The tilt ( tt ) mutation of Drosophila melanogaster maps to the cis-regulatory region of the Iroquois Complex (Iro-C) located in the sosondowah ( sowah) gene
Samuel Gruber, Arno Houtman, Hailey Reisert, Mina Amini, Caroline Fiore, Paula Gonzalez, Veronica Han, Aeva Jazic, Mie Kusupholnand, Max Miller, Jiung Nam, Ziqin Wang, Yang Yu, Peter Dong, Allen S. W. Oak, Arun Sharma, Eric P Spana

TL;DR
This paper identifies the genetic location of the tilt mutation in fruit flies and links it to regulatory elements of the Iroquois Complex genes.
Contribution
The study maps the tilt mutation to the cis-regulatory region of the Iroquois Complex within the sosondowah gene.
Findings
The tilt mutation is located in the sosondowah gene, which regulates the Iroquois Complex genes araucan and caupolican.
A gtwin retrotransposon with su(Hw) insulator binding sites is associated with the tilt mutation.
Mutations in su(Hw) suppress the tilt phenotype, suggesting a functional link.
Abstract
The tilt ( tt ) mutation first described by Morgan and Bridges in 1915 has adult visible phenotypes in wing posture and vein formation. We have mapped tt to a genomic region within the sosondowah ( sowah ) gene that houses the cis-regulatory elements that control expression of the Iroquois Complex genes araucan ( ara ) and caup olican ( caup ) in the wing hinge and wing veins. Sequence analysis of the tt 1 allele has identified a gtwin retrotransposon containing su(Hw) insulator binding sites within this region of sowah . We find that mutations in su(Hw) suppress the tilt phenotype, providing a potential mechanism for the insertion to affect the function of the Iroquois Complex.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
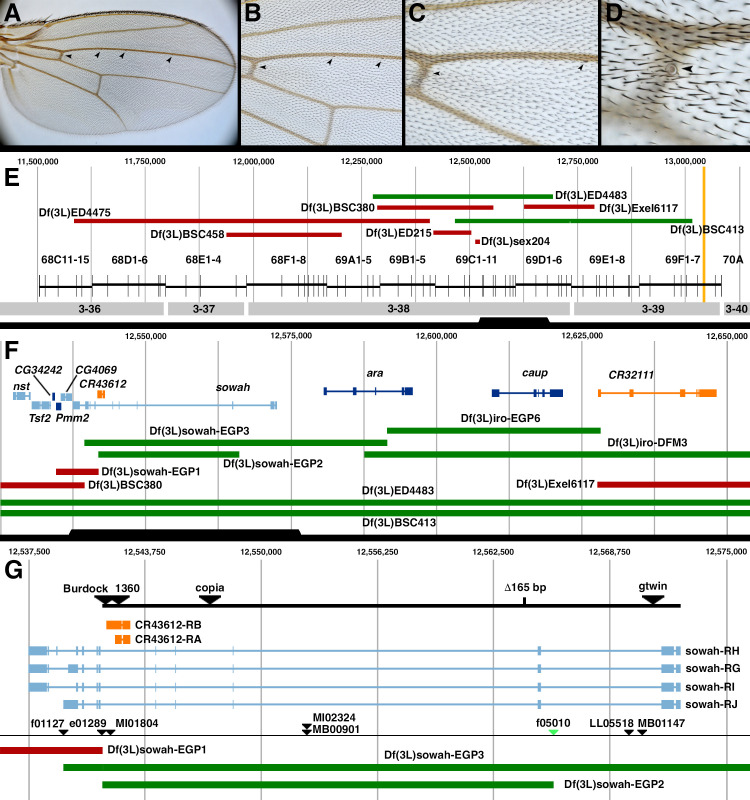 Figure 1
Figure 1|
Reagent Type |
Designation |
Identifier |
Reference/ Source |
Additional Information |
|
Genetic Reagent (
|
|
BDSC 623 |
Bloomington Drosophila Stock Center |
Genotype:
|
|
Genetic Reagent (
|
|
BDSC 1053 |
Bloomington Drosophila Stock Center |
Genotype:
|
|
Genetic Reagent (
|
OreR |
N/A |
Lab Strain |
Wild type |
|
Genetic Reagent (
|
Df(3L)BSC380 |
BDSC 24404 |
Bloomington Drosophila Stock Center |
Genotype:
|
|
Genetic Reagent (
|
Df(3L)BSC413 |
BDSC 24917 |
Bloomington Drosophila Stock Center |
Genotype:
|
|
Genetic Reagent (
|
Df(3L)BSC458 |
BDSC 24962 |
Bloomington Drosophila Stock Center |
Genotype:
|
|
Genetic Reagent (
