Refined cell transfer model reveals roles for Ascl2 and Cxcr3 in splenic localization of mouse NK cells during virus infection
Laura M Canaday, Andrew Cox, H Alex Feldman, Harsha Seelamneni, Ayad Ali, Jasmine A Tuazon, Lorena Botero Calderon, Sierra N Bennett, Allison Yan, Megan Wilson, Vijayakumar Velu, Stephen N Waggoner

TL;DR
Researchers improved a cell transfer model to study how NK cells position themselves in the spleen during viral infections, revealing the roles of ASCL2 and CXCR3.
Contribution
A refined cell transfer model using CRISPR-generated NK cells and identification of ASCL2 and CXCR3 roles in NK cell localization.
Findings
CRISPR-generated JAXBoy NK cells persist better in recipients than conventional CD45.1 BoyJ NK cells.
CXCR3 repositions NK cells in the spleen's white pulp after infection, aiding immunoregulation.
ASCL2 is essential for recruiting NK cells into the spleen and white pulp.
Abstract
Cell transfer experiments complement the rigorous investigation of antiviral and antitumor functions of natural killer (NK) cells. Success in these endeavors is enhanced by expansion of small numbers of input NK cells driven by viral antigens or homeostatic proliferation in immunodeficient hosts. In contrast, analysis of other NK-cell functions, including immunoregulation, are non-proliferative and require an intact immune system in recipient mice. We reveal poor persistence of conventional congenic (CD45.1) BoyJ NK cells following adoptive transfer in comparison to CRISPR-generated CD45.1+ (JAXBoy) NK cells. Reciprocal transfers between C57BL/6 and JAXBoy mice substantially improve seeding and maintenance of donor NK cells. Using this system, we confirm that CXCR3 re-positions NK cells in the white pulp of the spleen after infection, which is vital for immunoregulation. Moreover, we…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
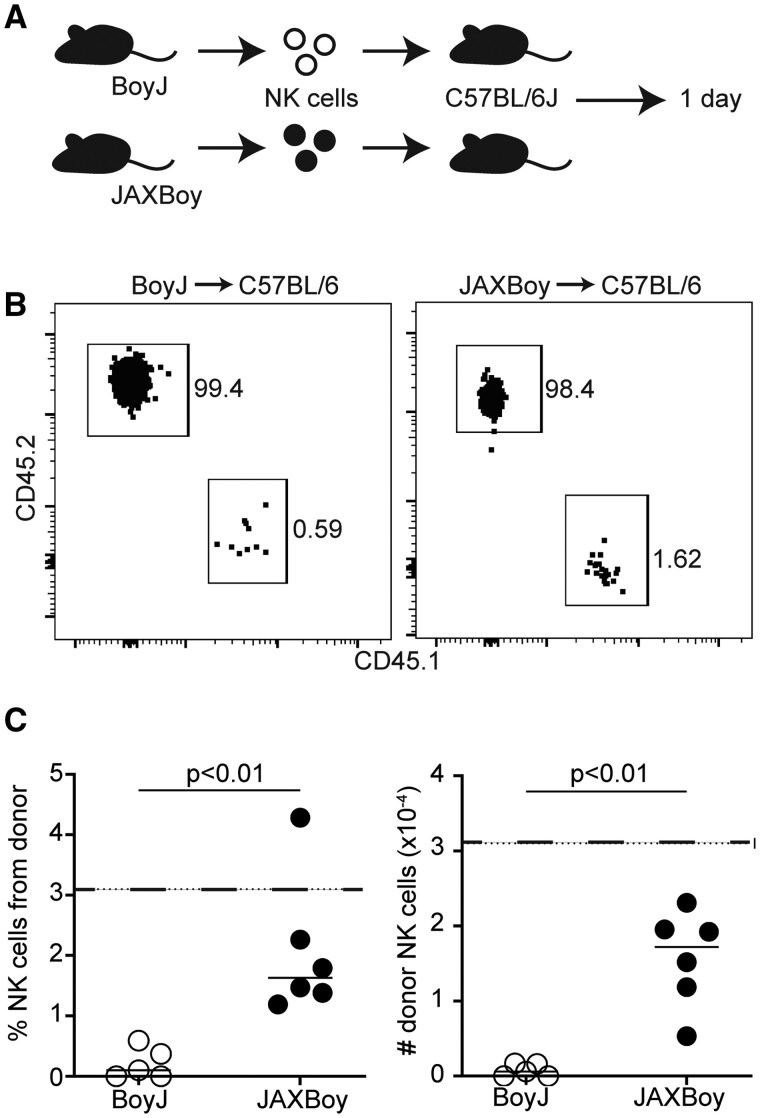 Figure 1
Figure 1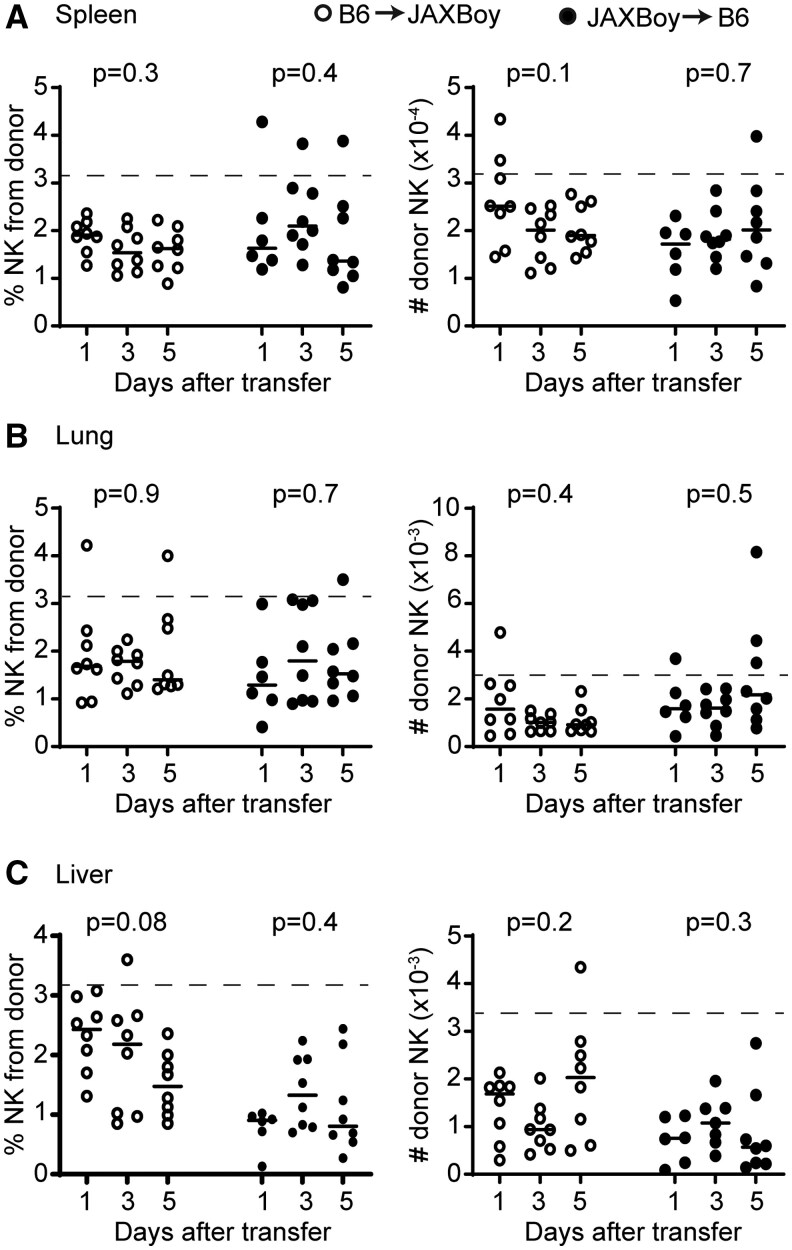 Figure 2
Figure 2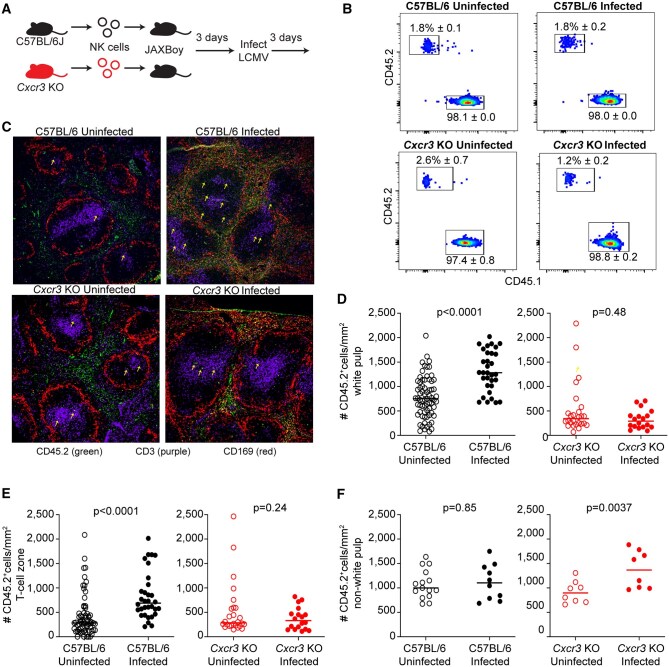 Figure 3
Figure 3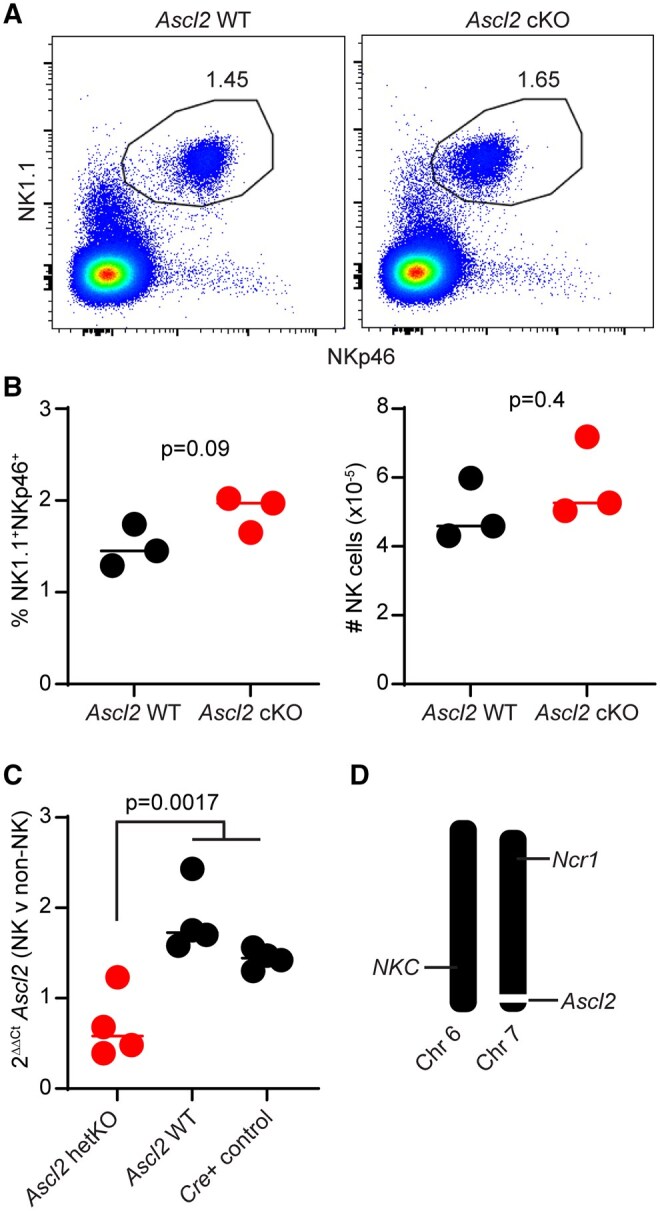 Figure 4
Figure 4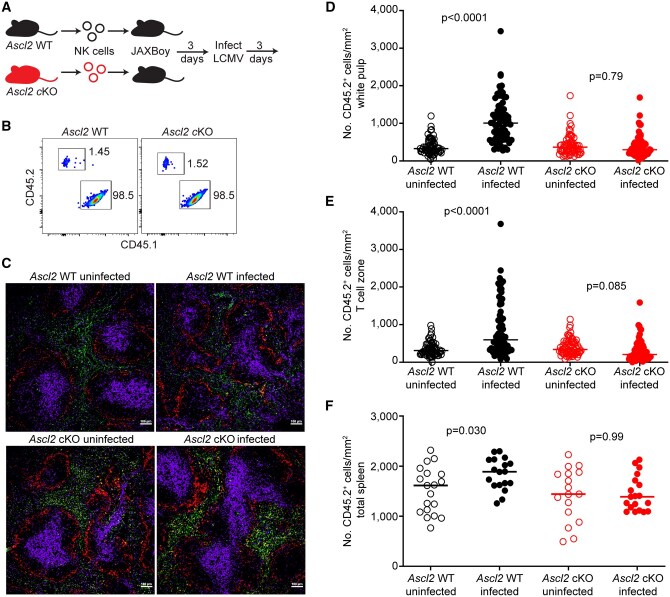 Figure 5
Figure 5| JAXBoy into C57BL/6 transfer | ||||||
|---|---|---|---|---|---|---|
| JAXBoy donor NK cells | C57BL/6 host NK cells | |||||
