq=4.46, P<0.001, 95% CI: 2.25–6.68) and by the third cycle for samples stored at −80 °C (ΔCq=2.34, P=0.008, 95% CI: 2.25–6.68), suggesting a slightly greater freeze-thaw stability when stored at −80 °C.
When tested using CE-DNA extraction, adding BHI+glycerol to the saliva significantly increased overall bacterial detection following freeze-thaws at −80 °C (ΔCq unsupplemented vs glycerol=3.79, P<0.001, 95% CI: 2.58–5.00) and −20 °C (ΔCq unsupplemented vs glycerol=5.22, P<0.001, 95% CI: 3.65–6.78); however, glycerol supplementation did not affect the stability of detection through consecutive freeze-thaws (ΔCq 0 h vs 72 h=1.62, P=0.251, 95% CI: −1.16 to 4.41) (Fig. 1c). Similarly, while overall detection of NCC2 was lower than that of NCC1 strains (ΔCq=4.20, P<0.001, 95% CI: 3.17–5.25), stability of detection across freeze-thaw cycles was unaffected by NCC (ΔCq=0.42, P=0.781, 95% CI: −2.56 to 3.40).
Processing of NESp using an extraction-free method leads to greater stability of detection which is less dependent on storage conditions and NCC
To better understand the impact of culture enrichment on the detection of NESp strains, spiked saliva samples which underwent the same storage conditions were also tested using the extraction-free method, which does not include CE. Similarly to when tested using CE-DNA extraction, detection was stable over 72 h when stored at 4 °C (ΔCq=0.50, P=0.127, 95% CI: −0.15 to 1.15). However, when stored at room temperature, there was a slight increase in overall detection at 24 h (ΔCq=−1.14, P=0.017, 95% CI: −2.07 to −0.21), which became a decrease in detection by 72 h (ΔCq=1.24, P=0.10, 95% CI: 0.31–2.16), suggesting that some strains may have grown a small amount in the saliva. At 30 °C, overall detection dropped after 48 h (ΔCq=4.91, P<0.001, 95% CI: 2.86–6.97).
Unlike with CE-DNA extraction, there was no difference in stability of detection of NCC at either 4 °C or room temperature when processed using the extraction-free method (Fig. 1b). Detection according to NCC only deviated when stored at 30 °C, in which detection of NCC2 strains decreased significantly after 24 h (ΔCq=2.88, P<0.027, 95% CI: 0.35–5.40), whereas detection of NCC1 strains, like at room temperature, increased at 24 h (ΔCq=−2.93, P<0.002, 95% CI: −4.55 to 1.30), before decreasing by 72 h (ΔCq=1.92, P<0.025, 95% CI: 0.29–3.54). This suggests that NCC1 strains, and less so NCC2 strains, can grow in whole saliva at temperatures between 20 and 30 °C.
Detection of NESp remained stable over all three freeze-thaw cycles when processed using the extraction free method (ΔCq=0.47, P=0.058, 95% CI: −0.02 to 0.95), and we did not observe any impact on NESp detection over repeated freeze-thaw cycle by NCC strain (ΔCq=−0.14, P=0.470, 95% CI: −0.51 to 0.23), or storage temperature (ΔCq=−0.12, P=0.507, 95% CI: −0.23 to 0.47) (Fig. 1d). Unexpectedly, addition of glycerol supplementation resulted in slightly lower overall detection when samples were processed using the extraction-free method (ΔCq unsupplemented vs supplemented=−2.22, P<0.001, 95% CI: 2.57–1.87). Glycerol is typically considered a qPCR enhancer and cryoprotectant [51]; however, it is possible that the addition of glycerol in this concentration or formulation reduced the efficiency of the proteinase K or heat lysis step, resulting in less DNA available for qPCR.
Overall, there were twice as many samples in which lytA was not detectable at the endpoint following CE-DNA extraction (7.4%) than extraction-free processing (3.4%) (Table 2). Almost all the samples in which lytA could not be detected (32 out of 663) were originally spiked with an NCC2 strain, except for four unsupplemented saliva samples which were spiked with an NCC1 strain and underwent ≥2 freeze-thaw cycles.
