Field responses of Glossina pallidipes and Glossina fuscipes fuscipes tsetse flies to Novel Repellent Blend and Waterbuck Repellent Compounds in Kenya
Benson M. Wachira, Richard Echodu, Johnson O. Ouma, Imna I. Malele, Daniel Gamba, Michael Okal, Kennedy O. Ogolla, Clement Mangwiro, Robert Opiro, Deusdedit J. Malulu, Bernard Ochieng, Raymond E. Mdachi, Paul O. Mireji

TL;DR
This study compares two new repellents for tsetse flies in Kenya, finding one more effective against a specific species.
Contribution
The study evaluates and compares the field efficacy of two novel tsetse fly repellent blends against key fly species in Kenya.
Findings
NRB is significantly more effective than WRC in repelling Glossina pallidipes.
Neither WRC nor NRB effectively repels Glossina fuscipes fuscipes.
NRB could enhance protection against G. pallidipes in trypanosomiasis control programs.
Abstract
Control of tsetse flies constitutes a cornerstone of trypanosomiasis control and elimination efforts in Africa. The use of eco-friendly odor-based bait technologies has been identified as a safer method for control of tsetse flies. These technologies are significantly augmented by development of effective repellents that reduce contact between trypanosome-infected tsetse flies and their vertebrate hosts. Waterbuck Repellent Compounds (WRC) and Novel Repellent Blend (NRB) are recently developed tsetse fly repellent formulations. Information on relative efficacy of these formulations against major tsetse fly vectors of trypanosomiasis in Kenya is limited. Such information can inform choices of repellent technology for optimal control of the flies. Here we assessed relative field responses of Glossina pallidipes and G. fuscipes fuscipes, representative of savannah (morsitans) and riverine…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
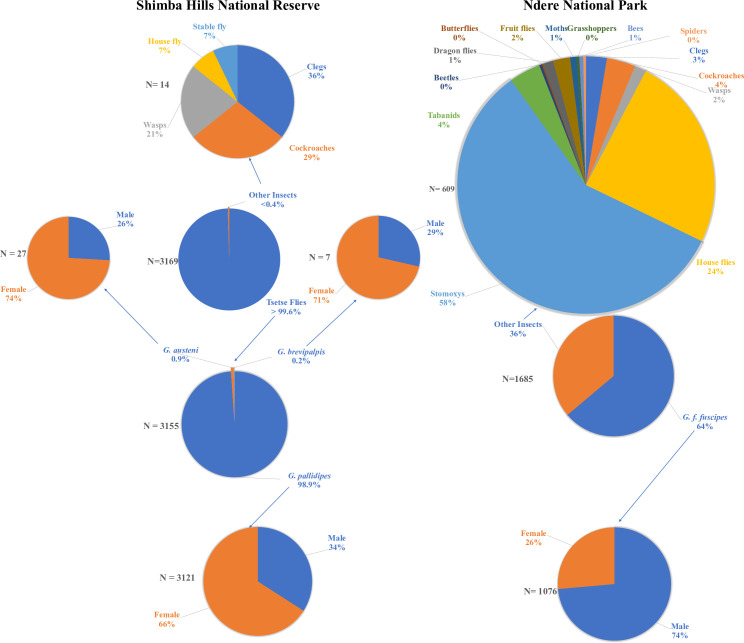 Fig 1
Fig 1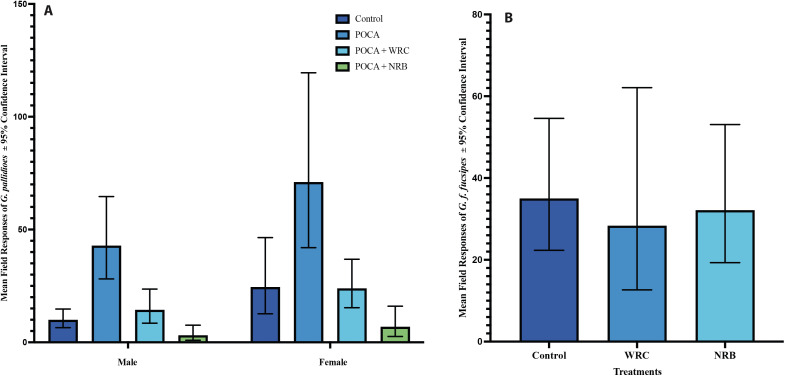 Fig 2
Fig 2- —http://dx.doi.org/10.13039/100000009Foundation for the National Institutes of Health
- —http://dx.doi.org/10.13039/100000009Foundation for the National Institutes of Health
- —Bioinnovate Africa Program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasites and Host Interactions · Parasite Biology and Host Interactions · Insect Utilization and Effects
Introduction
Human African Trypanosomiasis (HAT) and Animal African Trypanosomosis (AAT) are among the most Neglected Tropical Diseases (NTDs) with devastating health and economic consequences in sub-Sahara Africa [1,2]. The AAT is responsible for mortality of about three million cattle and loss of about US$ 4.75 billion per year in terms of agricultural Gross Domestic Product [3]. With no effective vaccines, and limited chemotherapeutic and chemoprophylactic management options against HAT and AAT, control of tsetse flies constitutes a cornerstone in the campaign towards suppression and elimination of trypanosomiasis. Insecticide application to cattle for tsetse control was demonstrated in southern Africa in the mid 1980s, but seemed of limited use where the intention was to remove tsetse flies from spatially wide areas or where cattle were absent or could not be introduced [4]. However, the application is currently considered the most economical method of tsetse fly control [5], with recent application estimated to significantly enhance mortality of the flies by 5–14% daily at the interface of wildlife and