Novel secondary metabolite from a new species of Hypoxylon saxatilis sp. nov. for suppressing bacterial wilt in tomato
Thanapat Suebrasri, Wasan Seemakram, Awat Wisetsai, Thanawan Gateta, Sutarin Preepram, Phornnapa Saentao, Sophon Boonlue

TL;DR
A new fungus produces tetrahydrofuran, a natural compound that effectively fights tomato wilt disease caused by Ralstonia solanacearum.
Contribution
Discovery of tetrahydrofuran as a novel natural secondary metabolite with bactericidal activity against Ralstonia solanacearum.
Findings
Hypoxylon saxatilis produces tetrahydrofuran, a previously unknown natural secondary metabolite.
Tetrahydrofuran inhibits Ralstonia solanacearum and damages its cell wall structure.
The compound reduced tomato wilt disease severity by up to 83.33% under greenhouse conditions.
Abstract
•We reported a new endophytic species with bactericidal activity against Ralstonia solanacearum.•We elucidated that tetrahydrofuran is produced by the newly identified endophytic fungus Hypoxylon saxatilis.•We were the first to identify tetrahydrofuran as a previously undescribed secondary metabolite from natural sources.•We were the first to report tetrahydrofuran bactericidal compounds effective against R. solanacearum. We reported a new endophytic species with bactericidal activity against Ralstonia solanacearum. We elucidated that tetrahydrofuran is produced by the newly identified endophytic fungus Hypoxylon saxatilis. We were the first to identify tetrahydrofuran as a previously undescribed secondary metabolite from natural sources. We were the first to report tetrahydrofuran bactericidal compounds effective against R. solanacearum. The bacterium Ralstonia solanacearum is an…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 2
Figure 2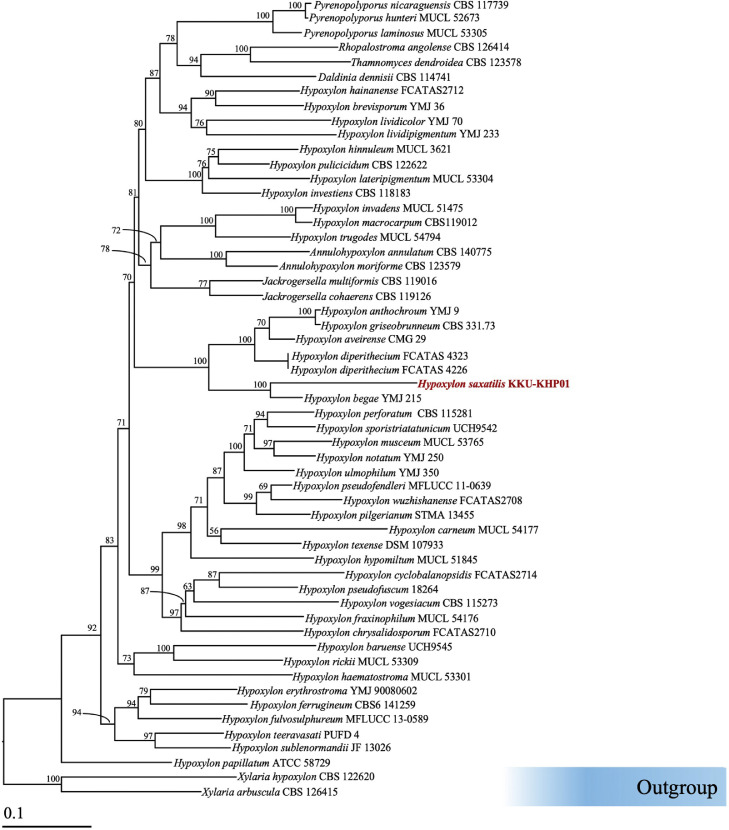 Figure 3
Figure 3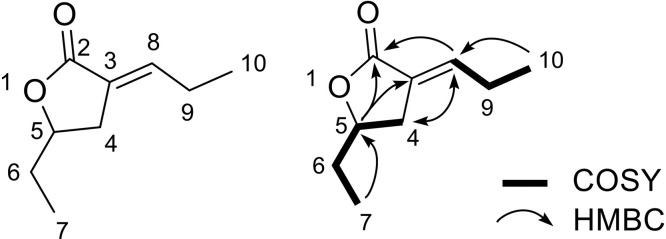 Figure 4
Figure 4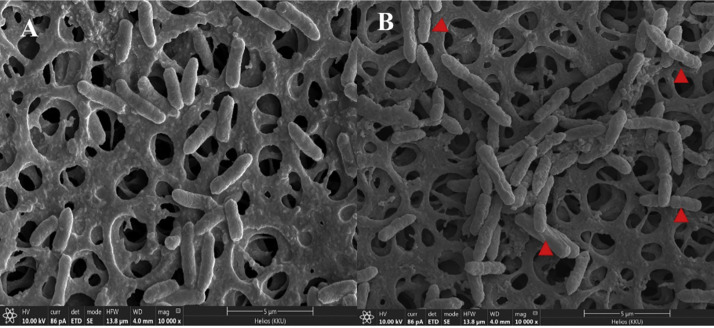 Figure 5
Figure 5 Figure 6
Figure 6Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant-Microbe Interactions and Immunity · Plant Pathogenic Bacteria Studies · Plant Pathogens and Fungal Diseases
Introduction
1
Tomato (Lycopersicon esculentum Mill.) is a fruit widely consumed worldwide. It is popular for use in various types of dishes, such as tomato sauce, salads, and soups, and as ingredients in many other foods, both fresh and processed (Liu et al., 2024). It is often produced on an industrial scale to make products that are used in daily life. In 2024, global consumption of tomatoes, including both fresh and processed forms, was estimated to have reached approximately 45.7 million tons, with the majority coming from processed products such as tomato sauce and tomato juice (World Processing Tomato Council, 2024). the main areas for the cultivation of tomatoes in Thailand are in the northern and northeastern regions. According to annual tomato cultivation statistics from the Northeast Region of the Department of Agricultural Extension, the area for tomato production increased from 2490.24 hectares in 2019 to 6328.8 hectares in 2020 (Office of Agricultural Economics, 2020). However, tomatoes are invaded by pests and diseases caused by microorganisms, which can lead to reduced yields, especially bacterial wilt diseases caused by Ralstonia solanacearum. It is one of the most significant plant pathogens worldwide, infecting over 450 host species. It colonizes the xylem vessels of various plants, leading to bacterial wilt disease. Its host range encompasses numerous plant species, including tobacco, bananas, peppers, potatoes, geraniums, and especially tomatoes (Vailleau and Genin., 2023). Bacterial wilt is prevalent in tropical, subtropical, and temperate regions, causing substantial economic losses due to yield reduction and the costs associated with disease management (Wicker et al., 2007).
