The cystic fibrosis pathogen Achromobacter xylosoxidans inhibits biofilm formation of Pseudomonas aeruginosa
Cecilia Sahl, Agnes Andersson, Natalie Larsson, Magnus Paulsson, Oonagh Shannon, Lisa I. Påhlman

TL;DR
Achromobacter xylosoxidans can prevent Pseudomonas aeruginosa from forming biofilms in cystic fibrosis, which may impact lung health.
Contribution
This study reveals that A. xylosoxidans inhibits P. aeruginosa biofilm formation through direct live-cell interactions during early stages.
Findings
Early-stage A. xylosoxidans isolates inhibit P. aeruginosa biofilm formation in a concentration-dependent manner.
The inhibition is not due to nutrient depletion but requires live-cell interactions during the attachment phase.
A. xylosoxidans adheres more strongly to surfaces than P. aeruginosa.
Abstract
Background. Achromobacter xylosoxidans and Pseudomonas aeruginosa are two pathogens that cause persistent airway infections in individuals with cystic fibrosis (CF). The persistence of P. aeruginosa is partly due to a high capacity to form biofilms and the ability to exert antagonism against other bacteria. Loss of microbial diversity in conjunction with chronic P. aeruginosa colonization is strongly correlated with low lung function in CF. A. xylosoxidans and P. aeruginosa are frequently co-isolated in CF airway cultures. This study aims to investigate the reciprocal effects on growth inhibition and biofilm formation between P. aeruginosa and A. xylosoxidans in vitro. Method. Six isolates of A. xylosoxidans, isolated from three CF patients in early and late stages of a chronic infection, were cultured together with a CF isolate of P. aeruginosa. Biofilm formation was assessed using a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
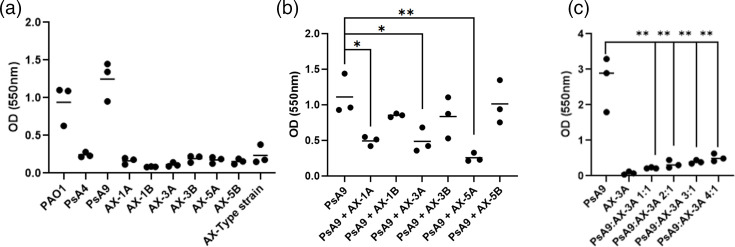 Fig. 1
Fig. 1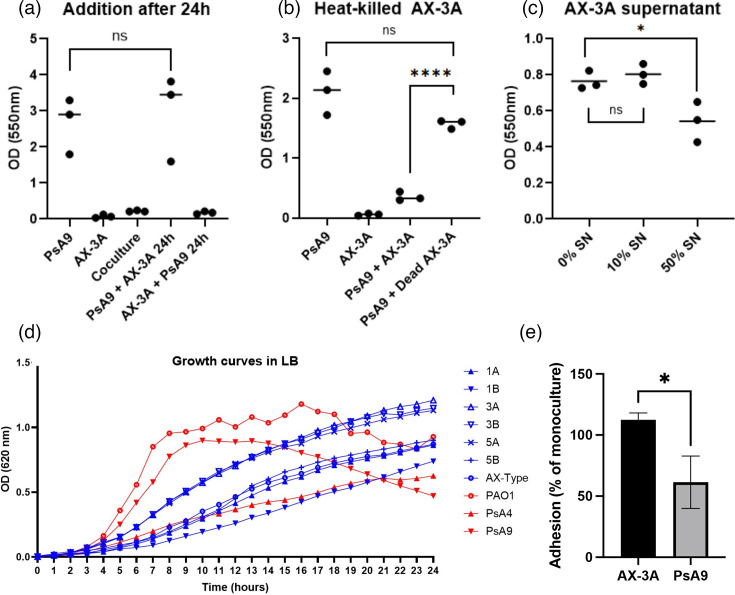 Fig. 2
Fig. 2| Isolate name | Species | Origin |
|---|---|---|
| PAO1 |
| Reference strain |
| PsA4 |
| CF sputum, Lund, Sweden |
| PsA9 |
| CF sputum, Lund, Sweden |
| KM543 |
| Type strain, ear infection, Japan |
| AX-1A |
| CF sputum, Lund, Sweden |
| AX-1B |
| CF sputum, Lund, Sweden |
| AX-3A |
| CF sputum, Lund, Sweden |
| AX-3B |
| CF sputum, Lund, Sweden |
| AX-5A |
| CF sputum, Lund, Sweden |
| AX-5B |
| CF sputum, Lund, Sweden |
| Patient | Early | Colonization at the time of early isolation | Interval (years) between early and late isolates | Late | Colonization at the time of late isolation |
|---|---|---|---|---|---|
| CF 1 | AX-1A |
| 6.8 | AX-1B |
|
| CF 3 | AX-3A |
| 3.6 | AX-3B |
|
| CF 5 | AX-5A |
| 7.2 | AX-5B |
|
- —Knut and Alice Wallenberg Foundation, medical faculty at Lund University and Region Skåne
- —Swedish government funds for clinical research
- —http://dx.doi.org/10.13039/501100004359 Vetenskapsrådet