livestock in Serengeti, Tanzania [6]. Control of tsetse flies through aerial and ground spraying is limited by detrimental environmental effects of the aerially delivered insecticides on non-target organisms [7]. Alternative eco-friendly bait “pull” technology for tsetse fly control consists of stationary traps and targets that exploits visual and olfactory responses of tsetse flies to their hosts [8]. The traps and targets are typically baited with attractant odor blends that mimics the smell of buffalo (Syncerus caffer), a preferred host of most savannah tsetse fly species [9]. The targets are usually impregnated with biodegradable deltamethrin pyrethroid insecticide that effectively kills or knocks-down the flies when they land on the target. This technology has been used successfully to control tsetse flies in environmentally sensitive and diverse forest, riverine and savannah ecosystems due to the relatively high specificity and minimal environmental contamination components [10].
On the other hand, technology of spatial odor repellent of tsetse flies has recently been devised [11,12]. The technology deters tsetse flies from interacting with vertebrate hosts of the flies, such as humans and their livestock, effectively impeding transmission of trypanosomes through infective bites from the flies. This technology complements the bait technology above, by establishing a formidable “push (repellent) – pull (attractant)” technology that can rapidly suppress tsetse fly populations and associated transmission of HAT and AAT causing trypanosomes. Formulation of the spatial repellents was informed by tsetse fly’s (Glossina pallidipes or Glossina morsitans morsitans) active avoidance behaviors of waterbuck bovid (Kobus ellipsiprymnus defassa) vertebrate animals [9,13]. Assessments of chemical composition of the allomones emitted by the waterbuck revealed guaiacol, geranylacetone, pentanoic acid and δ-octalactone as the key components [14]. These components were formulated and field optimized into a Waterbuck Repellent Compound (WRC) blend, comprising of 2:1:3:3 blend of guaiacol, geranylacetone, pentanoic acid and δ-octalactone respectively for effective field repellence of G. pallidipes [12]. This formulation provided substantial protection to livestock from tsetse fly transmitted trypanosomes [15]. Follow-up studies on the effects of structural variants (analogs) of δ-octalactone on olfactory/behavioral responses of G. pallidipes or G. m. morsitans established that replacing δ-octalactone with its δ-nonalactone analog enhanced repellency to both species [16], and combining δ-nonalactone, heptanoic acid, 4-ethyl guaiacol and geranyl acetone in a 6:4:2:1 proportion generated a Novel Repellent Blend (NRB) effective against both tsetse fly species [11]. However, relative efficacy of these repellents (WRC and NRB) against these tsetse fly species and G. f. fuscipes, an important vector of HAT in eastern and central Africa, is poorly understood. Such information can inform choice of appropriate repellent for more efficacious protection of humans and their livestock from tsetse fly bites and associated trypanosomiasis transmission. This study was initiated to establish the relative field repellence of G. pallidipes and G. f. fuscipes by WRC and NRB in Shimba Hills National Reserve and Ndere Island National Park where G. pallidipes and G. f. fuscipes respectively are naturally abundant.