Several chemicals, such as copper-based bactericides, copper hydroxide, copper oxychloride (Boucher et al., 2005), and sodium hypochlorite (Leelaporn et al., 2008), have been used to control bacterial wilt caused by R. solanacearum. Chemicals can be used to eliminate bacterial wilt disease. The application of these chemicals must be approached with caution, as improper use or excessive quantities can lead to adverse effects on both the environment and the health of consumers and farmers. Currently, biocontrol is becoming increasingly popular because of its safety for consumers and its environmentally friendly nature. Endophytic microbes are another promising alternative for controlling plant pathogens, as they reside within plant tissues without harming the host (Chauhan et al., 2024). Furthermore, they can promote plant growth and produce secondary metabolites that inhibit the growth of pathogens (Kloepper et al., 2018; Singh et al., 2020). In addition, endophytic fungi isolated from medicinal plants are selected over those from diseased host plants due to the well-documented diversity of endophytes in medicinal plants capable of producing bioactive secondary metabolites. These endophytic fungi often possess antimicrobial properties that enhance the natural defense mechanisms of their host plants (Strobel and Daisy, 2003). Therefore, medicinal plants represent a promising source for isolating endophytic fungi with potential applications in the development of novel biocontrol agents for plant disease management. Previous reports have indicated that the endophytic bacteria Bacillus subtilis (VSB-1) and Streptomyces thermodiastaticus (ORA-1) are effective against bacterial wilt under pot culture conditions (Agarwal et al., 2021). The endophytic root microbiomes, including Pseudomonas simiae E18, Bacillus amyloliquefaciens E60, and Paenibacillus polymyxa E29, are also highly effective against R. solanacearum (Zhang et al., 2022). In addition, important natural bactericidal compounds, such as 2-azaanthraquinone derivatives produced by the endophytic fungus Lophiostoma sp. (Mao et al., 2021) and lipopeptide secreted from Bacillus subtilis R31, effectively inhibit the growth of R. solanacearum (Li et al., 2023). However, the role of endophytic fungi and their secondary metabolites in controlling bacterial wilt in tomatoes caused by R. solanacearum is poorly understood. Moreover, the secondary metabolites that exhibit efficacy in reducing bacterial wilt incidence hold promising potential for future commercial application. Therefore, the present work aimed to isolate and identify endophytic fungi from medicinal plants and determine the ability of endophytic fungi to produce bioactive compounds for controlling R. solanacearum. Furthermore, the bioactive compounds isolated from endophytic fungi were subjected to structural elucidation. Their potential to protect tomato plants from R. solanacearum infection was evaluated under greenhouse conditions. The findings suggest that these compounds hold promise as biological control agents against bacterial wilt, offering a sustainable approach to enhance tomato yield in the future.
Materials and methods
2
Collection of plant materials
2.1
A total of twenty medicinal plants were collected from the mountain in Lao Hai Ngam Subdistrict, Kuchinarai District, Kalasin Province, Thailand (16°45′06.3″N 104°22′00.2″E). The plant samples were stored in a sterilized plastic bag at 4 °C until they were processed.
Isolation of endophytic fungi
2.2
The plant samples were cleaned with tap water to remove dust. Then, they were cut into pieces with a size of 1–2 mm. The plant segments were sterilized by first soaking them in 70 % (v/v) ethanol for 3 min, followed by immersion in 5 % (v/v) sodium hypochlorite for 1 min. Afterward, the segments were rewashed with 70 % (v/v) ethanol for 3 min, rinsed three times with sterile distilled water, and dried on sterile paper under airflow to ensure complete drying. The sterilized samples were then placed onto half-strength potato dextrose agar (PDA) plates supplemented with 2000 mg/L chlortetracycline to inhibit the growth of endophytic bacteria. Fungi growing from the segment samples were subcultured onto fresh PDA media until a pure culture was obtained. Finally, to confirm the absence of contamination on the sterilized plant segments, an aliquot of the sterilizing solution was placed on agar plates and incubated for 2 weeks to ensure that no fungal growth occurred (Suebrasri et al., 2020).