| Days | CD27-CD11b+ | CD27+CD11b+ | CD27+CD11b- | CD27-CD11b+ | CD27+CD11b+ | CD27+CD11b- |
|
| 4.6 ± 0.02 | 21.6 ± 0.03 | 65.9 ± 0.1 | 6.8 ± 0.03 | 25.1 ± 0.1 | 60.3 ± 0.1 |
|
| 2.7 ± 0.02 | 20.9 ± 0.03 | 68.5 ± 0.04 | 5.6 ± 0.03 | 25.2 ± 0.1 | 59.2 ± 0.1 |
|
| 3.6 ± 0.01 | 14.8 ± 0.03 | 73.5 ± 0.1 | 5.0 ± 0.03 | 21.0 ± 0.1 | 66.0 ± 0.1 |
- —National Institutes of Health10.13039/100000002
- —Pediatric Scientist Development Program supported
- —NIH10.13039/100000002
- —Infectious Disease Clinical Research Consortium Pilot Award
- —Cincinnati Children’s Research Foundation10.13039/100010327
- —Cincinnati Research In Residency Program
- —Albert J Ryan Foundation
- —Cincinnati Children’s Hospital Flow Cytometry Core
- —DK078392
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsImmune Cell Function and Interaction · T-cell and B-cell Immunology · CAR-T cell therapy research
Introduction
Natural killer (NK) cells are innate lymphocytes with critical roles in the killing of virus-infected cells and cancers.1^,^2 NK cells also determine the magnitude and quality of T and B cell responses via perforin-dependent killing of a subset of activated CD4+ T cells.3–5 Enrichment of NK cells within the T and B cell rich white pulp regions of lymphoid tissues after infection is essential for this immunoregulation.6^,^7 Specifically, infection results in elevated expression of CXCR3 ligands in the white pulp that promotes transient re-localization of NK cells into T cell zones, thereby facilitating physical interactions between NK cells and target T cells.
Investigations of NK cell immunoregulatory activity have largely depended upon genetic or antibody-mediated depletion of NK cells or on the generation of mixed bone marrow chimeras.8–15 These methods aid in demonstrating the necessity of NK cells or specific NK-cell mediators in immunoregulation but are inadequate for “rescue” studies that show sufficiency of these cells or genes of interest. Adoptive transfer of NK cells between mice would be an ideal tool to evaluate rescue of immunoregulatory activities. However, traditional adoptive transfer systems are complicated by high cell count requirements and limited proliferation or survival of donor NK cells in recipient animals with intact immune systems.