Materials and methods
Study areas
The studies were conducted at Shimba Hill National Reserve (-4°15′26″S, 39°23′16″E; altitude 403 m) in Kwale County along the coast, and Ndere Island National Park (-0°12′22″S, 34°30′44″E; altitude 1,228 m) along the shores of Lake Victoria, in western Kenya. Tsetse fly species in the Shimba Hill National Reserve include G. pallidipes, G. austeni and G. brevipalpis while G. f. fuscipes is the only tsetse fly species inhabiting the Ndere Island National Park. The Shimba Hill National Reserve is about 300 Km^2^ and is inhabited by sable antelopes, buffalos, syke monkeys, elephants, giraffes, leopards, genets, waterbucks, African bush babies, civet cats, hyenas, bush pigs, coastal black and white colobus monkeys, red, blue and bush duikers, greater galagoes, black-faced vervet monkeys, bushbucks, serval cats, black and red shrews, knob-bristled suni shrew and many more. The vegetation in the reserve consists mostly of coastal rain forest and semi-evergreen woodland and grassland. The reserve region experiences long and short rainy seasons from April to June, and October to November, respectively, with 855 mm −1682 mm annual rainfall, mean annual temperature of 24.2 °C, with the highest daily temperatures of 33°C in February-March and November, and lowest temperature of 21 °C in July-August. The Ndere Island National Park is a 4.2 km^2^ island and supports a variety of animals that include hippos, monitor lizards, Nile crocodiles, several fish species, snakes, baboons, impalas, the rare sitatunga antelopes, water bucks, zebras and warthogs among others. The island is inhabited by grassland and shrubs, indigenous trees and woodland forest along the shorelines of the lake. The park features a tropical rainforest climate with no true dry season and significant rainfall year-round. Annual rainfall is about 276.22 mm. Mean annual temperature is 22.9°C, with the highest daily temperatures of 30 °C. Approvals to perform experiments in the protected areas were obtained from the Kenya Wildlife Service (KWS)(permit No. KWS/BRM/5001), National Commission for Science, Technology & Innovation (NACOSTI) (permit No. NACOSTI/P/18/28381/22226), and the National Environment Management Authority (NEMA) (permit No. NEMA/AGR/93/2018) in Kenya.
Test chemical compounds and blends
We sourced for pure (98–99%) δ-nonalactone, δ-octalactone, geranyl acetone, acetone, heptanoic acid, pentanoic acid, 4-ethyl guaiacol, 1-octen-3-ol, 3-n-propylphenol and p-cresol (4-methylphenol) from Sigma-Aldrich, Taufkirchen, Germany. For experiments, we separately formulated 1) WRC blend comprising of guaiacol, geranylacetone, pentanoic acid and δ-octalactone (2:1:3:3) (Saini et al., 2017), 2) NRB comprising of δ-nonalactone, heptanoic acid, 4-ethyl guaiacol and geranyl acetone (6:4:2:1) [11] and 3) POCA, comprising of 3-n-propylphenol (P), octanol (O) and p-cresol (C) (1:4:8), which together with separately released acetone (A), form an established odor blend attractant of G. pallidipes [17,18].