Screening endophytic fungi as biological control agents against R. solanacearum
2.3
The endophytic fungi were screened for their ability to antagonize the pathogenic bacteria R. solanacearum using the agar plug diffusion method. The bacteria were cultivated in nutrient broth and shaken at 180 rpm at 37 °C for 24 h. After incubation, the turbidity of the bacterial culture was adjusted to 0.5 McFarland, and the bacteria were spread on nutrient agar. Five-day-old fungal endophytes grown on potato dextrose agar media were plugged and placed on the surface of nutrient agar plates, which were spread with the bacteria R. solanacearum. A clear zone around the colony of fungal endophytes was observed after incubation at 37 °C for 5 d
Fungal endophyte identification
2.4
Fungal endophyte isolates capable of inhibiting the bacterial pathogen R. solanacearum were initially identified based on their morphology using light microscopy. Additionally, the fungal colony morphology was observed by culturing on an oatmeal agar plate. To confirm fungal identification, gene sequencing targeting universal primers, including the internal transcribed spacer regions (ITS1–4) and β-tubulin 2 (TUB2), was performed. The primer sequences for the ITS gene and β-tubulin 2 gene were as follows: ITS1 (5′-TCC GTA GGT GAA CCT GCG G-3′), ITS4 (5′-TCC TCC GCT TAT TGA TAT GC-3′), Btub2 (5′-GTB CAC CTY CAR ACC GGY CAR TG-3′), and Btub4 (5′-CCR GAY TGR CCR AAR ACR AAG TTG TC-3′). PCRs were carried out in a total volume of 25 µL, consisting of 5 µL of 10 × PCR buffer, 1.5 µL of 25 mM MgCl₂, 0.5 µL of 1.25 mM dNTPs, 1.5 µL of each primer (150 µM), 100 ng of fungal DNA template, and 0.2 µL of Taq DNA polymerase (5 U/µL) (Thermo Fisher Scientific, Lithuania, EU). The volume was adjusted to 25 µL with sterile distilled water. PCR amplification was performed using a thermal cycler under the following conditions: initial denaturation at 95 °C for 5 min; 35 cycles of denaturation at 95 °C for 1 min, annealing at 52 °C for 1 min, and extension at 72 °C for 90 s; and a final extension at 72 °C for 10 min. The PCR products were purified using the GeneJET™ PCR Purification Kit (Fermentas, Canada). The PCR products were subsequently sequenced with the 1st BASE DNA Kit (Malaysia). Subsequently, the nucleotide sequences were analyzed using the BLAST tool available on the NCBI website.
The authentic DNA sequences from GenBank (http://www.ncbi.nlm.nih.gov) and those obtained from the PCR products were combined with the novel sequences generated in this study, as shown in Table 1. Xylaria hypoxylon CBS 122,620 and X. arbuscula CBS 126,415 were used as the outgroup. Multiple sequence alignment was performed using MUSCLE (Edgar, 2004) with default settings, and adjustments were made as needed using BioEdit version 6.0.7 (Hall, 2013). The combined alignment of the ITS and β-Tubulin sequences was deposited in TreeBASE.Table 1. Details of the sequences used in the molecular phylogenetic analyses in this study. The samples and sequences obtained in this study are in bold.Table 1. Hypoxylon speciesStrain/IsolateCountryGenBank accession numberReferencesITSβ-Tubulin**Annulohypoxylon annulatumCBS 140,775USAKU604559KX376353Kuhnert et al. (2017)A. moriformeCBS 123,579MartiniqueKX376321KX271261Kuhnert et al. (2017)Daldinia dennisiiCBS 114,741AustraliaJX658477KC977262Stadler et al. (2014)Hypoxylon anthochroumYMJ 9MexicoJN660819AY951703Hsieh et al. (2005)H. aveirenseCMG 29 TPortugalMN053021MN066636Vicente et al. (2021)H. baruenseUCH9545 TPanamaMN056428MK908142Cedeño-Sanchez et al. (2020)H. begaeYMJ 215USAJN660820AY951704Hsieh et al. (2005)H. brevisporumYMJ 36Puerto RicoJN660821AY951705Hsieh et al. (2005)H. carneumMUCL 54,177FranceKY610400KX271270Wendt et al. (2018)H. chrysalidosporumFCATAS2710 TChinaOL467294OL584229Ma et al. (2022)H. cyclobalanopsidisFCATAS2714 TChinaOL467298OL584232Ma et al. (2022)H. diperitheciumFCATAS 4226 TChinaON178671ON365565Zhu et al. (2024)H. diperitheciumFCATAS 4323ChinaON178672ON365566Zhu et al. (2024)H. erythrostromaYMJ 90,080,602ChinaJN979416AY951716Hsieh et al. (2005)H. ferrugineumCBS6 141,259AustriaKX090079KX090080Friebes G & Wendelin I (2016)H. fraxinophilumMUCL 54,176FranceKC968938KC977301Kuhnert et al. (2014)H. fulvosulphureumMFLUCC 13–0589 TThailandKP401576KP401584Sir et al. (2015)H. griseobrunneumCBS 331.73 TIndiaKY610402KC977303Kuhnert et al. (2014)H. hematostromaMUCL 53,301 ETMartiniqueKC968911KC977291Wendt et al. (2018)H. hainanenseFCATAS2712 TChinaOL467296OL584231Ma et al. (2022)H. hinnuleumMUCL 3621 TMUCL 3621MK287537MK287575Daranagama et al. (2015)H. hypomiltumMUCL 51,845GuadeloupeKY610403KX271249Wendt et al. (2018)H. invadensMUCL 51,475 TFranceMT809133MT813038Becker et al. (2020)H. investiensCBS 118,183MalaysiaKC968925KC977270Wendt et al. (2018)H. lateripigmentumMUCL 53,304 TMartiniqueKC968933KC977290Wendt et al. (2018)H. lividicolorYMJ 70ChinaJN979432AY951734Helaly et al. (2018)H. lividipigmentumYMJ 233MexicoJN979433AY951735Hsieh et al. (2005)H. macrocarpumCBS119012GermanyON792785ON813071Helaly et