Here, we developed a protocol for the adoptive transfer of NK cells into immunocompetent animals that is based on CRISPR-generated congenic mice (JAXBoy) that overcome genetic issues with conventional congenic BoyJ mice models.16 We use this system to define the necessity of the chemokine receptor CXCR3 and the migration-associated factor ASCL2 in the enrichment of NK cells in the splenic white pulp after infection. Our results also reinforce the inaccuracy of presumptions regarding the degree of genetic differences between C57BL/6 and congenic BoyJ mice commonly used in cell transplantation studies.16^,^17
Materials and methods
Mice
C57BL/6 (Strain no. 000664), B6.SJL-Ptprc^a^Pepc^b^/BoyJ (BoyJ) (Strain no. 002014), C57BL/6J-Ptprc^em6Lutzy^/J (JAXBoy) (Strain #033076), and B6.129P2-Cxcr3^tm1Dgen^/J (CXCR3^KO^) (Strain no. 005796) mice were purchased from Jackson Laboratory (Bar Harbor, Maine, USA). Ncr1-iCre mice were a gift from Eric Vivier (Marseille, France) and Ascl2-floxed mice (See details below) were a gift from Hans Clevers (Utrecht, Netherlands) and James Bridges.18^,^19 Ncr1-iCre mice were bred with Ascl2-floxed mice and littermates were used in experiments. Both male and female mice between 8–20 weeks of age were routinely utilized in experiments with no apparent sexual dimorphism in resulting data, thus both sexes were combined in data presentation. Mice were housed under barrier conditions and experiments performed under ethical guidelines approved by the Institutional Animal Care and Use Committees of Cincinnati Children’s Hospital Medical Center in compliance with the US Department of Health and Human Services Guide for the Care and Use of Laboratory Animals. Staff responsible for processing samples and acquiring data were blinded to the genotype and treatment status of experimental groups of animals.
Ascl2-floxed mice
Genetic editing was initially performed in 129/Ola-derived IB10 embryonic stem cells before extensive in-house backcrossing of Ascl2-floxed mice (8 generations) onto the C57BL/6 genetic background. Key NK cell receptor expression was determined by flow cytometry analysis of blood or spleen cells using the following Biolegend antibodies: Ly49H (3D10)- PE/Cyanine7, NK-1.1 (PK136)- BV421, and CD3ε (500A2)- FITC. Taconic Biosciences SNP Genotyping service was used on DNA from a female Ascl2-floxed mouse to assess degree of fixation to the C57BL/6J genetic background.