Experimental design
We employed randomized Latin Square Design (LSD) experimental approach in assessment of field responses/catches of adult G. pallidipes or G. f. fuscipes to the treatments. The responses of G. pallidipes and G. f. fuscipes were assessed in Shimba Hills National Reserve and Ndere Island National Park, respectively. In assessing the responses of G. pallidipes to NRB or WRC, a series of NG2G traps [19] were deployed in the field, at intervals of 300 m to minimize spatial interaction between treatment effects in an LSD experiment comprising of four traps (treatments) in four sites randomly rotated within four days, such that each treatment was deployed on each site once as we previously described [11,16,20]. Treatments comprised of the trap with or without 1) odor treatment (no-odor control) 2) POCA, 3) POCA with WRC or 4) POCA with NRB in three independent replicates, conducted in three separate independent blocks 1–2 km apart within the study area. Consequently, we deployed nine (3X3) independent replicates of each treatment. We dispensed 1) 3-n-propylphenol, octanol and p-cresol (POC), WRC or NRB using sealed thin-walled polythene sachets constructed from polyethylene lay flat tubing (150 microns thick) folded to form a surface area of about 50 cm^2^ [11] and acetone (A) using 28 ml clear glass universal bottles. The sachets were pegged 25 cm above the ground and 30 cm downwind from the center of respective traps, and trapped flies were collected at 24-hr intervals, at 5:00 pm each day, to encompass morning and afternoon bimodal G. pallidipes activity peaks [21]. The flies were then identified to species level using standard taxonomic keys as outlined in Pollock [22]. We established the release rates of the odors and documented catches (counts) of the flies (G. pallidipes) by sex, treatment, site (trap), day and block. Mean release rates for WRC and NRB were established based on daily loss in masses, pre- vs post- deployment, as 6.45 ± 0.14 and 6.32 ± 0.09 mg/h respectively. POCA bait with two componentsPOC and Acetone had mean release rates of 7.89 ± 1.02 and 495.35 ± 6.11 mg/h, respectively.
We assessed responses of G. f. fuscipes to WRC or NRB using the same approach we used in assessing responses to G. pallidipes as described above. However, POCA was excluded from the treatments since G. f. fuscipes responses are not influenced (non-responsive to) by POCA [23,24]. This effectively reduced the experimental design to three treatments (no-odor control, WRC and NRB) for G. f. fuscipes. Additionally, we replaced the NG2G traps [19] with targets (with sticky surface) that have been demonstrated to have better efficacy in trapping G. f. fuscipes [25]). Consequently, the LSD experiment comprised of three targets (treatments) in three sites randomly rotated within three days.
Data analysis
We assessed for differences in patterns of catches between sexes of G. pallidipes or G. f. fuscipes tsetse flies using Chi-Square, and where the patterns of catches did not significantly differ between the sexes, we pooled data for both sexes for Analysis of Variance (ANOVA), or separately analyzed the data, using ANOVA, where the differences were significant. For ANOVA, we first transformed counts of tsetse fly catches for each treatment and replicate to log (n + 1) to normalize the distributions and homogenize the variances. We used general linear model one-way ANOVA for LSD data with transformed catches as response variable, treatments as factors and sites/days as blocks. We separated the means by Tukey’s Honestly Significant Difference (HSD) post hoc test and back-transformed the data to antilog-1 for reporting the geometric means (± 95% Confidence Intervals) [26]. We obtained catch indices for G. pallidipes or G. f. fuscipes for different treatments by expressing the response of each treatment category as a proportion of responses to control without odor treatment. We ensured that the transformed data met the normal distribution assumption requirement for t test using Shapiro-Wilk test, and homogeneity of variance requirement for ANOVA using Levene test. Additionally, we assessed for additivity of variance by evaluating the interactions between sites, days and treatments. We summarized descriptive statistics, where necessary, and presented them as means with their ± 95% Confidence Intervals. Analyses were performed using R [27] and GraphPad Prism version 10.0.0 for Mac [28].