- —http://dx.doi.org/10.13039/501100009235 Stiftelsen Riksförbundet Cystisk Fibros Forskningsfond
- —http://dx.doi.org/10.13039/501100005390 Alfred Österlunds Stiftelse
- —http://dx.doi.org/10.13039/501100003793 Hjärt-Lungfonden
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial biofilms and quorum sensing · Infections and bacterial resistance · Microbial Community Ecology and Physiology
Introduction
The microbiome composition of the cystic fibrosis (CF) airway correlates with the severity of lung disease, where a loss of microbial diversity and colonisation with certain bacterial species, including Pseudomonas aeruginosa, often indicates more severe outcomes [13]. The cystic fibrosis transmembrane regulator (CFTR) mutation in CF causes an accumulation of viscous mucus in the airways, which constitutes a permissive environment for bacterial and fungal growth. The infections lead to progressive airway damage due to primarily neutrophilic inflammation [4].
Biofilm formation is one of the main causes of persistent P. aeruginosa infections in people with CF [5]. By embedding within a matrix of polysaccharides, nucleic acids and proteins, bacteria can evade host immune responses and antibiotics. Once a chronic P. aeruginosa infection has been established, antibiotic treatment is used to reduce the bacterial load but is rarely sufficient to eradicate the pathogen [6], even after treatment with CFTR modulators that reduce mucus production. In addition to resisting host immune defences and antibiotic treatments, the capacity of P. aeruginosa biofilms to persist in the CF airway is also affected by its interactions with other species in the microbiome. It has been previously shown that P. aeruginosa employs multiple antagonistic mechanisms towards Staphylococcus aureus, the most common CF infection in early life, to gain a competitive advantage [7].
Achromobacter xylosoxidans is a Gram-negative bacterial species that is associated with decreased lung function and more frequent requirement for intravenous antibiotics and hospitalizations in people with CF (pwCF) [810]. Similarly to P. aeruginosa, these infections are difficult to eradicate once established and may persist for many years. In the respiratory tract, A. xylosoxidans undergoes adaptation and in-host evolution to increase its fitness in the niche [1112]. Previous studies show that both P. aeruginosa and A. xylosoxidans are independently associated with increased inflammatory markers [1314] and low lung function [15], but patients infected with both species have the worst outcomes [16]. In a large-scale screening study of interactions between the four CF pathogens P. aeruginosa, Staphylococcus aureus, A. xylosoxidans and Stenotrophomonas maltophilia, Menetrey et al. [17] show that only A. xylosoxidans was able to negatively impact the growth of P. aeruginosa.