Ascl2 and Cre genotyping
Genomic DNA was extracted from tail clips using the HotSHOT DNA extraction protocol (Bento Bioworks). PCR was performed with the following primers: Ascl2 flox forward [GCG AAG CAG TAA GGG AGA CAC G], Ascl2 flox revere [AGA ACC TGC CCG CCG TGA C], iCre forward [TTC TCT GAA CAC ACC TGG AAG ATG], and iCre reverse [AGA GTT CTC CAT CAG GGA TCT GAC]. For Ascl2 polymerase chain reaction (PCR), reactions combined 2 µl DNA, 1 µl of each 10 µM primer, and 10 µl of 2X Econotaq Plus Green mastermix (Biosearch Technologies, Cat no. F93481-1) in a total volume of 20 µl. The PCR cycling conditions were 94°C for 3 min, followed by 30 cycles of 94°C for 30 s, 60°C for 30 s, and 72°C for 1 min, with a final extension at 72°C for 7 min. PCR products were separated on a 2% agarose gel and visualized under UV light, with the floxed allele at 433 bp and the wild-type allele at 383 bp. For Ncr1-iCre PCR, reactions combined 1 µl DNA, 2 µL of each 10 µM primer, and 6 µL of 2X Econotaq Plus Green mastermix (Biosearch Technologies, Cat no. F93481-1) in a total volume of 20 µL. The PCR cycling conditions were 95°C for 90 s, followed by 32 cycles of 95°C for 30 s, 63°C for 20 s, and 72°C for 30 s, with a final extension at 72°C for 30 s. PCR products were separated on a 2% agarose gel and visualized under UV light, with the KI band at 348 bp.
Confirmation of selective Ascl2 targeting in NK cells
NK cells were enriched from Cre(+) control, Cre(+) Ascl2^fl/WT^ and Cre(-) Ascl2^fl/WT^ littermate mice using the anti-NKp46 MicroBead Kit (Miltenyi, Cat no. 130-095-390) according to the kit protocol. Non-NK cells were obtained from the column flowthrough. A subset of cells was taken for pre- and post-sorting purity checks using the following Biolegend antibodies: CD335 (29A1.4)- BV605, CD3ε (500A2)- FITC, and Zombie NIR Fixable Viability Kit. Remaining enriched NK cells were then immediately lysed to extract RNA using the RNeasy mini kit (Qiagen, Cat no. 74104). RNA was reverse transcribed into cDNA using the iScript Reverse Transcription Supermix for RT-qPCR kit (BioRad, Cat no.1708840). RT-qPCR was performed using TaqMan Gene Expression Assays to detect Ascl2 (Mm01962673_s1) and Gapdh (Mm99999915_g1). Reactions were run on QuantStudio 3 and 5 Real-Time PCR Systems (ThermoFisher). Ascl2 Ct was normalized to Gapdh Ct to determine ΔCt. Then the NK cell ΔCt was normalized to non-NK ΔCt for each mouse, yielding ΔΔCt. The fold-change in transcripts was then quantified (2^-ΔΔCt^).
Adoptive transfer of NK cells
JAXBoy, BoyJ, or C57BL/6 spleens were harvested and processed into single cell suspensions (see tissue processing) from which NK cells were column enriched by depletion of non-NK cells per manufacturer’s instructions (Miltenyi Biotec, Germany) to >90% purity. One million NK cells in 100 μl Hank’s Balanced Saline Solution (HBSS) were injected retro-orbitally with a 28G needle (BD Biosciences, San Jose, CA) into C57BL/6 or JAXBoy recipient mice. Transfers were done from donor mice of same sex as recipient mice.
Virus injection
Mice were infected with the Armstrong strain of LCMV via intraperitoneal injection of 5 × 10^4^ plaque forming units per mouse in 200 μl of HBSS using a 28G needle. Viral stocks were previously generated in house via propagation on BHK21 cells with viral titers determined using Vero cells.20
Tissue processing and cell staining for flow cytometry
Spleens, lungs, and livers were harvested from mice following euthanasia. Single-cell leukocyte suspensions were prepared from spleens by mechanical homogenization of tissues between frosted glass microscope slides (VWR, Radnor, PA) and filtration through a 70 μm nylon mesh strainer with RPMI medium 1,640 + 10% fetal bovine serum + 50,000U penicillin-streptomycin (Gibco, Waltham, MA) + 2 mM L-glutamine (Gibco, Waltham, Massachusetts, USA). Lungs were homogenized by crushing with the rubber end of a 10 mL syringe through a 70 μm nylon mesh with RPMI medium 1640 + 10% fetal bovine serum + 50,000U penicillin-streptomycin + 2 mM L-glutamine. Livers were homogenized with surgical scissors in RPMI + 10% fetal bovine serum. After resuspension, cells were centrifuged at 300xg for 10 min. Cell pellets were homogenized in a 33% Percoll solution diluted in RPMI 1,640 medium. All cell suspensions were then subjected to red blood cell lysis via ACK lysing buffer for 5 min at 37°C. Following lysis of red blood cells, cells were plated at 2 × 10^6^ per well in 96-well round-bottom plates and immediately subjected to flow staining. Leukocytes were washed and resuspended in FACS buffer (Hanks Buffered Salt Solution + 5% fetal bovine serum + 0.5 mM EDTA), stained for 5 min at room temp with a live/dead stain, washed twice with FACS buffer, and nonspecific Fc receptor-mediated binding was then blocked via a 5-min incubation at 4°C in anti-mouse CD16/CD32 (BD BioSciences, San Jose, California, USA). Cells were stained with the following antibodies: CD4 (GK1.5)- BV737 [1:200], CD8α (53-6.7)- PE-CF594 [1:200], CD19 (1D3-CD19)-PerCP-Cy5.5 [1:200], CD49a (HMa1)- APC [1:100], NK1.1 (PK136)- BV711 [1:200], CD49b (DX5)- PE [1:100], NKp46 (29A1.4)- BV605 [1:50], CD45.1 (A20)- AF700 [1:100], CD45.2 (104)- AF488 [1:100], CD11b (M1/70)- BV786 [1:100], CD27 (LG.3A10)- BV421 [1:100], and live/dead- Zombie UV [1:1000]. All antibodies were purchased from BioLegend (San Diego, California, USA) or BD Biosciences (San Jose, California, USA). Cells were surface stained for 30 min at room temperature. Following staining, cells were washed and fixed with BD Cytofix buffer (BD Biosciences, San Jose, California, USA) for 5 min at 4°C. Cells were washed twice and resuspended in 200 µl FACS buffer and analyzed on a BD LSR Fortessa cytometer. All data were analyzed using FlowJo software (FlowJo Inc., Ashland, Oregon, USA).