Results
We have summarized the data on abundance of arthropods captured in the traps and targets in Shimba Hills National Reserve and Ndere Island National Park in Fig 1. Tsetse flies were most abundant, constituting 97 and 64% of arthropods catches in the reserve and park, respectively. Among the tsetse flies, G. pallidipes, G. austeni and G. brevipalpis were caught in traps in Shimba Hills National Reserve, where G. pallidipes was the most abundant, while only G. f. fuscipes were captured in targets in Ndere Island National Park. The pattern of catches between male and female G. pallidipes to the treatments was significantly different (X^2^ (3, N = 8) = 17.12, p < 0.001). On the other hand, pattern of catches between sexes in G. f. fuscipes responses to the treatments were similar (X^2^ (2, N = 6) = 2.70, p = 0.2594). Consequently, ANOVA for male and female G. pallidipes responses to treatments were conducted separately, whereas data on responses of male and female G. f. fuscipes to the treatments were pooled for the analyses. Overall, indices of responses of male G. pallidipes to WRC and NRB, relative to no-odor control, were 1.4 and 0.3 respectively, while the indices for responses of the females to the same were 1.0 and 0.3 respectively (S1 Table). On the other hand, indices of responses of G. f. fuscipes to WRC and NRB were 0.8 and 0.9 respectively (S1 Table). Consequently, both sexes of G. pallidipes were most and least attracted to traps baited with POCA and POCA+NRB blends respectively (Fig 2A). Responses of both sexes of G. pallidipes in the traps baited with WRC and control traps (without odor) were similar (p > 0.05). However, G. pallidipes responses to traps baited with NRB were significantly less than those responding to traps baited with WRC, where male and female G. pallidipes’ responses to traps baited with NRB were 2.0 (F_(3, 47)_ = 18.8015, p < 0.001) and 1.6 (F_(3, 47)_ = 12.0391, p < 0.001) folds less respectively, than the flies responding to WRC (Fig 2A, S1 Table). Responses of G. f. fuscipes to the traps with WRC, NRB or control (un-baited) were similar (F (2, 26) = 0.1227, p = 0.8855) (Fig 2B, S1 Table).
Distribution of catches of arthropods in traps and targets in Shimba Hills National Reserve and Ndere National Park respectively, in Kenya.
Geometric mean catches of G. pallidipes and G. f. fuscipes in traps and targets with various odor treatments in Shimba Hills National Reserve and Ndere National Park respectively, in Kenya.
Discussion
Humans and their livestock can be protected from infectious and/or nuisance bites from arthropod by avoiding arthropod habitats or by using arthropod behavior modifying repellents active against target arthropod(s). Repellent technologies constitute a fundamental approach for prevention of transmission of arthropod-borne diseases, with spatial repellents (spatial emanators) augmenting contact repellents and pesticides within an integrated vector management framework at minimal cost. The spatial repellents can also be deployed alone to potentially protect humans and their livestock from the vectors, especially in settings with limited coverage by other vector control interventions. Most spatial repellents have been developed for and deployed against mosquito-borne diseases [29]. Development of spatial repellents against tsetse flies is fairly recent, with only Waterbuck Repellent Compound (WRC) [15] and Novel Repellent Blend (NRB) [11] being described to date for potential protection of humans and their livestock. Our findings in the present study reveal 3.5 and 4.7 -folds better repellence (mean indices of fly catches in WRC relative to NRB baited traps) of female and male G. pallidipes, respectively, by NRB relative to WRC. These remarkable observations in behavioral responses of the flies may be due to differences in their olfactory perception of the repellents, suggesting that NRB can potentially provide enhanced protection of humans and their livestock from G. pallidipes bites and associated trypanosomiasis (HAT/AAT) transmission and nuisance. The WRC exhibits >80% efficacy in protection of cattle against G. pallidipes [15]. Consequently, NRB is expected to exhibit near absolute efficacy against G. pallidipes, based on the current comparative performance against WRC, which should be empirically established. The absence of efficacy of either repellents against G. f. fuscipes, suggests that NRB and WRC may have a narrow range of tsetse fly species specificity and hence necessitate a need for expanded search for repellents active against G. f. fuscipes. Additionally, extent of efficacy or lack thereof of the repellents within morsitans/savannah and palpalis/riverine group, of flies, to which G. pallidipes and G. f. fuscipes respectively belong remains to be determined. Such group-wide responses have been observed in relative efficacy of POCA attractant against morsitans and palpalis, with POCA being effective and ineffective against the former and latter, respectively [17,18,30]. Additionally, these findings suggest that protection of vertebrate hosts by either repellents would be sufficient or partial in the field, where G. pallidipes is the only species or sympatric with G. f. fuscipes, respectively. Unfortunately, G. pallidipes is sympatric with other savannah species and/or palpalis group of tsetse flies in most of their habitats in sub-Sahara Africa, suggesting a need to: 1) assess efficacy of either repellents against other tsetse fly species such as Glossina austeni, Glossina swynnertoni, Glossina morsitans centralis that are typically sympatric with G. pallidipes, or 2) prospect for and formulate effective novel repellents against G. f. fuscipes.