In this study, we investigate interactions between clinical isolates of P. aeruginosa and longitudinal pairs of A. xylosoxidans isolated from the sputum of pwCF. The pathogens were grown in mono- and co-cultures to study both planktonic growth and biofilm formation with the aim of investigating competition between these pathogens throughout chronic infections and to determine the mechanisms involved.
Methods
Bacterial isolate selection
The isolates included in this study are presented in Table 1. Clinical isolates of A. xylosoxidans and P. aeruginosa from CF sputum cultures were obtained from the Clinical Microbiology laboratory, Skåne University Hospital, Lund, Sweden. Species identification was performed according to standard laboratory methods, including matrix-assisted laser desorption/ionization-time of flight MS. Species identification of A. xylosoxidans was further confirmed using whole-genome sequencing [11]. The A. xylosoxidans type strain KM543 was purchased from the Culture Collection University of Gothenburg. The P. aeruginosa reference strain PAO1 was a kind gift from Professor Arne Egesten, Lund University. All isolates were grown in Luria–Bertani (LB) broth (Sigma-Aldrich).
Biofilm formation and biomass quantification
Overnight cultures of each strain were normalized to OD_620_=0.6, diluted 1:10 in LB (18+162 µl) and grown in a 96-well plate. Co-cultures were prepared by mixing monocultures of P. aeruginosa and A. xylosoxidans in varying proportions (1:1, 2:1, 3:1 or 4:1), and 18 µl of the mixed suspensions were added to 162 µl LB in a 96-well plate. The plates were incubated for 72 h at 37 ℃ with 5% CO_2_ and 150 r.p.m. Wells were washed three times with PBS by pipetting to remove planktonic bacteria, and washed biofilms were then fixated with 200 µl of methanol for 10 min. The methanol was removed, and the plate was dried at room temperature for 3 h. 0.1% crystal violet was added in a volume of 200 µl, and the plate was incubated for 5 min. After washing the wells three times with PBS, the remaining stained biofilm was dissolved with 20:80 acetone–ethanol and diluted 1:3 before absorbance readout at 550 nm. Experiments were performed in biological and technical triplicate.
In separate experiments, PsA9 and AX-3A were grown in monocultures as described earlier. After 24 h of incubation, 18 µl of AX-3A was added to PsA9, 18 µl of PsA9 was added to AX-3A and bacteria were co-cultured for another 48 h. Alternatively, PsA9 was co-cultured 1:1 with AX-3A that had been heat-killed by incubation at 95 ℃ for 5 min or in the presence of 10% or 50% of cell-free AX-3A growth medium supernatant. The supernatants were prepared by growing AX-3A overnight in LB, pelleting the cells by centrifugation, followed by sterile filtering of the cell-free supernatants through a 0.22-µm syringe filter. After incubation for 72 h, biofilm mass was quantified using crystal violet staining as described earlier.
Bacterial DNA quantification from planktonic bacteria and biofilms
Overnight cultures were centrifuged at 4,000 g for 5 min and resuspended to a concentration of ~2×10^9^ c.f.u. ml^−1^. Fifty microlitres of this suspension were added to 5 ml LB broth or 50+50 µl in co-culture. Cells were grown until reaching the stationary phase, ~24 h. 1.8 ml of the resulting culture was collected for homogenization using BeadBug (Benchmark Scientific), followed by DNA extraction using the DNeasy PowerLyzer Microbial Kit (QIAGEN) according to the manufacturer’s instructions.
Mono- or co-cultured biofilms, grown and washed with PBS as described earlier, were collected in 100 µl of PowerBead solution by scraping the wells with the pipette tip to dislodge the biofilms. Triplicate wells were pooled into one PowerBead tube before proceeding with homogenization and DNA extraction as described earlier. Extracted bacterial DNA was quantified with quantitative PCR (qPCR) using iTaq Universal SYBRGreen Supermix (Bio-Rad) using specific primers targeting Achromobacter spp.; AcForward: CACTAGCTCACGAACTCCAAGC, AcReverse: CAGCTTCAATCCTACCTAACTTTCCT [18] and P. aeruginosa rpoS; forward: TTGAGATACAGCTGCGTTGC, reverse: CTCCAAAAGCCACCACTTCC. Known concentrations of DNA from A. xylosoxidans or P. aeruginosa in tenfold dilutions were used as standards.