Tissue processing for immunofluorescent microscopy
Tissues to be analyzed by fluorescence microscopy were placed into 4% paraformaldehyde solution for 5 hr followed by overnight dehydration in a 30% sucrose solution at 4°C (Sigma-Aldrich, St Louis, Missouri, USA). Samples were washed with phosphate buffered saline prior to embedding within optimal cutting temperature (OCT) media (Sakura Finetek, Maumee, Ohio, USA) and frozen using a dry ice slurry in 100% ethanol and stored at −20°C. Tissues were sectioned (7 μm thick) using a cryostat (Leica CM3050 S) and affixed to positively charged Denville slides (Thomas Scientific, Swedesboro, New Jersey, USA). Slides were dried at room temperature for 10 min preceding a 10-min fixation in −20°C chilled 100% acetone. Slides were subsequently dried at room temperature for 10 min and washed twice in chilled phosphate-buffered saline before being placed in saturation buffer containing 10% normal donkey serum (Sigma-Aldrich, St Louis, Missouri, USA) and 0.1% Triton-X (Sigma-Aldrich, St Louis, Missouri, USA) in phosphate-buffered saline for blocking at room temperature for 45 min. Slides were incubated overnight at 4°C with a primary antibody cocktail: 1:100 mouse anti-CD45.2-AF488 (104, BioLegend, San Diego, California, USA), 1:200 hamster anti-CD3e-AF594 (500A2, BioLegend, San Diego, California, USA) and 1:200 rat anti-CD169-AF647 (3D6.112, BioLegend, San Diego, California, USA). The following day, slides were washed twice in chilled phosphate-buffered saline, mounted with ProLong diamond mounting media (Thermo-Fisher Scientific, Waltham, Massachusetts, USA), covered with a coverslip, and allowed to cure overnight prior to imaging.
Confocal microscopy and NK-cell enumeration
Confocal imaging was performed using a Laser Scanning Nikon A1RSi Inverted Confocal Microscope with NIS Elements Confocal software. Z-stacked tissue images were acquired through a 10× objective (Nikon Plan Apo λ) from which a maximum intensity projection was generated prior to cell enumeration. Using tools available in NIS Elements Analysis software and guided by CD169- and CD3-staining, we drew borders around the white pulp (CD169 boundary) and T cell zones (CD3 boundary) while visualizing only each individual channel. The channel for donor NK cell staining was hidden at this stage to prevent bias of NK cell localization on drawing of these boundaries. Thereafter, NK cells located in the previously defined T cell zones and white pulp regions were enumerated using unbiased NIS Elements-derived algorithms and plotted using GraphPad Prism (San Diego, California, USA). To quantify NK cells outside the white pulp, the quantity of white pulp NK cells was subtracted from the total number of NK cells detectable within a section and normalized to mm^2^. Brightness and contrast for each representative image were adjusted equally across all channels using NIS Elements Analysis.
Statistical analysis and experimental rigor
All individual datapoints are represented in plots along with group mean, while tabular data are presented as the mean and standard deviation. Statistical differences between control and experimental groups were calculated using statistical tests indicated in each figure legend, including unpaired 2-tailed Mann–Whitney (non-normally distributed data with unequal variance across 2 groups), unpaired 2-tailed Student’s T test (normally distributed data with equal variance across 2 groups), Kruskal–Wallis (non-parametric comparison of 2 groups across time) with post-hoc Dunn’s test (quantification of Ascl2 levels), or Welch ANOVA with Dunnett’s T3 multiple comparisons testing (for more than 2 groups displaying unequal variance). A *P-*value of less than 0.05 was considered significant. Graphing and statistical analysis were routinely performed using GraphPad Prism (San Diego, California, USA). Researchers performing the tissue processing and experimental measurements were blinded to genotype and treatment conditions of the animals. This resulted in randomization of the processing order.
Results
Traditional CD45.1 congenic (BoyJ) NK cells fail to thrive upon adoptive transfer
Allelic variation among inbred mouse strains at the Ptprc locus (encoding Ly5/CD45) has been exploited as a tool to track donor leukocytes after adoptive transfer based on the specificity of monoclonal antibodies for the Ptprc^a^ (Ly5.1/CD45.1) and Ptprc^b^ (Ly5.2/CD45.2) alleles.16 Several groups have successfully tracked donor CD45.1+ NK cells in recipient CD45.2+ mice for extended periods of time, albeit in contexts where the donor NK cells undergo significant clonal expansion (eg, murine cytomegalovirus infection)11^,^21 or possess homeostatic advantages over host cells (eg, common gamma chain deficient mice).15
We aimed to use similar adoptive transfer model systems to study NK cell immunoregulation of T and B cells in immunocompetent mice after infection or vaccination. To our surprise, adoptive transfer of splenic NK cells from conventional B6.SJL-Ptprc^a^Pepc^b^/BoyJ (BoyJ) mice into C57BL/6J mice (Fig. 1A) resulted in a poor or exceedingly short-lived “take” (ie, recovery from recipient tissues) of donor cells within 24 hr of cell transfer (Fig. 1B). Based on limiting dilution studies, the expected take of donor lymphocytes (ie, T cells) in the spleen of recipient animals is 3.1%.22 In contrast to this expectation, donor BoyJ NK cells consistently comprised less than 0.6% of the overall pool of NK cells in recipient mice (Fig. 1C, left), with fewer than 1,000 donor CD45.1+ NK cells recovered from host spleens (Fig. 1C, right). Similar results were obtained with two-fold higher or lower inoculum sizes of donor NK cells (data not shown). Thus, BoyJ-derived NK cells fail to persist after transfer into immunocompetent mice, presumably due to minor histocompatibility mismatch or the effects of passenger genes located near the Ptprc locus.16
Superiority of JAXBoy over BoyJ NK cells in adoptive transfer experiments. (A) 1,000,000 splenic NK cells from BoyJ (CD45.1+) or JAXBoy (CD45.1+) mice were injected retro-orbitally into C57BL6/J recipients (n = 5–6/group). (B) Representative flow plots of gated NK cells (CD4- CD8- CD19- CD49a- NK1.1+ CD49b+ NKp46+) from host (CD45.2+) or donor (CD45.1+) in recipient spleen 1 d after transfer. (C) The proportion and number of donor-derived NK cells in the spleens of each recipient mouse. Data are representative of 2 independent experiments. Solid lines represent mean and dotted lines represent predicted cell take (3.1%, 31,000 cells). Statistically significant differences determined by Mann–Whitney test.