Effective administration of the NRB for human and livestock protection will require development and optimization of a novel, needs specific, delivery system for the repellent. The current delivery system employs sealed thin-walled polythene sachets constructed from polyethylene lay-flat tubing adapted from delivery systems designed for dispensation of POC components of POCA [31] on stationary traps and targets. The NRB on the other hand should ideally be dispensed from moving subjects (hosts) and at a rate sufficiently controlled to extend the utility/longevity of the product. The waterbuck-like smell associated with these repellents can also be masked with a pleasant smell to enhance acceptance for use within proximity of vertebrates, including humans. The repellent can potentially be integrated into the woven fabric and nets to broaden its utility. So far, NRB has successfully been micro-encapsulated in beta cyclodextrin nano-particles to moderate release of the repellent [32]. However, field performance of the encapsulated product in repellence of the flies remains to be determined.
In conclusion, NRB appears to be several-fold more repellent than WRC against G. pallidipes. The impact of this enhanced repellence on livestock protection remains to be determined. Both NRB and WRC are ineffective against G. f. fuscipes, suggesting possible tsetse fly species specificity among the repellents and the need to determine a spectrum of tsetse fly species against which these repellents are efficacious. Development and optimization of a novel delivery system for NRB can partially expand its utility and application in vector control.
Supporting information
S1 TableAnalysis of variance of responses of G. pallidipes and G. f. fuscipes to various odor treatments in Shimba Hills National Reserve and Ndere National Park respectively, in Kenya.(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Brun R, Blum J, Chappuis F, Burri C. Human African trypanosomiasis. Lancet. 2010;375(9709):148–59. doi: 10.1016/S 0140-6736(09)60829-1 19833383 · doi ↗ · pubmed ↗
- 2Hotez PJ, Fenwick A, Savioli L, Molyneux DH. Rescuing the bottom billion through control of neglected tropical diseases. Lancet. 2009;373(9674):1570–5. doi: 10.1016/S 0140-6736(09)60233-6 19410718 · doi ↗ · pubmed ↗
- 3Vreysen MJB, Seck MT, Sall B, Bouyer J. Tsetse flies: their biology and control using area-wide integrated pest management approaches. J Invertebr Pathol. 2013;112(Suppl):S 15-25. doi: 10.1016/j.jip.2012.07.026 22878217 · doi ↗ · pubmed ↗
- 4Vale GA, Mutika G, Lovemore DF. Insecticide-treated cattle for controlling tsetse flies (Diptera: Glossinidae): some questions answered, many posed. Bull Entomol Res. 1999;89:569–78.
- 5Torr SJ, Maudlin I, Vale GA. Less is more: restricted application of insecticide to cattle to improve the cost and efficacy of tsetse control. Med Vet Entomol. 2007;21(1):53–64. doi: 10.1111/j.1365-2915.2006.00657.x 17373947 · doi ↗ · pubmed ↗
- 6Lord JS, Lea RS, Allan FK, Byamungu M, Hall DR, Lingley J, et al. Assessing the effect of insecticide-treated cattle on tsetse abundance and trypanosome transmission at the wildlife-livestock interface in Serengeti, Tanzania. P Lo S Negl Trop Dis. 2020;14:e 0008288.10.1371/journal.pntd.0008288 PMC 747352532841229 · doi ↗ · pubmed ↗
- 7Dangerfield JM. Rapid assessment of widespread aerial spraying of deltamethrin for tsetse fly control on non-target insects in the Okavango Delta, Botswana. In: FAO/IAEA international conference on area-wide control of insect pests: Integrating the sterile insect and related nuclear and other techniques. Book of extended synopses. 2005: 73.
- 8Vale GA. Development of baits for tsetse flies (Diptera: Glossinidae) in Zimbabwe. J Med Entomol. 1993;30(5):831–42. doi: 10.1093/jmedent/30.5.831 8254629 · doi ↗ · pubmed ↗