al. (2018)H. musceumMUCL 53,765GuadeloupeKC968926KC977280Kuhnert et al. (2014)H. notatumYMJ 250USAJQ009305AY951739Hsieh et al. (2005)H. papillatumATCC 58729TUSANR155153KC977258Wendt et al. (2018)H. perforatumCBS 115,281FranceKY610391KX271250Wendt et al. (2018)H. pilgerianumSTMA 13,455MartiniqueKY610412KY624315Wendt et al. (2018)H. pseudofendleriMFLUCC 11–0639ThailandKU940156N/A(Dai et al. (2017)H. pseudofuscum18,264 TGermanyMW367857MW373867Lambert et al. (2021)H. pulicicidumCBS 122622TMartiniqueJX183075JX183072Bills et al. (2012)H. rickiiMUCL 53,309 ETMartiniqueKC968932KC977288Wendt et al. (2018)H. saxatilisKKU-KHP01ThailandLC872603LC872604This studyH. sporistriatatunicumUCH9542TPanamaMN056426MK908140Cedeño-Sanchez et al. (2020)H. sublenormandiiJF 13,026Sri LankaKM610291KM610303Kuhnert et al. (2015)H. teeravasatiPUFD 4TIndiaN/AMG986894Phookamsak et al. (2019)H. texenseDSM 107933TUSAMK287536MK287574Sir et al. (2019)H. trugodesMUCL 54794TSri LankaKF234422KF300548Kuhnert et al. (2014)H. ulmophilumYMJ 350RussiaJQ009320AY951760Hsieh et al. (2005)H. vogesiacumCBS 115,273FranceKC968920KX271275Wendt et al. (2018)H.wuzhishanenseFCATAS2708TChinaOL467292OL584227Ma et al. (2022)Jackrogersella cohaerensCBS 119,126GermanyKY610396KY624314Wendt et al. (2018)J. multiformisCBS 119,016 ETGermanyKC477234KX271262Kuhnert et al. (2014)Pyrenopolyporus hunteriMUCL 52,673 ETIvory CoastKY610421KU159530Wendt et al. (2018)Py.laminosusMUCL 53305TMartiniqueKC968934KC977292Wendt et al. (2018)Py. nicaraguensisCBS 117,739Burkina FasoAM749922KC977272Wendt et al. (2018)Rhopalostroma angolenseCBS 126,414Ivory CoasKY610420KX271277Kuhnert et al. (2014)Thamnomyces dendroideaCBS 123,578 TFrench GuianaFN428831KY624313Stadler et al. (2014)X. arbusculaCBS 126,415GermanyKY610394KX271257Wendt et al. (2018)
Phylogenetic analysis was conducted using a combined dataset of ITS and β-tubulin sequences. A phylogenetic tree was constructed using both the maximum likelihood (ML) and Bayesian inference (BI) methods. An ML analysis was performed with 25 rate categories and 1000 bootstrap (BS) replicates under the GTRCAT model of nucleotide substitution, using RAxML-HPC2 version 8.2.12 (Felsenstein, 1985, Stamatakis, 2006) via the CIPRES web portal. For the BI analysis, the best-fit substitution models were determined using the Akaike information criterion (AIC) in jModelTest version 2.1.10 (Darriba et al., 2012): GTR for ITS and β-Tubulin. BI was carried out using MrBayes version 3.2.6 (Ronquist et al., 2012), with six simultaneous Markov chains run for one million generations, starting from random trees and sampling every 1000 generations. The first 20 % of trees, representing the burn-in phase, were discarded. The remaining trees were used to calculate posterior probabilities (PPs) in a majority rule consensus tree. The resulting phylogenetic trees were visualized using FigTree version 1.4.0 (Rambaut, 2012).
Extraction and purification of bioactive compounds from fungal endophytes
2.5
The fungal endophytes were cultured on potato dextrose agar (PDA) at 30 °C for 7 days. Next, 0.5 cm mycelial plugs (using a No. 5 cork borer) from 250 plugs were inoculated into 5000 mL Erlenmeyer flasks containing 2500 mL of potato dextrose broth (PDB) and incubated at room temperature for 30 days under static conditions. After 30 days, the fungal culture was filtered to remove the mycelium. The mycelium was then dried in a hot air oven at 55 °C for 5 days. The filtered broth and dried mycelium were subsequently extracted twice with an equal volume of ethyl acetate. The organic phases were evaporated to dryness at 40 °C using a rotary vacuum evaporator under reduced pressure. The fungal crude extract was then weighed. The crude extract of each endophytic fungal isolate was subsequently dissolved in dimethyl sulfoxide (DMSO) and then diluted with sterile distilled water to a concentration of 20 mg/mL. The effects of the fungal crude extracts on the growth of R. solanacearum were tested using the disc diffusion method. The crude ethyl acetate extract of the endophytic fungi from Compound 1 was subjected to repeated separations using column chromatography with an appropriate stationary phase and solvent system, as previously determined using thin layer chromatography (TLC). The crude extracts were then purified using silica gel column chromatography and eluted with a gradient of ethyl acetate and n-hexane or an isocratic system to obtain fractions using TLC analysis (Suebrasri et al., 2020).
Structure elucidation of the bioactive compound
2.6
Nuclear magnetic resonance (NMR) and mass spectrometry were used to predict the chemical structure of the isolated antifungal compounds. The compounds were analyzed using ¹H and ¹³C NMR spectra obtained on a Varian Mercury Plus 400 spectrometer. Chemical shifts are reported in δ (ppm), with CDCl₃ or CD₃OD as the solvent and residual CHCl₃ as the internal standard. The ¹H and ¹³C NMR data were recorded, detailing the number of protons, multiplicity, and coupling constants (J in Hz) (Suebrasri et al., 2020).