CRISPR-generated CD45.1+ congenic (JAXBoy) mice are a superior platform for NK cell transfer
The Jackson Laboratory recently developed coisogenic JAXBoy mice in which the Ptprc^a^ (Ly5.1/CD45.1) allele was introduced into C57BL/6J mice using CRISPR-Cas9.23^,^24 This editing introduced a lysine to glutamic acid mutation at amino acid 302 (K302E; AAA->GAA) and a silent nucleotide change N303N (AAC->AAT), resulting in reactivity with anti-CD45.1 monoclonal antibodies without the confounding genetics of proximal genomic regions inherited from SJL founders in conventional BoyJ mice.16
We transferred splenic NK cells from JAXBoy mice into C57BL/6 mice (Fig. 1A) and analyzed recovery of donor cells from recipient mouse tissues over the ensuing five days. The average recovery of CD45.1+ JAXBoy donor NK cells in the spleen of recipient C57BL/6 mice was five times greater than that observed with BoyJ donor NK cells (Fig. 1C). The proportion (Fig. 2A) and number (Fig. 2B) of CD45.1+ JAXBoy donor NK cells recovered from spleens of recipient C57BL/6 mice was relatively stable over a five-day window. Similar stability was seen in the recovery of CD45.1+ JAXBoy donor NK cells from recipient mouse lungs and liver over the same time span (Fig. 2C–F). A reciprocal transfer of splenic CD45.2+ C57BL/6 NK into CD45.1+ JAXBoy recipient mice revealed a similarly strong take of CD45.1+ donor cells comprising an average of 1%–2% of total NK and measurable numbers of donor cells in each host tissue that remained relatively stable over an experimental window of five days (Fig. 2). In either transfer direction, donor and host splenic NK cells in the C57BL/6-JAXBoy system displayed similar phenotypic patterns of distribution across maturation states comprising immature (CD11b^-^ CD27^+^), transitional (CD11b^+^ CD27^+^), and mature (CD11b^+^ CD27^-^) subsets of cells (Table 1).25 The data demonstrate that JAXBoy mice represent a superior model system for adoptive transfer studies of mouse NK cells, potentially due to the precision engineering that limits genetic effects adjacent to the Ptprc locus that are seen in conventional BoyJ mice.16
Persistence of donor NK cells in reciprocal transfer between C57BL/6 and JAXBoy mice. 1,000,000 C57BL/6 (CD45.2+, open circle) or JAXBoy (CD45.1+, closed circle) isolated splenic NK cells were transferred by retro-orbital injection into JAXBoy or B6 mice, respectively (n = 6–8/group). In total, 1, 3, and 5 d later, recipient mouse (A) spleens, (B) lungs, and (C) liver were subjected to flow cytometry to determine proportion (among total CD4- CD8- CD19- CD49a- NK1.1+ CD49b+ NKp46+ NK cells) and number of donor-derived NK cells. Data are representative of 2 independent experiments. Solid lines represent mean and dotted lines represent predicted cell takes. Statistically significant differences between time points for each transfer type were determined by Kruskal–Wallis test.
CXCR3-dependent localization of donor NK cells in splenic white pulp after infection
We recently demonstrated a transient CXCR3-dependent enrichment of NK cells in the white pulp of the spleen and lymph nodes after infection with the Armstrong strain of lymphocytic choriomeningitis virus (LCMV).7 This localization was required for NK cell suppression of T cell responses. To verify that donor NK cells exhibit similar CXCR3-dependent localization in recipient mice after infection,26 we transferred CD45.2+ NK cells from the spleens of C57BL/6 or Cxcr3-deficient (Cxcr3 KO) mice into JAXBoy (CD45.1+) recipient mice (Fig. 3A). Three days after cell transfer, recipient mice were intraperitoneally infected with LCMV Armstrong or left uninfected for three additional days. The recovery of CD45.2+ donor NK cells from either wild-type C57BL/6 or Cxcr3 KO donors within uninfected and infected recipient mice was assessed via flow cytometry (Fig. 3B). We also assessed donor (CD45.2+) NK cell positioning within the white pulp (defined by CD169+ margin surrounding a CD3+ T cell zone) using confocal microscopy at day three of infection (Fig. 3C). Infection triggered a statistically significant increase in the number of wild-type donor CD45.2+ (C57BL/6) NK cells in both the white pulp (Fig. 3D, left) and T-cell zones (Fig. 3D, right) of JAXBoy recipients relative to uninfected control hosts. No statistically significant enrichment of Cxcr3 KO donor CD45.2+ NK cells was observed in either the white pulp (Fig. 3E, left) or T-cell zones (Fig. 3E, right) of recipient JAXBoy mice after virus infection. Given the vanishingly small number of NK cells present in lymph nodes in prior studies,7 we focused the present work exclusively on donor cell localization in the spleen. By subtracting white pulp localized NK cells from the total, we estimated the number of NK cells in other regions of the spleen (Fig. 3F). This analysis revealed similar numbers of WT NK cells in non-white pulp regions before and after infection, while Cxcr3 KO NK cells were enriched in non-white pulp regions only after infection. These results confirm prior studies demonstrating CXCR3-dependent transient enrichment of NK cells in the white pulp regions of the spleen after infection,7 yet suggest that CXCR3 is not required for infection-induced accumulation of NK cells in the spleen. Moreover, these data highlight the utility of this NK-cell transfer model to aid the discovery of additional mediators governing critical immunoregulatory positioning of NK cells.
