Identification of Amino Acid Conservation in The Curli Accessory Protein CsgF
Karen Guerrero, Emma Smith, Shruti Sunder Rajkumar, Zachary Cairo, Jonathan Adame, Renad Rawas, Ranim Rawas, Sajith Jayasinghe

TL;DR
This study identifies conserved amino acids in CsgF, a protein involved in bacterial Curli assembly, which may help understand its structural and functional roles.
Contribution
The study reports novel conserved residues in CsgF that have not been previously linked to its function in Curli formation.
Findings
Conserved Pro and Gly residues in the N-terminal region may be important for loop conformation.
Conserved hydrophobic residues in the C-terminal β-sheet and C-terminus may be essential for Curli formation.
Several conserved residues identified in this study have not been previously reported as functionally important.
Abstract
The Curli-Specific gene product F (CsgF) plays an important role in the assembly of gram-negative bacterial cell surface filaments known as Curli. In order to evaluate amino acid conservation in the context of the solution and CsgG bound structures of CsgF we carried out a multiple sequence alignment of CsgF sequences from 35 gram-negative bacteria and correlated amino acid conservation to structural and functional importance. We identified conserved Pro and Gly residues within the N-terminal region of CsgF that may be required to adopt the loop confirmation observed in this region. Several conserved hydrophobic residues are found on the 3rd and 4th β-strands of the C-terminal β-sheet and extending to the C-terminal end, that may play a role in the reported observation that the C-terminus is needed for Curli formation. The importance of several conserved residues that were identified in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
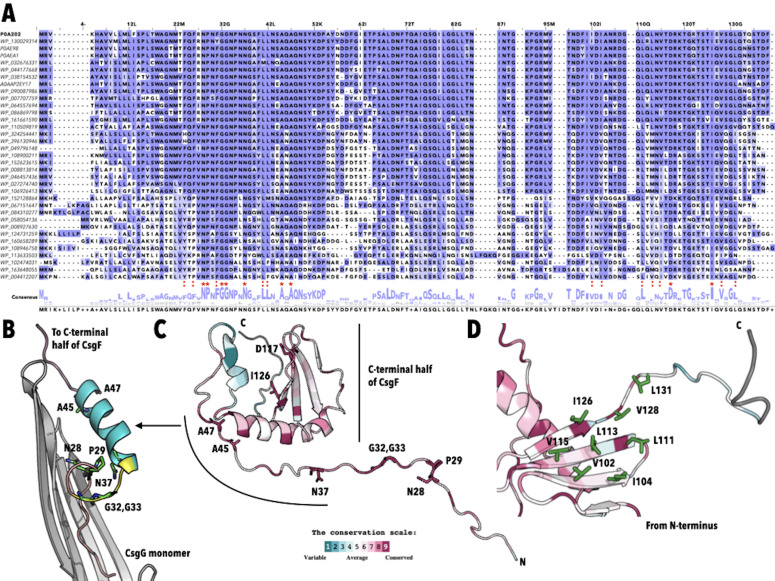 Figure 1
Figure 1- —National Institute of General Medical Sciences (United States)https://ror.org/04q48ey07
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Reproductive Biology · Biochemical and Structural Characterization · Legume Nitrogen Fixing Symbiosis
Description
The outer surface of gram-negative bacteria contain filaments, known as Curli, that play an important role in cell-cell interactions and host-cell colonization (Collinson et al., 1997, Sukupolvi et al., 1997, Austin et al., 1998, Römling et al., 1998). Curli are composed of two Curli specific gene (Csg) proteins, CsgA and CsgB, with CsgA being the major component of the Curli filaments (Chapman et al., 2002, Hammar et al., 1996, Bian & Normark, 1997). The assembly of CsgA/CsgB into Curli involves four other proteins: CsgC, CsgE, CsgF, and CsgG. CsgC and CsgE are periplasmic proteins that are thought to prevent the intracellular aggregation of CsgA/CsgB and target these proteins to the outer membrane channel CsgG (Gibson et al., 2007, Taylor et al., 2011, Evans et al., 2015). CsgA and CsgB traverse through CsgG and are secreted to the bacterial outer surface where they associate to form Curli (Cao et al., 2014, Goyal et al., 2014). In contrast, CsgF is located on the extracellular surface of bacteria and is required for Curli assembly (Robinson et al., 2006, Nenninger et al., 2009, Chapman et al., 2002). Structural studies have shown that the N-terminus of CsgF interacts with the CsgG pore, and in-vivo complementation assays have shown that the C-terminal half of the protein is required for proper Curli assembly (Zhang et al., 2020, Yan et al., 2020, Swasthi et al., 2023). A majority, if not all, of the structure/function studies reported so far have concentrated on E. Coli CsgF. We carried out a multiple sequence alignment (MSA) of CsgF sequences from 35 gram-negative bacteria (see extended data) to determine amino acid conservations and evaluated these in the context of the three-dimensional structures of CsgF to correlate amino acid conservation to structural and functional importance.