SEM analysis of the effects of the compounds on bacterial cells
2.7
A scanning electron microscope (SEM) was used to detect changes on the bacterial cell surface. Therefore, the effects of the compounds on the cell structure of the bacterial pathogen R. solanacearum were observed via SEM. To prepare the sample, the bacteria grown in nutrient broth for 18 h were centrifuged at 12,000 rpm and 4 °C for 2 min. The bacterial cells were washed with phosphate-buffered saline solution twice. The bacterial cell turbidity at 0.08–0.10 (10^6^ CFU/mL) was subsequently adjusted using a spectrophotometer at 600 nm. The 50 μL of the bacterial mixture was transferred to a microcentrifuge tube with 50 μL of the compound at 1 μg/mL, which was dissolved in 10 % DMSO. The mixture was incubated at 37 °C for 1 hour. After incubation, the solution was dropped on a nuclear pore polycarbonate membrane and fixed with 2.5 % glutaraldehyde overnight at 4 °C. Subsequently, the samples were dehydrated through a series of ethanol dilutions (50, 60, 70, 80, 90, 95, and 100 %) for 10 min at each gradient. The samples were placed onto copper stubs and coated with gold prior to observation using a scanning electron microscope (SEM) (LEO, Electron Microscopy Ltd., Cambridge, UK).
Disease suppression assay in tomato
2.8
The pot experiment was conducted in a greenhouse at the Department of Microbiology, Faculty of Science, Khon Kaen University, Khon Kaen, Thailand. The sandy loam soil used in the experiment had the following chemical properties: pH 7.26, electrical conductivity (EC) of 0.043 dS/m, 6.4 g kg⁻¹ of soil organic matter, 240 mg kg⁻¹ of total nitrogen (N) (with a soil C/N ratio of approximately 13), 146 mg kg⁻¹ total of phosphorus (P), 428 mg kg⁻¹ of extractable potassium (K), 61 mg kg⁻¹ of available phosphorus, 50 mg kg⁻¹ of exchangeable potassium, 655 mg kg⁻¹ of extractable calcium (Ca), and 50 mg kg⁻¹ of extractable sodium (Na). The soil was sterilized twice through autoclaving at 121 °C for 120 min at a pressure of 15 psi and kept at room temperature until it was used in the pot experiment. The pot design was conducted using a completely randomized design (CRD) with five replications as follows: T1, uninoculated with the pathogen; T2, pathogenic R. solanacearum; T3, crude extract of the endophytic fungus H. saxatilis KKU-KHP 01 + R. solanacearum; T4, endophytic fungus H. saxatilis KKU-KHP 01 + R. solanacearum; and T5, Bactericide.
The tomato seedlings were grown for 30 days and then transferred to 12-inch diameter pots filled with sterilized soil. Three days after planting, 50 mL of Hoagland’s solution was added. Fourteen days after planting, a bacterial culture of R. solanacearum at 0.5 McFarland, with a volume of 10 mL per pot, was prepared. The tomato roots were subsequently cut with a scalpel and inoculated with 10 mL of the bacterial culture. After 3 days of R. solanacearum infection, the endophytic fungus H. saxatilis KKU-KHP 01 from sorghum seeds was prepared, and the plants were treated with 10 mL of the crude extract of the endophytic fungus H. saxatilis KKU-KHP 01 at a concentration of 32 μg/mL. After seven days of R. solanacearum inoculation, 10 mL of the endophytic fungus H. saxatilis KKU-KHP 01 crude extract at a concentration of 32 μg/mL was added again. Regular watering was resumed (Kemboi et al., 2022).
Data collection
2.9
The plant data were collected 14 days after the inoculation of R. solanacearum to calculate the disease severity index (DSI) and disease reduction (DR) percentages of each treatment (Chiang et al., 2017). The severity of bacterial wilt disease was assessed using the a rating scale (0–5) from Kemboi et al. (2022), where 0 = no symptoms; 1 = 1–25 % of the leaves wilted; 2 = 26–50 % of the leaves wilted; 3 = 51–75 % of the leaves wilted; 4 = more than 75 % but less than 100 % of the leaves wilted; and 5 = all the leaves wilted and the tomato died (Kemboi et al., 2022).
Statistical analysis
2.10
Analysis of variance (ANOVA) was performed using SPSS version 26 (IBM). The data were analyzed using the least significant difference (LSD) test at a 0.05 probability level to determine significant differences between the treatments.
Results
3
Isolation and screening of the effects of fungal endophytes on bacterial growth
3.1
The 48 endophytic fungal isolates were isolated from 20 medicinal plants collected from a mountain in the Lao Hai Ngam subdistrict, Kuchinarai District, Kalasin Province, Thailand (Supplementary Table S1). All fungal endophytes were assayed for their antibacterial activity against R. solanacearum using the agar plug diffusion technique. Potential biological control activity of the fungal endophytes was observed against R. solanacearum using the agar plug diffusion test. The results indicate that among several endophytic fungi, the fungal endophyte isolates NGPM and KHP, which were isolated from Acanthus ebracteatus and Gardenia saxatilis (Geddes), respectively, could produce a clear zone around the agar plug at 18 mm and 15 mm, as shown in Fig. 1. Consequently, both endophytic fungal isolates were further identified.Fig. 1. The antagonistic activity of the endophytic fungi against R. solanacearum was determined using an agar plug diffusion test.Fig 1