Growth curves
Isolates were cultured overnight, diluted to an OD_620_ of 0.1 and added at a volume of 15 to 150 µl medium in a 96-well plate. The plate was incubated at 37 °C with 150 r.p.m. shaking and OD_620_ measurements were performed once every hour for 24 h using a SpectraMax M2 plate reader (Molecular Devices). In addition, growth rates were assessed in the presence of 10% or 50% bacterial cell-free supernatants. Growth rate experiments with supernatants were performed in 10-ml tubes containing 125 µl overnight culture (OD_620_ 0.1), 500 µl (10%) or 2,500 µl (50%) sterile-filtered supernatant, and fresh LB at a total volume of 5 ml.
Adhesion
Overnight cultures of each isolate were normalized to an OD_620_ of 0.1. In total, 1 ml of bacterial culture (1 ml of monoculture or 0.5 ml of each species in coculture) was added to 12 well-plates. After incubation at 37 °C with shaking for 1.5 h, the growth medium containing non-adherent bacteria was removed, and the wells were washed once with PBS. DNA was extracted by the addition of 300 µl PowerBead solution (Qiagen DNeasy PowerLyser Microbial Kit, QIAGEN) to each well, and bacteria were detached with a cell scraper. Samples were transferred to PowerBead Tubes for homogenization and subsequent DNA extraction according to the protocol provided by the manufacturer. A. xylosoxidans and P. aeruginosa DNA were quantified using qPCR as described earlier.
Results
Evaluation of biofilm formation in P. aeruginosa–A. xylosoxidans cocultures
Biofilm formation is an important virulence mechanism for the establishment of chronic infections in the CF airway. We first screened two clinical P. aeruginosa and six clinical A. xylosoxidans CF isolates for biofilm formation using a crystal violet assay. In addition, type strains of P. aeruginosa and A. xylosoxidans were included as a reference. The clinical A. xylosoxidans isolates were longitudinal pairs obtained from the same patient at an early and a late stage of chronic infection (Table 2). Isolates were selected based on the P. aeruginosa colonization status of the corresponding patients. The first pair (1A-1B) was derived from a patient who was not infected with P. aeruginosa. The second pair (3A-3B) originated from a patient who originally had a persistent P. aeruginosa infection that was subsequently cleared during A. xylosoxidans infection. The third pair (5A-5B) was isolated from a patient with persistent P. aeruginosa infection throughout both the early and late stages of A. xylosoxidans infection. In the initial screening of biofilm formation, the P. aeruginosa isolates PAO1 and PsA9 were strong biofilm producers, whereas PsA4 and all A. xylosoxidans isolates tested were poor biofilm producers (Fig. 1a). To further investigate biofilm formation in dual-species biofilms of P. aeruginosa and A. xylosoxidans, we selected the strong biofilm-producing isolate PsA9 to study the effects of the different A. xylosoxidans isolates on biofilm formation. When PsA9 was co-cultured with the early isolates AX-1A, AX-3A and AX-5A, a significant reduction of Pseudomonas biofilm mass was observed compared to PsA9 grown in monoculture. In contrast, no biofilm reduction was seen when PsA9 was co-cultured with the late isolates AX-1B, AX-3B and AX-5B (Fig. 1b). To further characterize biofilm production in co-cultures, we focused on the interaction between AX-3A and PsA9 since this isolate had out-persisted P. aeruginosa during clinical infection (Table 2). First, we added AX-3A in varying proportions to investigate whether the observed effect was concentration-dependent (Fig. 1c). The biofilm-inhibiting effect decreased with lower concentrations of AX-3A. However, a significant inhibition of biofilm formation was still observed at a P. aeruginosa vs. AX-3A ratio of 4:1 (Fig. 1c).