The MSA shows significant conservation throughout the sequence ( Figure 1A ). There are nine amino acids that are conserved in all 35 sequences (numbered according to the position in the Salmonella sequence): N28, P29, G32, G33, N37, A45, A47, D117, and I126 ( Figure 1A, red star "*"). We evaluated if these residues contributed to the interaction of CsgF with the CsgG channel using the cryo-EM structures of the CsgF-CsgG complexes available in the protein data bank (PDB ID 6L7C and 6LQH) (Zhang et al., 2020) (Yan et al., 2020). The N-terminal residues (20-31) of CsgF are observed to interact with the inner surface of the CsgG channel in an extended conformation ending in a loop ( Figure 1B, salmon). The loop which extends between residues 29-36 of CsgF ( Figure 1B, yellow) is followed by an 𝛼-helix that spans residues 37-46 ( Figure 1B, teal). Seven of the nine amino acids found in all 35 sequences, N28, P29, G32, G3, N37, A45, and A47, are found in the region that contains these three structural elements ( Figure 1B ). N28 (100% conservation) and N30, which is conserved in 74% of the sequences included in this study, forms H-bonds with E216 of CsgG suggesting that they may play a role in anchoring CsgF to CsgG. Yan et. al report a reduced interaction of CsgF with CsgG for the N28A variant but not for the N30A variant (Yan et al., 2020). The loop region contains P29, G32, and G33 which are found in all 35 sequences. These Pro and Gly residues, both of which are found in structural motifs requiring tight turns or flexibility, may play an important role in this region’s ability to adopt the loop observed in the CsgG bound form of CsgF. N37 which is conserved in all 35 sequences forms a H-bond with the backbone carbonyl of R27 and may also interact with the backbone carbonyl of P29 from an adjacent CsgF (which is 3.5 A away), and therefore may also play a role the formation of the loop conformation. As far as we know the impact of variants of these four residues have not been reported but would help in determining the influence of the loop region on the function of CsgG bound CsgF. A45 and A47, which are also found in all 35 sequences, are found on the C-terminal end of the 𝛼-helix that associates with the upper part of the CsgG pore. Again, no variants of these two alanine residues have been investigated and therefore, it is not known if these residues play a role in the association of CsgF with CsgG or its function.
Residues that have been identified as helping anchor CsgF to CsgG via van der Waals or hydrophobic interactions F24 (60% F, 34% Y, F26 (66% F, 34% Y, F31 (97% F, 3% Y), L41 (97% L) and L42 (91% L 6% F), are also conserved ( Figure 1A, red " :" ) (Zhang et al., 2020, Yan et al., 2020). Yan et. al report that the L41D and L42D variants of CsgF reduced its interaction with CsgG and that the F24D, F26D, and F31D, completely abolished the ability of CsgF to interact with CsgG (Yan et al., 2020) . These observations correlate with the conservation observed for these five amino acids.
The cryo-EM structure of the CsgF-CsgG complex does not contain the C-terminal residues of CsgF. In order to visualize the amino acid conservation in the context of structure we used the ConSurf server to map evolutionary conservation onto the three-dimensional solution structure of CsgF (PDBID 5M1U, Figure 1C ) (Yariv et al., 2023)(Schubeis et al., 2018) (Landau et al., 2005). We observe significant conservation throughout the solution structure of CsgF ( Figure 1C, purple) similar to what was observed via the MSA. In addition to conservation in the N-terminal half of the protein there is considerable conservation of residues in the 𝛼-helix and 𝛽-sheet found in the C-terminal half of the solution structure of CsgF. It has been observed that the 𝛽-sheet contains a hydrophobic surface formed from positions 102, 104, 113, and 115 on the 3rd and 4th 𝛽-strands, and it was speculated that this hydrophobic surface may facilitate protein interactions (Schubeis et al., 2018). The MSA and the evolutionary conservation predicted by ConSurf indicate that the hydrophobic surface is conserved and extends towards the unstructured C-terminal end to include position 126, 128 and 131 ( Figure 1D ). I126 is found in all 35 sequences investigated here and positions 102 (69% V, 23% I, 9% L), 104 (66% I, 31% V, 3% L), 111 97% L, 3% F), 113 (40% L, 21% V, 9% I), 115 (54% V, 46% I), 128 (94% V, 3% I, 3% L), 131 (77% L, 11% I, 3% V) are occupied by amino acids with hydrophobic side chains ( Figure 1A red :). The conservation of these hydrophobic residues correlates well with the observation that this region is important for Curli assembly. In vivo complementation assays have shown that the C-terminal half of CsgF is needed for Curli assembly and that this region binds the nucleator protein CsgB (Swasthi et al., 2023, Yan et al., 2020, Schubeis et al., 2018). It appears that the 11 residues (between 128-138) at the C-terminal end of CsgF is required for interaction with CsgB and the proper assembly of Curli (Swasthi et al., 2023). D117 is conserved in all 35 sequences of CsgF used in this study however its importance to the structure or function of CsgF is not known.