Identification of fungal endophytes
3.2
The endophytic fungus KHP, cultured on oatmeal agar, exhibited slow growth at 28 °C for 14 days. The colony appeared white with fluffy mycelia. In contrast, when grown on potato dextrose agar under the same conditions, the colonies displayed yellowish‒white coloration. Microscopic examination revealed hyphal extremities and chlamydospores. The endophytic fungus was subsequently identified based on the ITS and β-Tubulin sequences. The results of the BLAST search for sequence similarity of the ITS and β-Tubulin regions are presented in Table 2. A comparison of the ITS and TUB2 sequences between the endophytic fungus KHP and the reference strain revealed that the sequence similarity value was less than 93.02 %. Based on these results, the endophytic fungal strain KHP can likely be classified as a new species. Therefore, phylogenetic placement was observed using multigene phylogenetic analyses.Table 2. The similarity of the ITS and β-Tubulin sequences between the fungal strains in this study and the top five related species of Hypoxylon in the GenBank database.Table 2. Related species of HypoxylonSequence similarity value ( %)ITSTUB2**H. begae YMJ 21593.0293.00H. aveirense CMG 2988.8087.03H. griseobrunneum CBS:331.7387.1386.40H. anthochroum YMJ 987.0086.84H. diperithecium FCATAS422686.8087.77
The phylogenetic tree generated from the maximum likelihood (ML) analysis is shown in Fig. 2. This tree placed the endophytic fungal strain KHP in a distinct clade, separated from previously known Hypoxylon species, with strong support (BS = 100 %, PP = 1.00). Based on these results, the endophytic fungal strain KHP was designated as H. saxatilis KKU-KHP01.Fig. 2. Phylogenetic tree derived from maximum likelihood analysis of 55 fungal strains harboring combined ITS and β-Tubulin genes. Xylaria hypoxylon CBS 122,620 and X. arbuscula CBS 126,415 were used as the outgroup. The numbers above the branches represent the maximum likelihood bootstrap percentages. Bootstrap values ≥70 %. The scale bar represents the expected number of nucleotide substitutions per site. The sequences of the fungal strains obtained in this study are in red.Fig 2
Biological activity of crude extracts from fungal endophytes
3.3
The ethyl acetate crude extracts of Hypoxylon sp. NGPM and Hypoxylon saxatilis KKU-KHP 01 were tested for antagonistic activities against R. solanacearum, a causative agent of bacterial wilt in tomatoes. The crude extract from both the mycelia and fermentation broth of fungal endophytes has antagonistic effects on R. solanacearum. However, fermentation broth effectively inhibited the growth of R. solanacearum, especially the fermentation broth of the endophytic fungus H. saxatilis KKU-KHP 01. Therefore, the crude extract of the endophytic fungus H. saxatilis KKU-KHP 01 was further analyzed to elucidate the structures of the bioactive compounds.
Structure elucidation of the bioactive compound
3.4
The EtOAc crude extract of H. saxatilis KKU-KHP 01 was subjected to purification using silica gel chromatography (CC) with gradients of EtOAc in n-hexane, resulting in four fractions, KHP1–KHP4. All the fractions were tested for their activity against R. solanacearum. Fraction KHP2 was purified by CC and eluted with CH_2_Cl_2_−hexanes (2:8, v/v), yielding compound 1 as a colorless oil.
Compound 1 was obtained as a colorless oil. Its molecular formula (C_9_H_14_O_2_) was deduced from the positive-ion HRESIMS m/z 155.1071 [M + H]^+^, (calcd for 155.1067, C_9_H_15_O_2_^+^). The ^1^H NMR spectrum (Table 3) revealed an olefinic proton resonance at δ_H_ 6.53 (m), an oxygenated methine resonance at δ_H_ 4.96 (m), diastereotopic methylene resonances at δ_H_ 2.84 (dd, 16.5, 7.5) and 2.33 (d, 16.5), two methylene resonances at δ_H_ 1.59 (m) and 2.06 (quintet, 7.5), and two methyl resonances at δ_H_ 0.95 (t, 7.5) and 0.86 (t, 7.5) ppm. The ^13^C NMR spectra (Table 3) revealed ten carbon resonances, including one ester carbonyl resonance at δ_C_ 170.9; two olefinic resonances at δ_C_ 141.5 and 125.8; one oxymethine resonance at δ_C_ 78.6; three methylene resonances at δ_C_ 30.4, 29.2, and 23.2; and two methyl resonances at δ_C_ 12.4 and 8.8 ppm. The planar structure of compound 1 was determined through analysis of its COSY, HSQC, and HMBC spectra. The key correlations are shown in Fig. 3. The COSY spectrum revealed two spin systems: H-4/H-5/H-6/H-7 and H-8/H-9/H-10.Table 3^1^H (400 MHz) and ^13^C NMR (100 MHz) Spectroscopic Data of **1.**Table 3. No.1 [mult, J in Hz] in CDCl_3_δHδ_C_1––2–170.93–125.842.84 (dd, 16.5, 7.5) 2.33 (d, 16.5)30.454.93 (m)78.661.59 (m)29.270.86 (t, 7.5)8.886.53 (m)141.592.06 (quintet, 7.5)23.2100.95 (t, 7.5)12.4Fig. 3Key COSY and HMBC correlations of compound **1.**Fig 3
The HMBC spectrum of H-5 to C-2 and C-3, H-8 to C-2 and C-4, and H-10 to C-8 demonstrated that the chain was located at C-3. Moreover, HMBC correlations were observed between H-7 and C-5, H-5 and C-2, and C-3, indicating that this chain was connected to C-5. Therefore, the core structure of compound 1 was identified as tetrahydrofuran. Moreover, compound 1 was therefore identified as an undescribed secondary metabolite from natural sources.