Following the observation that NCC2 strains were less stable in saliva than NCC1 strains following CE, we sought to determine if this was driven by the presence of aliC and aliD. Strains belonging to NCC1 (MNZ11) and NCC2 (MNZ41) were spiked into saliva and BHI in parallel and immediately tested using the CE-DNA extraction method. Recovery of viable NCC1 strains was significantly higher than NCC2 following spiking into saliva (mean ΔCq=−3.52, 95% CI: −4.67 to −2.37, P<0.0001; Fig. 2a). However, there was no significant difference when spiked into BHI (mean ΔCq=−0.06, 95% CI: −1.19 to 1.08, P=0.9972; Fig. 2b). Recovery of both strains was lower and more varied in saliva than in BHI, suggesting that a component of saliva has an impact on bacterial viability during CE, which was more pronounced for NCC2 strains.
*Recovery of NCC2 NESp is lower than NCC1 immediately after spiking into saliva, but not BHI, and this is not dependent on aliC and aliD, or pspK. MNZ11 (red, filled circles) is NCC1, MNZ41 (blue, filled circles) is NCC2 and MNZ1131 (red, open circles) and JLB01 (blue, open circles) are pspK and aliC/aliD gene knockouts of MNZ11 and MNZ41, respectively. Strains were spiked at a final concentration of 15,000 c.f.u. ml−1 into either BHI (n=6 for MNZ11 and MNZ41, n=3 for MNZ1131 and JLB01) or matched pairs of lytA- and piaB-negative saliva from healthy individuals (n=11 for MNZ11 and MNZ41, n=6 for MNZ1131 and JLB01). Line is the mean, and significance is shown for multiple comparisons between MNZ11 vs MNZ41, MNZ11 vs MNZ1131 and MNZ41 vs JLB01, **P<0.005, ***P<0.0005, ***P<0.0001.
Knockout mutants MNZ1131 (ΔpspK) and JLB01 (ΔaliCaliD) were also tested, and while recovery in BHI was significantly lower for both when compared to their wild types (Fig. 2b), there were no significant differences in recovery from saliva between the respective wild types and mutants. This suggests that while pspK, aliC and aliD do play a role in overall cell fitness, they are not primarily driving the effect on viability of NESp during CE in saliva.
Discussion
Carriage studies are reporting general increases in the prevalence of NESp, which is of concern, particularly in the case of NCC1 and NCC2 strains, due to the increased virulence of these strains compared with other NESp, and their propensity to acquire AMR genes [57131516]. This is particularly true for pspK+ strains, which have some of the highest frequencies of genetic exchange among pneumococci [17]. There has also been an increase in recent years in the use of saliva as a sample type for investigating pneumococcal carriage; however, the impact of saliva sample storage on the detection and recovery of NESp has not yet been described [34,36, 38, 45, 5254].
Therefore, in this study, we sought to build upon the existing evidence examining encapsulated pneumococci viability in saliva across a range of storage conditions and determine if similar findings applied to NESp. We previously demonstrated that the viability of pneumococci remained stable in self-collected, unsupplemented saliva samples stored between 4 and 30 °C for up to 24 h, and a portion of viable pneumococci could be detected for up to 72 h, though at lower quantities [40]. Findings from this current study demonstrate that while the NCC1 strains tested here exhibited equivalent storage stability in saliva to encapsulated strains [40], the NCC2 strains tested were less able to survive CE, leading to lower detection than NCC1 following CE-DNA extraction (Figs 1a, c and 2a).
Overall, NESp detection using an extraction-free method was generally lower compared with detection following CE-DNA extraction. This finding was expected, due to the lack of enrichment and DNA concentration steps in the extraction-free method. Interestingly, though, compared with CE-DNA extraction, the extraction-free method yielded more stable detection of NESp over both time (Fig. 1b) and freeze-thaw cycles (Fig. 1d), with negligible variation between NCC1 and NCC2. There were also fewer samples with a total loss of detection at the endpoint when using the extraction-free method compared with CE-DNA extraction (3.4% vs 7.4%, Table 2).