Conclusion: Using a multiple sequence alignment of CsgF sequences from 35 bacterial species, as well as an evolutionary conservation analysis using the ConSurf server, we were able to identify significant amino acid conservation throughout the protein. We were able to correlate several of the conserved positions to published reports describing the regions important for the structure and function of CsgF. However there remains positions of amino acid conservation whose importance is not yet known. A systematic investigation of the conserved positions identified here may help add to our understanding of Curli assembly.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Austin John W Sanders Greg Kay William W Collinson S.Karen 199851 Thin aggregative fimbriae enhance Salmonella enteritidis biofilm formation FEMS Microbiology Letters 16220378-109729530110.1111/j.1574-6968.1998.tb 13012.x 9627964 · doi ↗ · pubmed ↗
- 2Bian Zhao Normark Staffan 1997101 Nucleator function of Csg B for the assembly of adhesive surface organelles in Escherichia coli The EMBO Journal 16190261-41895827583610.1093/emboj/16.19.58279312041 PMC 1170214 · doi ↗ · pubmed ↗
- 3Cao Baohua Zhao Yan Kou Yongjun Ni Dongchun Zhang Xuejun Cai Huang Yihua 2014121 Structure of the nonameric bacterial amyloid secretion channel Proceedings of the National Academy of Sciences 111500027-842410.1073/pnas.1411942111 PMC 427332625453093 · doi ↗ · pubmed ↗
- 4Chapman Matthew R. Robinson Lloyd S. Pinkner Jerome S. Roth Robyn Heuser John Hammar Mårten Normark Staffan Hultgren Scott J. 200221 Role of Escherichia coli Curli Operons in Directing Amyloid Fiber Formation Science 29555560036-807585185510.1126/science.106748411823641 PMC 2838482 · doi ↗ · pubmed ↗
- 5Collinson S. Karen Clouthier Sharon C. Doran James L. Banser Pamela A. Kay William W. 1997 Characterization of the Agf BA Fimbrial Operon Encoding Thin Aggregative Fimbriae of Salmonella Enteritidis Advances in Experimental Medicine and Biology 0065-259824724810.1007/978-1-4899-1828-4_379192021 · doi ↗ · pubmed ↗
- 6Evans Margery L. Chorell Erik Taylor Jonathan D. Åden Jörgen Götheson Anna Li Fei Koch Marion Sefer Lea Matthews Steve J. Wittung-Stafshede Pernilla Almqvist Fredrik Chapman Matthew R. 201521 The Bacterial Curli System Possesses a Potent and Selective Inhibitor of Amyloid Formation Molecular Cell 5731097-276544545510.1016/j.molcel.2014.12.02525620560 PMC 4320674 · doi ↗ · pubmed ↗
- 7Gibson D. L White A. P Rajotte C. M Kay W. W 200741 Agf C and Agf E facilitate extracellular thin aggregative fimbriae synthesis in Salmonella Enteritidis Microbiology 15341350-08721131114010.1099/mic.0.2006/000935-017379722 · doi ↗ · pubmed ↗
- 8Goyal Parveen Krasteva Petya V. Van Gerven Nani Gubellini Francesca Van den Broeck Imke Troupiotis-Tsaïlaki Anastassia Jonckheere Wim Péhau-Arnaudet Gérard Pinkner Jerome S. Chapman Matthew R. Hultgren Scott J. Howorka Stefan Fronzes Rémi Remaut Han 2014914 Structural and mechanistic insights into the bacterial amyloid secretion channel Csg G Nature 51675300028-083625025310.1038/nature 1376825219853 PMC 4268158 · doi ↗ · pubmed ↗