*Biofilm formation of CF pathogens in mono- and co-culture. (a) Crystal violet quantification of biofilm formation of P. aeruginosa and A. xylosoxidans isolates in monoculture. (b) Biofilm formation of P. aeruginosa isolate PsA9 grown in monoculture or together with three pairs of early (AX-1A, AX-3A and AX-5A) and late (AX-1B, AX-3B and AX-5B) A. xylosoxidans isolates. (c) PsA9 was co-cultured with increasing proportions of A. xylosoxidans AX-3A, followed by quantification of biofilm formation. Each dot in the graphs represents the average value of 3 replicates, and bars represent the mean. *P<0.05, *P<0.01.
Biofilm inhibition by A. xylosoxidans is caused by interactions between live bacteria during the early stages of biofilm formation
Quantification of PsA9 and AX-3A DNA in co-culture biofilms revealed that the biomass of the dual-species biofilms was dominated by P. aeruginosa, but in lowered quantity compared to in parallel monocultures (Fig. S1A, available in the online Supplementary Material). A similar domination by P. aeruginosa was seen in dual biofilms with PsA9 and AX-3B (Fig. S1A). To study the mechanisms of competition by A. xylosoxidans, we next established a biofilm by culturing PsA9 or AX-3A for 24 h, after which planktonic AX-3A or PsA9 was added for the remaining 48 h of incubation. Adding A. xylosoxidans to an existing P. aeruginosa biofilm had no inhibitory effect on the biofilm mass. In contrast, PsA9 could not establish a biofilm in wells already colonized by AX-3A (Fig. 2a), indicating that the inhibitory interaction takes place during the establishment phase and not as a disruption of the P. aeruginosa biofilm. Next, we determined that heat-killed AX-3A did not affect biofilm production of PsA9 (Fig. 2b), indicating that the interaction requires live bacteria. Adding secreted Achromobacter products by incubating PsA9 together with 50% sterile-filtered supernatant of AX-3A moderately lowered the biofilm production (Fig. 2c), but not as strongly as live bacteria.
*Mechanisms of interaction between P. aeruginosa and A. xylosoxidans during biofilm formation. (a–c) PsA9 and AX-3A were grown in dual- or single-species biofilms in 96-well microtiter plates for 48 h. (a) Addition of AX-3A to an established PsA9 biofilm after 24 h does not affect biofilm formation, and PsA9 cannot establish a biofilm in wells already colonized by AX-3A. (b) Co-culturing of live PsA9 with heat-killed AX-3A. (c) PsA9 was allowed to form biofilm in the presence of 0%, 10% or 50% of cell-free culture supernatant from AX-3A. (d) Growth curves of seven A. xylosoxidans and three P. aeruginosa isolates grown as planktonic monocultures in 96-well microtiter plates during 24 h with continuous hourly OD measurement. (e) Comparison of bacterial adhesion to microtiter plates. Equal volumes of PsA9 and AX-3A were incubated in microtiter plates for 90 min to allow bacterial adhesion to the wells. Monocultures were incubated in parallel for comparison. After washing steps, attached bacteria were detached, and A. xylosoxidans and P. aeruginosa DNA were quantified using qPCR. The graph shows bacterial DNA from adhered bacteria in co-cultures compared to monocultures in three replicates. All statistics are unpaired t-tests, *P≤0.05, ***P≤0.0001, ns, not significant.