CE amplifies all viable gentamicin-resistant strains in a sample, the totality of which is harvested for detection [3445]. Thus, for pneumococcal detection using CE, any pneumococcal strains present must be both viable at the time of plating and able to survive on the culture plate during incubation. In the case of saliva, this method enriches for pneumococcus alongside other gentamicin-resistant mitis-group streptococci, which all compete for resources and space [55]. Meanwhile, the extraction-free method used in this study consists simply of heat and enzymatic lysis step and direct qPCR, meaning that bacterial viability is not a requirement for detection.
Therefore, we hypothesized that the observed reduction in NCC2 strain detection following the CE-DNA extraction method was due to reduced cell survival in saliva and/or growth during CE. To test this, we compared the recovery of cells immediately after spiking into either saliva or BHI and found that while there was no difference in NESp recovery between strains when culture-enriched from BHI, recovery of NCC2 strains was significantly lower than that of NCC1 when culture-enriched from saliva (Fig. 2). These findings suggest that while all cells were viable at the time of plating, the NCC2 strains, but not NCC1, had a growth disadvantage specifically in saliva during culture enrichment. Potential explanations for this include a lower competitive fitness of these NCC2 strains against the other gentamicin-resistant bacteria present in saliva. Alternatively, these NCC2 strains may have been more susceptible to growth inhibition by a component of saliva, such as immune cells or proteases.
Previous studies have shown that aliC and aliD in NCC2 strains can compensate for the lack of capsule and aid survival in blood, by binding IgA, as well as by upregulating cbpAC, which increases resistance to complement deposition and neutrophil killing [24]. Additionally, these oligopeptide transporters bind peptides from other species, which can enhance carbohydrate metabolism, thus growth and colonization [56]. The presence of PspK has been shown to be necessary for NCC1 nasopharyngeal colonization in mice with a suggested role in cell adhesion similar to that of pspC [48]. Based on these findings, we hypothesized that pspK or aliC and aliD would also have compensatory roles in bacterial growth or survival in saliva. We found that aliCaliD and pspK knockouts both showed reduced viability compared to the wild types when spiked into BHI and tested using CE-DNA extraction. However, there were no significant differences between each mutant and wild-type strains when spiked into saliva (Fig. 2), suggesting that under these conditions and in these strains, pspK and aliC/aliD do not directly impact the growth or survival in saliva, and that this is determined by a more complex set of interactions.
The lack of capsule itself likely leads to reduced fitness compared with encapsulated strains, which, unlike the NCC1 strains, cannot be compensated for by NCC2 strains. A study of 57 NCC2 strains found all strains tested lacked several genes, including virulence factors pilus islet-1, cbpA, pspA, nanE, glf, ntpK and rlrA [57]. Further studies support these findings of major alterations in NCC2 genomes [19], beyond just changes in the cps locus, which may explain their reduced fitness during saliva-CE [8]. Further studies are needed which test the recovery of NCC1 and NCC2 strains from either heat-treated, filtered or fractionated saliva, to systematically determine the salivary component driving this apparent reduced NCC2 fitness during CE.
This study examined stability at two different bacterial loads, 1,000 and 10,000 c.f.u. ml^−1^, and the observed stability trends were generally replicated between each, with the expected ~3 Cq difference (Fig. 1). This suggests that the detection and quantification methods employed here were robust. Moreover, the recovery of representative NCC1 and NCC2 strains spiked into BHI was both highly replicable and similar between strains (Fig. 2), suggesting that the observed variations between strains could not be explained by variations in the initial quantity or viability of bacteria spiked into saliva. However, this study was limited by testing the stability of only a small number of NESp strains, and only in the group II, NCC1 and NCC2 clades. Moreover, we could not test any NCC2 strains from the MLST groups ST344 and ST448, which are predominant in conjunctivitis outbreaks [7]. Given the inter-strain differences observed here, it would be interesting to assess both additional group II NESp strains, as well as group I NESp strains. Gaining a comprehensive knowledge of the impact of capsule absence on pneumococcal viability is essential for informing future carriage study sample collection and storage practices.