CXCR3 is required for donor NK cells to localize in spleen white pulp and T cell zones after infection. (A) 1,000,000 CD45.2+ C57BL/6 or Cxcr3 KO NK cells were retro-orbitally injected into JAXBoy (CD45.1+) recipient mice (n = 3/group). Three days after transfer, recipient mice were infected with LCMV or left uninfected. (B) Three days later, take of donor cells (CD45.2+) was confirmed via flow cytometry in recipient mice (CD45.1+). (C) CD45.2+ (green) donor NK cell positioning within spleen relative to CD169+ marginal zone macrophages (Red) and CD3+ T cells (Purple) assessed by confocal microscopy for both C57BL/6 and Cxcr3 KO donor NK cells. Arrows demarcate representative donor NK cell positioning within these sections. The relative positioning of C57BL/6 or Cxcr3 KO donor NK cells in the (D) CD169+ bound white pulp or (E) CD3+ T cell zones were quantified in 10–12 follicles per recipient mouse (horizontal line represents mean). Open symbols reflect uninfected mice, and closed symbols reflect mice infected with LCMV. (F) The number of C57BL/6 or Cxcr3 KO donor NK cells outside the white pulp was calculated for each image by subtracting the number of NK cells quantified within the CD169-defined white pulp from the total number of NK cells in the image. Data are representative of 2 independent staining experiments. Statistical significance differences in donor NK cell densities in various spleen regions between uninfected and infected host mice were determined by Mann–Whitney analysis.
Role of ASCL2 in NK cell migration to white pulp during infection
Achaete-Scute Complex Homolog 2 (ASCL2) is a transcription factor implicated in the differentiation of follicular helper T cells and their localization within lymphoid tissues.27 ASCL2 transcripts are measurably expressed in mouse28 and human29 NK cells, but the function of this gene in innate lymphocytes remains unexplored. We hypothesized that NK cells may require ASCL2 for precise localization to subdomains of lymphoid tissues during infection (ie, infiltration of white pulp). We generated mice in which NK cells lack Ascl2 by crossing Ncr1-Cre18 animals with mice in which the second exon of Ascl2 is flanked by LoxP sites.19 The resulting Cre+ conditional Ascl2-deficient mice (Ascl2 cKO) were viable, fertile, and contained statistically similar proportions and numbers of NK cells in their spleens in comparison to control Ascl2-floxed Cre-negative (Ascl2 WT) littermates (Fig. 4A, B). Of note, the MFI of NKp46 expression is reduced in Ascl2 cKO NK cells due to hemizygous expression of NKp46 in Ncr1-iCre+ animals, as previously described.18
Ascl2 is dispensable for the development of mouse NK cells. (A) Spleens from Ascl2 WT (Cre-negative Ascl2fl/fl,) and Ascl2 conditional knockout (cKO, Cre+ Ascl2fl/fl) mice (n = 3/group) were collected and stained to quantify NK cells by flow cytometry. (B) The percentage and number of NK cells (CD3- NK1.1+NKp46+) in the spleen is quantified and plotted for Ascl2 WT (black) and Ascl2 cKO (red). The horizontal line represents median. Statistically significant differences were determined by unpaired Student’s T-test. Data are representative of 2 independent staining experiments. (C) NK cells were isolated by positive selection and corresponding non-NK cells were collected from the spleens of Ncr1-Cre+ Ascl2fl/WT (Ascl2 hetKO, red), Ncr1-Cre(-) Ascl2fl/WT (Ascl2 WT, black), and Ncr1-Cre+ control mice (black). Quantitative PCR amplification of Ascl2 and Gapdh, Ascl2 Ct normalized to Gapdh (ΔCt), NK cell ΔCt was normalized to non-NK ΔCt (ΔΔCt), and fold-change transposition (2-ΔΔCt) were performed. Significant differences between genotypes assessed by Kruskal-Wallis with a post-hoc uncorrected Dunn’s test. (D) Visual representation of SNP genotyping analysis (Table S1) for key chromosomes in Ascl2-floxed mouse colony, where black is homozygous for C57BL/6 SNP and white bars equate with homozygous for non-C57BL/6 SNP. Important NK cell receptor gene complex (NKC) on chromosome 6 and Ncr1 (NKp46) region on chromosome 7 are homozygous C57BL/6, while locus around floxed Ascl2 gene on chromosome 7 is not C57BL/6-derived, as expected.
Although detection of ASCL2 protein proved challenging, we confirmed by real-time quantitative PCR that Ascl2 mRNA expression was selectively reduced in NK cells but not in other immune cells of mice with conditional deletion of a single Ascl2 allele relative to control mice (Fig. 4C). Since Ascl2 genetic targeting was initially performed in 129/Ola mouse-derived embryonic stem cells, we performed genome-wide SNP genotyping on our colony. After more than 8 backcrosses to C57BL/6, the Ascl2-floxed mice were >96% C57BL/6 (Table S1), expressed key C57BL/6-associated NK cell receptors NK1.1 and Ly49H (not shown), and displayed homozygous C57BL/6 congenic genotype across chromosomes 6 (site of NK cell receptor gene complex) and *Ncr1-*end of chromosome 7 while exhibiting non-C57BL/6 DNA proximal to the floxed Ascl2 alleles (Fig. 4D and Table S1).
We intravenously transferred splenic CD45.2+ NK cells from either Ascl2 cKO or Ascl2 WT mice into JAXBoy mice 3 d before infection with LCMV Armstrong (Fig. 5A). The recoveries of donor CD45.2+ Ascl2 WT or Ascl2 cKO NK cells from the spleens of recipient CD45.1+ JAXBoy mice were assessed by flow cytometry (Fig. 5B). We analyzed the positioning of donor NK cells in recipient mouse spleens on day three of infection and in uninfected hosts (Fig. 5C). Infection triggered a statistically significant increase in the number of Ascl2 WT donor NK cells present within the white pulp or T cell zones (Fig. 5D, E) compared to uninfected mice. In contrast, Ascl2 cKO NK cells were not enriched in the white pulp or T cell zones after infection (Fig. 5D, E). In contrast to loss of Cxcr3 in Fig. 3, the accumulation of NK cells in total spleen after infection was only observed for Ascl2 WT but not Ascl2 cKO NK cells (Fig. 5F). These data indicate that Ascl2 is important for infection-induced accumulation of NK cells in the splenic white pulp potentially by regulating their recruitment to the spleen itself.