Biofilm inhibition is not caused by differential growth rates
To investigate whether biofilm inhibition was caused by A. xylosoxidans outcompeting P. aeruginosa in growth kinetics, we analysed the planktonic growth rates of the bacterial strains included in the study (Fig. 2d). PsA9 was the most rapidly growing strain with the steepest slope during the exponential phase. None of the A. xylosoxidans strains displayed rapid growth during the exponential phase, indicating that biofilm inhibition was not caused by A. xylosoxidans outgrowing P. aeruginosa. To confirm that the observed inhibitory effect was not predominantly caused by nutrient depletion, we performed growth curves of PsA9 grown in 10% and 50% AX-3A supernatant. PsA9 did not exhibit growth inhibition in the presence of either concentration of AX-3A supernatant (Fig. S1B). Finally, we performed DNA quantification of broth co-cultures to investigate whether inhibitory effects also occur in planktonic cultures. After growth in liquid culture media until the stationary phase, the final DNA concentration of planktonic PsA9 was not lowered by the presence of AX-3A (Fig. S1C), indicating that the reduced biofilm formation observed in biofilm co-cultures was not caused by growth inhibition of PsA9.
AX-3A exhibits stronger adhesion ability than PsA9
As planktonic A. xylosoxidans cultures did not grow faster than PsA9, we speculated that the interaction causing biofilm inhibition takes place at the early adhesion stage. To investigate adhesion ability, PsA9 and AX-3A/AX-3B in mono- or co-cultures were allowed to adhere to microtiter wells for 90 min. After washing, attached cells were detached, and bacterial DNA was quantified to assess the bacterial load. All three isolates adhered to a similar extent when incubated in monocultures (Fig. S2A). By comparing DNA concentrations from co-cultures versus monocultures, we found that a significantly lower proportion of PsA9 DNA was retrieved from co-cultures compared to AX-3A, suggesting a lower adhesion rate for PsA9 (Fig. 2e). In contrast, no significant difference was observed between PsA9 and AX-3B when grown in co-culture (Fig. S2B). Taken together, the data suggest that AX-3A negatively impacted PsA9 biofilm formation due to a higher adherence rate to the plastic of the microtitre well.
Discussion
In this study, we report that A. xylosoxidans is capable of inhibiting biofilm formation by P. aeruginosa. We demonstrate that the interaction takes place during the early stages of biofilm development and is likely related to how efficiently the strain adheres to a surface. These findings are consistent with previous reports showing that approximately half of A. xylosoxidans strains possess this inhibitory capacity against P. aeruginosa biofilms [19]. Notably, all three isolates causing biofilm inhibition in our study were obtained from early-stage infections, whereas the corresponding late-stage isolates from the same patient lacked this ability. This observation aligns with earlier work demonstrating that A. xylosoxidans undergoes adaptive changes during chronic infections in CF airways, resulting in attenuation of virulence [1112]. Similar persistence mechanisms have also been reported for P. aeruginosa [20]. In a previous study that included all three AX strains examined in the present work, A. xylosoxidans became less motile, slower growing and more resistant to antibiotics over time during chronic infection of CF airways [11]. Whole-genome sequencing revealed that mutations frequently occurred in transcriptional regulators, including loci associated with biofilm formation. Whether these genetic changes are relevant for inter-species interactions remains to be determined.
The clinical impact of these microbial interactions warrants further investigation. One of the key pathogenic features of chronic P. aeruginosa infection is its ability to dominate the lower respiratory microbiota, thereby reducing microbial diversity [121]. Similarly, A. xylosoxidans is also capable of dominating the bacterial community and can reach over 90% relative abundance [22]. Given that both pathogens appear to be equally capable of causing inflammation and lung injury in CF [1315,23], a replacement of P. aeruginosa by A. xylosoxidans would not necessarily lead to improved clinical outcomes. Importantly, co-infection with both pathogens has been associated with lower pulmonary function in people with CF compared to mono-infections with either A. xylosoxidans or P. aeruginosa [16]. This observation raises the question of whether co-localization of the two species affects bacterial virulence. In experimental models of oral biofilms, both biofilm formation and the expression of virulence-associated genes were altered in Streptococcus mutans when co-cultured with other oral pathogens [24]. Investigating virulence factors other than biofilm formation in A. xylosoxidans–P. aeruginosa co-cultures may provide further insights into the clinical impact of these interactions.