SEM studies of the effects of the compounds on bacterial cells
3.5
SEM revealed the effects of compound 1 from H. saxatilis KKU-KHP 01 on the cell structure of R. solanacearum. Compound 1 caused changes to the bacterial cell surface, with the areas exposed to the compound appearing wrinkled. Therefore, the inhibition of the growth of the pathogenic bacterium R. solanacearum may be due to the ability of pure compound 1 to damage the bacterial cell wall, as shown in Fig. 4.Fig. 4. Effects of compound 1 on the structural changes of R. solanacearum cells using SEM: (A) R. solanacearum cells without exposure to compound 1, (B) R. solanacearum cells exposed to compound 1, (arrowhead) bacterial cells showing wrinkles after exposure to compound 1.Fig 4
Disease suppression assay
3.6
Fourteen days after R. solanacearum infection, the percentage of disease severity index and disease reduction were calculated. The results reveal that when R. solanacearum was inoculated and treated with the crude extract of the endophytic fungus H. saxatilis KKU-KHP 01 at a concentration of 32 μg/mL, the disease severity index (DSI) was 16 %. This value was significantly lower than that of the co-inoculation of the endophytic fungi H. saxatilis KKU-KHP 01 and R. solanacearum, which had a DSI of 36 % (p < 0.05). Additionally, the crude extract of the endophytic fungus H. saxatilis KKU-KHP 01 was able to reduce the severity of wilt disease caused by R. solanacearum by 83.33 %, which was significantly higher (p < 0.05) than that resulting from the co-inoculation of the endophytic fungus and R. solanacearum and bactericide, resulting in disease reduction percentages of 62.50 % and 70.00 %, respectively, as shown in Table 4 and Fig. 5.Table 4. Effect of the crude extract and the endophytic fungus H. saxatilis KKU-KHP 01 on the inhibition of wilt disease in tomatoes caused by R. solanacearum under greenhouse conditions 14 days after inoculation with R. solanacearum.Table 4. TreatmentDisease severity index ( %)Disease reduction ( %)Uninoculation with the pathogen0.00± 0.000.00± 0.00R. solanacearum96.00 ± 0.08^a^0.00 ± 0.00^c^Crude extract of H. saxatilis KKU-KHP 01 + R. solanacearum16.00 ± 0.08^c^83.33 ± 0.09^a^Endophytic fungus H. saxatilis KKU-KHP 01+R. solanacearum36.00± 0.16^b^62.50 ± 0.17^b^Bactericide30.00±0.12^b^70.00±0.13^b^The disease severity index (DSI) and disease reduction (DR) were calculated 14 days after the inoculation of R. solanacearum, and the values are the means (±SD) of the percentage of R. solanacearum inhibited. The results represent the average number of tomatoes planted in the pots, with five replications per treatment. Different letters indicate statistically significant differences between the data according to the LSD test (p < 0.05).Fig. 5. Effects of the crude extract and the endophytic fungus H. saxatilis KKU-KHP 01 on the inhibition of wilt disease in tomatoes caused by R. solanacearum under greenhouse conditions 14 days after inoculation with R. solanacearum. (A) Negative control, uninoculated with the pathogen; (B) positive control, inoculated with the pathogen R. solanacearum; (C) plants inoculated with R. solanacearum and treated with the crude extract of the endophytic fungus H. saxatilis KKU-KHP 01; (D) plants inoculated with R. solanacearum and co-inoculated with the endophytic fungus H. saxatilis KKU-KHP 01; and (E) bactericide.Fig 5
Discussion
4
The ability of endophytic fungi to inhibit bacterial wilt disease caused by R. solanacearum in tomatoes was tested. Twenty species of medicinal plants were collected from mountains in the Lao Hai Ngam subdistrict, Kuchinarai District, Kalasin Province, Thailand. A total of 48 endophytic fungal isolates were obtained. This finding is consistent with recent research highlighting that medicinal plants serve as reservoirs for diverse endophytic fungi with potential bioactive properties (Gupta et al., 2023). The inhibitory effect of these fungal isolates against R. solanacearum supports their promising role as biological control agents in managing bacterial wilt disease in tomatoes. Recent studies have emphasized the ability of endophytic fungi to produce antimicrobial compounds and induce systemic resistance in host plants, thereby suppressing pathogenic bacteria (Lee et al., 2022; Zhang et al., 2024). Endophytic fungi are microorganisms that live within plant tissues without causing disease or harm to the host plant (Smith and Doe, 2023). These fungi can protect plants from pathogen invasion, such as Sclerotium rolfsii, which causes root rot disease (Suebrasri et al., 2020). Endophytic fungi have also been used to inhibit Fusarium wilt disease in tomatoes caused by Fusarium oxysporum f. sp. lycopersici (Muhorakeye et al., 2024) and to suppress bacterial spot disease in tomatoes caused by Xanthomonas vesicatoria (Rashid, 2021). However, research on the role of endophytic fungi in the inhibition of R. solanacearum is relatively limited. Therefore, the antagonistic activity of 48 endophytic fungal isolates against R. solanacearum was evaluated using the agar plug diffusion test. Two fungal isolates, NGPM and KHP, inhibited R. solanacearum. These endophytic fungi were isolated from the midribs of Acanthus ebracteatus (NGPM) and the petioles of Gardenia saxatilis (Geddes) (KHP). The isolates were subsequently classified based on their morphological and molecular characteristics. The fungi were identified as Hypoxylon sp. NGPM and Hypoxylon sp. KHP, with 100 % and 95.52 % similarity, respectively, based on ITS1–4 primers. However, based on the similarity of the TUB2 sequence to that of the reference strains, including H. begae, H. aveirense, H. griseobrunneum, H. anthochroum, and H. diperithecium, the similarity did not meet the TUB2 threshold of ≥ 87.77 % (Table 2). Therefore, H. saxatilis can be considered a new and different species from these five species. Moreover, the secondary metabolites from these two endophytic fungi were extracted using both fermentation broth and mycelia. Notably, the fermentation broth had a greater inhibitory effect on R. solanacearum than did the mycelial extracts. Between the two fungi, H. saxatilis KKU-KHP 01 exhibited more efficient inhibition. This