Ascl2 is required for NK cell positioning in splenic white pulp after infection. (A) Ascl2 WT or Ascl2 cKO NK cells (CD45.2+) NK cells were adoptively transferred into JAXBoy (CD45.1+) mice (n = 2/group) 3 d prior to infection with LCMV or being left uninfected for an additional 3 d. (B) The take of CD45.2+ donor cells was confirmed via flow cytometry in uninfected CD45.1+ recipient mouse spleens. (C) CD45.2+ (green) donor NK cells were quantified based on positioning within spleen using microscopy relative to CD169+ marginal zone macrophages (red) and CD3+ T cells (purple). Representative images at 10× magnification are shown. (D–F) The densities of CD45.2+ Ascl2 WT (black circles) or Ascl2 cKO (red circles) donor NK cells in specific splenic regions of uninfected (open circles) or infected (closed circles) host mice were quantified in 10–12 follicles per recipient mouse (horizontal line represents mean). Regions include (D) CD169+ bound white pulp, (E) CD3+ T cell zones, and (F) total spleen. Data are representative of 2 independent staining experiments. Statistically significant differences between uninfected and infected donor NK cell densities in various spleen regions were determined by post-hoc Dunnett’s T3 multiple comparisons testing with Welch ANOVA (latter P values are shown).
Discussion
These results demonstrate the improved utility of CRISPR-generated JAXBoy mice in comparison to conventional BoyJ mice for adoptive transfer and tracking of donor NK cells in immunocompetent recipient animals. This distinction is likely due to the limited number of genetic variants (two nucleotides) introduced into JAXBoy mice to confer CD45.1 reactivity on the C57BL/6 background. In contrast, prior work revealed large and variable co-inheritance of non-C57BL/6 alleles (SJL derived) near the Ptprc locus in conventional BoyJ mice.16 These include genes that affect survival (Bcl2), migration (Cxcr4, Sell), antiviral interferon responses (Rnasel), and immune function (Pdcd1, Il10), any of which are likely important in the survival and positioning of NK cells in cell transfer model systems. The JAXBoy mouse likely holds similar advantages over conventional BoyJ mice in adoptive transfer experiments with other types of CD45-expressing leukocytes. The difference seen in donor cell uptake in our experiments also raises concerns for the use of conventional CD45.1+ congenic mice in other transfer experimental systems, including those where donor T cell or NK cell proliferation may mask these drawbacks.
We present a new cell transfer model applicable to the investigation of intrasplenic migration of NK cells. We use this model to confirm the necessity of CXCR3 for positioning of NK cells in the white pulp near T and B cells after virus infection. Yet, our data additionally indicate that CXCR3 is dispensable for the general accumulation of NK cells in the spleen during infection. The mispositioning of CXCR3-deficient NK cells in the red pulp likely explains the inability of these CXCR3-less cells to regulate T cell responses observed in our prior publication.7 Furthermore, we develop an original strain of mice with conditional deletion of Ascl2 in NK cells and employ this new transfer model to demonstrate a role for ASCL2 in enrichment of NK cells in the spleen and white pulp after virus infection. ASCL2 controls the expression of numerous target genes involved in follicular T cell27^,^30 and germinal center B cell31 biology, including cell migration receptors CXCR5 and CXCR4. ASCL2 also intersects with other transcriptional networks implicated in various facets of NK cell biology, including ETS131 and BLIMP1.30 Thus, the discovery of a role for ASCL2 in NK cell positioning in the spleen is the first step in a new area of research into the role of this transcriptional regulator in NK cell biology. This and future studies are likely to reveal ASCL2-regulated pathways vital for immunoregulatory functions of NK cells that can be targeted to circumvent NK cell suppression of vaccine response or to bolster NK-cell based therapies targeting pathogenic T and B cells in autoimmune disease.
Supplementary Material
vkaf122_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Waggoner SN et al Roles of natural killer cells in antiviral immunity. Curr Opin Virol. 2016;16:15–23.26590692 10.1016/j.coviro.2015.10.008PMC 4821726 · doi ↗ · pubmed ↗
- 2Bjorkstrom NK , Strunz B, Ljunggren HG. Natural killer cells in antiviral immunity. Nat Rev Immunol. 2022;22:112–123.34117484 10.1038/s 41577-021-00558-3PMC 8194386 · doi ↗ · pubmed ↗
- 3Rydyznski CE et al Affinity maturation is impaired by natural killer cell suppression of germinal centers. Cell Rep. 2018;24:3367–3373.e 3364.30257198 10.1016/j.celrep.2018.08.075PMC 6192537 · doi ↗ · pubmed ↗
- 4Rydyznski C et al Generation of cellular immune memory and B-cell immunity is impaired by natural killer cells. Nat Commun. 2015;6:6375.25721802 10.1038/ncomms 7375 PMC 4346304 · doi ↗ · pubmed ↗
- 5Cox A et al Targeting natural killer cells to enhance vaccine responses. Trends Pharmacol Sci. 2021;42:789–801.34311992 10.1016/j.tips.2021.06.004PMC 8364504 · doi ↗ · pubmed ↗
- 6Voskoboinik I , Whisstock JC, Trapani JA. Perforin and granzymes: function, dysfunction and human pathology. Nat Rev Immunol. 2015;15:388–400.25998963 10.1038/nri 3839 · doi ↗ · pubmed ↗
- 7Ali A et al Natural killer cell immunosuppressive function requires CXCR 3-dependent redistribution within lymphoid tissues. J Clin Invest. 2021;131:e 146686.
- 8Ni J et al Adoptively transferred natural killer cells maintain long-term antitumor activity by epigenetic imprinting and CD 4(+) T cell help. Oncoimmunology. 2016;5:e 1219009.27757318 10.1080/2162402 X.2016.1219009 PMC 5048776 · doi ↗ · pubmed ↗