NESp prevalence is likely underreported in carriage surveillance due to the limited focus on detection and characterization of these strains. We recently sought to address this by developing a method to aid the detection of NESp, particularly in oral sample types [31]. To further address the knowledge gap around NESp carriage, this current study aimed to determine the conditions for saliva collection, storage and testing for optimal NESp detection. As such, we have demonstrated that NESp can be reliably detected in saliva for up to 72 h when stored at 4 °C, or 24 h when stored at room temperature. We have also demonstrated that while glycerol supplementation of saliva slightly improves NESp detection when using CE, it is not otherwise necessary. We also showed that NCC1 strains tested here exhibited comparable resilience to encapsulated pneumococci, while NCC2 strains tested here demonstrated a competitive disadvantage during CE. Therefore, while CE may enhance detection of encapsulated pneumococci and NCC1 NESp, it can cause undue suppression of NCC2 NESp detection and possibly contribute to underreporting of NESp prevalence. Given that the extraction-free method is cheaper and less labour-intensive than CE-DNA extraction [43], it may be advisable to employ this, or other culture-free methods to ensure accurate detection of NESp. Further investigation is required to understand the mechanism underlying our observation of reduced NCC2 fitness in saliva during CE. These findings underscore the importance of refining detection methodologies and considering strain-specific differences when designing future pneumococcal carriage studies.
Supplementary material
10.1099/mic.0.001585Uncited Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Manna S Ortika BD Werren JP Pell CL Gjuroski I et al Streptococcus pneumoniae serotype 33H: a novel serotype with frameshift mutations in the acetyltransferase gene wci G Pneumonia 202517710.1186/s 41479-025-00162-240128891 PMC 11934437 · doi ↗ · pubmed ↗
- 2Blacklock CB Weinberger DM Perniciaro S Wyllie AL Streptococcus pneumoniae serotypes 2025 https://pneumococcalcapsules.github.io/serotypes/accessed 3-April-2025
- 3Lo SW Gladstone RA van Tonder AJ Lees JA du Plessis M et al Pneumococcal lineages associated with serotype replacement and antibiotic resistance in childhood invasive pneumococcal disease in the post-PCV 13 era: an international whole-genome sequencing study Lancet Infect Dis 20191975976910.1016/S 1473-3099(19)30297-X 31196809 PMC 7641901 · doi ↗ · pubmed ↗
- 4Waight PA Andrews NJ Ladhani SN Sheppard CL Slack MPE et al Effect of the 13-valent pneumococcal conjugate vaccine on invasive pneumococcal disease in England and Wales 4 years after its introduction: an observational cohort study Lancet Infect Dis 20151553554310.1016/S 1473-3099(15)70044-725801458 · doi ↗ · pubmed ↗
- 5Nunes S Félix S Valente C Simões AS Tavares DA et al The impact of private use of PCV 7 in 2009 and 2010 on serotypes and antimicrobial resistance of Streptococcus pneumoniae carried by young children in Portugal: Comparison with data obtained since 1996 generating a 15-year study prior to PCV 13 introduction Vaccine 2016341648165610.1016/j.vaccine.2016.02.04526920470 · doi ↗ · pubmed ↗
- 6Sá-Leão R Nunes S Brito-AvôA Frazão N Simões AS et al Changes in pneumococcal serotypes and antibiotypes carried by vaccinated and unvaccinated day-care centre attendees in Portugal, a country with widespread use of the seven-valent pneumococcal conjugate vaccine Clin Microbiol Infect 2009151002100710.1111/j.1469-0691.2009.02775.x 19392883 · doi ↗ · pubmed ↗
- 7Keller LE Robinson DA Mc Daniel LS Nonencapsulated Streptococcus pneumoniae: emergence and pathogenesism Bio 20167 e 01792 Epub ahead of print 22 March 10.1128/m Bio.01792-1527006456 PMC 4807366 · doi ↗ · pubmed ↗
- 8Iranzadeh A Alisoltani A Kiran AM Breiman RF Chaguza C et al Comparative pangenomics of Streptococcus pneumoniae from Malawi: uncovering genetic variability and pathogenicity Microb Genom 20251100137010.1099/mgen.0.00137040232949 PMC 12282314 · doi ↗ · pubmed ↗