difference can be attributed to the fact that the fermentation broth contains extracellular metabolites secreted by the fungi during growth, which are often more bioavailable and potent against pathogens (Kusari et al., 2012). The secondary metabolites were purified using column chromatography and identified as compound 1, a tetrahydrofuran (THF) derivative. NMR analysis confirmed that compound 1 was the first report of a new naturally occurring secondary metabolite. In addition, the minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) of compound 1 were found to be 0.5 µg/mL and 1.0 µg/mL, respectively, which were lower than those of the two antibiotics used as positive controls, chloramphenicol and kanamycin. SEM analysis revealed that R. solanacearum exposed to tetrahydrofuran (THF) exhibited cell wall wrinkles, a well-known characteristic of THF toxicity against bacteria (Yao et al., 2010). The general mechanism of action of THF includes damage to the bacterial cell membrane (Zhang et al., 2014), protein denaturation (Hochkoeppler et al., 2007), inhibition of bacterial metabolism by targeting enzymes involved in energy production and growth (Tian et al., 2019), and DNA damage (Hedrick et al., 2014). Therefore, tetrahydrofuran (THF), which is secreted by the endophytic fungus H. saxatilis KKU-KHP 01, can potentially destroy the bacterial pathogen R. solanacearum. Moreover, this study is the first report on the use of the novel secondary metabolite tetrahydrofuran produced by the endophytic fungus H. saxatilis KKU-KHP 01 for inhibiting the growth of R. solanacearum. Previously reported, crude extracts from the endophytic fungus Trichoderma spp. reduced bacterial wilt severity in tomatoes caused by R. solanacearum by 60–75 % (Siddiqui et al., 2017), and crude extracts from the endophytic fungus Penicillium spp. reduced bacterial wilt severity by 40–60 % (Sarma et al., 2015). In the present study, the crude extract of H. saxatilis KKU-KHP 01 was evaluated under greenhouse conditions for its ability to reduce the severity of bacterial wilt disease caused by R. solanacearum. The results showed an 83.33 % reduction in disease severity, which was significantly higher than the treatment with bactericide and those in previous reports. Therefore, tetrahydrofuran isolated from the endophytic fungus H. saxatilis KKU-KHP 01 exhibits promising potential for controlling bacterial wilt disease in tomatoes under field conditions. Furthermore, this compound could be further developed into a commercial biocontrol agent for sustainable agricultural applications.
Conclusion
5
This study demonstrated that endophytic fungi isolated from medicinal plants, particularly Hypoxylon saxatilis KKU-KHP 01, possess strong antibacterial activity against R. solanacearum, the causative agent of wilt disease in tomatoes. Notably, H. saxatilis KKU-KHP 01 was identified as a novel fungal species capable of producing tetrahydrofuran, a compound not previously reported as a natural fungal metabolite. Tetrahydrofuran significantly inhibited the pathogen by disrupting bacterial cell wall integrity, as observed through scanning electron microscopy. Under greenhouse conditions, the crude extract of H. saxatilis KKU-KHP 01 reduced wilt severity by up to 83.33 %. These findings highlight the potential application of tetrahydrofuran as a biocontrol agent for managing bacterial wilt in tomato cultivation.
Credit author statement
NameCreditThanapat SuebrasriConceptualization, Methodology, Investigation, Data analysis and Writing - Original DraftWasan SeemakramConceptualization, Methodology, Investigation, data analysis and funding acquisitionAwat WisetsaiInvestigation some experiment and data analysisThanawan GatetaInvestigation some experiment and data analysisSutarin PreepramInvestigation some experiment and data analysisPhornnapa SaentaoInvestigation some experiment and data analysisSophon BoonlueConceptualization, Writing- Reviewing and Editing, funding acquisition
Funding
This research was funded by a grant from the Fundamental Fund of 10.13039/501100004071Khon Kaen University through the National Science, Research and Innovation Fund (NSRF). We are grateful to the Plant Genetic Conversion Project under the Royal Initiative of Her Royal Highness Princess Maha Chakri Sirindhorn, Khon Kaen University.
Data availability statement
The datasets obtained and analyzed in the current study are available from the corresponding author upon reasonable request.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Agarwal A.Singh P.Yadav A.Evaluation of endophytic microbial consortium for the management of bacterial wilt of tomato caused by Ralstonia solanacearum Biocontrol. Sci. Technol.314202145346510.1080/09583157.2021.1893761 · doi ↗
- 2Becker K.Lambert C.Wieschhaus J.Stadler M.Phylogenetic assignment of the fungicolous hypoxylon invadens (Ascomycota, xylariales) and investigation of its secondary metabolites Microorganisms 89202011410.3390/microorganisms 8091397 PMC 756571632932875 · doi ↗ · pubmed ↗
- 3Bills G.F.González-Menéndez V.Martín J.Platas G.Fournier J.Peršoh D.Stadler M.Hypoxylon pulicicidum sp. nov. (Ascomycota, Xylariales), a pantropical insecticide-producing endophyte P Lo S. One 710201210.1371/journal.pone.0046687 PMC 346729023056404 · doi ↗ · pubmed ↗
- 4Boucher C.A.Copper-based bactericides for controlling bacterial wilt caused by Ralstonia solanacearum Plant Dis. J.891220051373137810.1094/PD-89-1373 · doi ↗
- 5Cedeño-Sanchez M.Wendt L.Stadler M.Mejía L.C.Three new species of Hypoxylon and new records of Xylariales from Panama Mycosphere 11120201457147610.5943/mycosphere/11/1/9 · doi ↗
- 6Chauhan P.Verma P.Mishra A.Bio Avengers: how do endophytic microorganisms alter a plant's defense mechanisms?J. Plant Sci. Phytopathol.8202400100610.29328/journal.jpsp.1001123 · doi ↗
- 7Chiang K.S.Liu H.I.Bock C.H.A discussion on disease severity index values. Part I: warning on inherent errors and suggestions to maximise accuracy Ann. Appl. Biol.1712201713915410.1111/aab.12362 · doi ↗
- 8Dai D.Q.Phookamsak R.Wijayawardene N.N.Li W.J.Bhat D.J.Xu J.C.Taylor J.E.Hyde K.D.Chukeatirote E.Bambusicolous fungi Fungal. Divers.8212017110510.1007/s 13225-016-0367-8 · doi ↗