A key limitation of this study is the potential discrepancy between in vitro observations and in vivo biofilm formation within the CF airways. Specifically, bacterial adhesion mechanisms in the lung environment may differ markedly from those observed in the microtiter well. Notably, previous studies have demonstrated competition on agar between P. aeruginosa and A. xylosoxidans in terms of growth, motility and pigmentation [17], suggesting that factors other than adhesion to microtiter wells are involved. Additionally, bacteria may respond differently to the in vitro growth conditions used in this study compared to the lung environment, where they may be under pressure from host defences, antibiotics and other microorganisms. Future studies could incorporate more in vivo-like conditions, such as using artificial CF sputum medium or investigating the effects of antibiotic pressure on microbial interactions. Furthermore, it would be of interest to select a biofilm-producing strain of A. xylosoxidans to study whether the inhibition may also be exerted by P. aeruginosa. Another technical limitation is the phenotypic similarity of A. xylosoxidans and P. aeruginosa on agar plates, as well as their shared resistance to commonly used antibiotics. These features prevented the use of c.f.u. counts and selective agar to quantify growth in co-culture experiments. Instead, we used qPCR and species-specific primers to estimate bacterial load via DNA quantification.
Given the increasing threat posed by antibiotic-resistant micro-organisms, bacterial biofilms are an important target for the development of novel therapeutic strategies [2527]. The observed inhibition of P. aeruginosa biofilms by A. xylosoxidans warrants further investigation to identify specific molecular mechanisms involved. Such insights could improve our understanding of interspecific interactions and potentially identify new therapeutic targets against biofilm-associated infections, especially in the context of CF airway infections.
Supplementary material
10.1099/jmm.0.002051Uncited Supplementary Material 1.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cuthbertson L Walker AW Oliver AE Rogers GB Rivett DW et al Lung function and microbiota diversity in cystic fibrosis Microbiome 202084510.1186/s 40168-020-00810-332238195 PMC 7114784 · doi ↗ · pubmed ↗
- 2Flight WG Smith A Paisey C Marchesi JR Bull MJ et al Rapid detection of emerging pathogens and loss of microbial diversity associated with severe lung disease in cystic fibrosis J Clin Microbiol 2015532022202910.1128/JCM.00432-1525878338 PMC 4473198 · doi ↗ · pubmed ↗
- 3Françoise A Héry-Arnaud G The microbiome in cystic fibrosis pulmonary disease Genes 20201153610.3390/genes 1105053632403302 PMC 7288443 · doi ↗ · pubmed ↗
- 4Cantin AM Hartl D Konstan MW Chmiel JF Inflammation in cystic fibrosis lung disease: pathogenesis and therapy J Cyst Fibros 20151441943010.1016/j.jcf.2015.03.00325814049 · doi ↗ · pubmed ↗
- 5Moreau-Marquis S Stanton BA O’Toole GA Pseudomonas aeruginosa biofilm formation in the cystic fibrosis airway Pulm Pharmacol Ther 20082159559910.1016/j.pupt.2007.12.00118234534 PMC 2542406 · doi ↗ · pubmed ↗
- 6Ciofu O Tolker-Nielsen T Jensen PØ Wang H Høiby N Antimicrobial resistance, respiratory tract infections and role of biofilms in lung infections in cystic fibrosis patients Adv Drug Deliv Rev 20158572310.1016/j.addr.2014.11.01725477303 · doi ↗ · pubmed ↗
- 7Biswas L Götz F Molecular mechanisms of Staphylococcus and Pseudomonas interactions in cystic fibrosis Front Cell Infect Microbiol 20211182404210.3389/fcimb.2021.82404235071057 PMC 8770549 · doi ↗ · pubmed ↗
- 8Firmida MC Pereira RHV Silva EASR Marques EA Lopes AJ Clinical impact of Achromobacter xylosoxidans colonization/infection in patients with cystic fibrosis Braz J Med Biol Res 201649 e 509710.1590/1414-431X 2015509726909788 PMC 4792508 · doi ↗ · pubmed ↗