|
Df(3L)iro-DFM3 |
BDSC 36531 |
Bloomington Drosophila Stock Center |
Genotype:
Df(3L)iro-DFM3,
|
|
Genetic Reagent (
|
Df(3L)iro-EGP6 |
N/A |
Sonsoles Campuzano (Universidad Autónoma de Madrid) (Carrasco-Rando et al., 2011) |
Genotype: w*; Df(3L)iro-EGP6/TM3, Sb, Ser |
|
Genetic Reagent (
|
Df(3L)ED215 |
BDSC 8071 |
Bloomington Drosophila Stock Center |
Genotype:
|
|
Genetic Reagent (
|
Df(3L)ED4475 |
BDSC 8069 |
Bloomington Drosophila Stock Center |
Genotype:
|
|
Genetic Reagent (
|
Df(3L)ED4483 |
BDSC 8070 |
Bloomington Drosophila Stock Center |
Genotype:
|
|
Genetic Reagent (
|
Df(3L)Exel6117 |
BDSC 7596 |
Bloomington Drosophila Stock Center |
Genotype:
|
|
Genetic Reagent (
|
Df(3L)Sex204 |
BDSC 9337 |
Bloomington Drosophila Stock Center |
Genotype:
|
|
Genetic Reagent (
|
Df(3L)sowah-EGP1 |
N/A |
Sonsoles Campuzano (Universidad Autónoma de Madrid) (Maeso et al., 2012) |
Genotype: w*; Df(3L)sowah-EGP1/TM3, Sb |
|
Genetic Reagent (
|
Df(3L)sowah-EGP2 |
N/A |
Sonsoles Campuzano (Universidad Autónoma de Madrid) (Maeso et al., 2012) |
Genotype: w*; Df(3L)sowah-EGP2/TM3, Sb |
|
Genetic Reagent (
|
Df(3L)sowah-EGP3 |
N/A |
Sonsoles Campuzano (Universidad Autónoma de Madrid) (Maeso et al., 2012) |
Genotype: w*; Df(3L)sowah-EGP3/TM3, Sb |
|
Genetic Reagent (
|
sowah-f01127 |
BDSC 18412 |
Bloomington Drosophila Stock Center |
Genotype:
|
|
Genetic Reagent (
|
sowah-e01289 |
BDSC 85121 |
Bloomington Drosophila Stock Center |
Genotype:
|
|
Genetic Reagent (
|
sowah-MI01804 |
BDSC 37311 |
Bloomington Drosophila Stock Center |
Genotype:
|
|
Genetic Reagent (
|
sowah-MI02324 |
BDSC 35109 |
Bloomington Drosophila Stock Center |
Genotype:
|
|
Genetic Reagent (
|
sowah-MB00901 |
BDSC 22927 |
Bloomington Drosophila Stock Center |
Genotype:
|
|
Genetic Reagent (
|
sowah-f05010 |
N/A |
Sonsoles Campuzano (Universidad Autónoma de Madrid) (Maeso et al., 2012) |
Genotype: w*; sowah-f05010/TM6B |
|
Genetic Reagent (
|
sowah-LL5518 |
Kyoto 141612 |
Kyoto Drosophila Stock Center |
Genotype:
|
|
Genetic Reagent (
|
sowah-MB01147 |
BDSC 23001 |
Bloomington Drosophila Stock Center |
Genotype:
|
|
Software |
Helicon Focus |
N/A |
| |
|
Software |
minimap2 |
N/A |
| |
|
Software |
Tablet |
N/A |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDevelopmental Biology and Gene Regulation · Genomics and Chromatin Dynamics · Neurobiology and Insect Physiology Research
Description
Mutations that changed the size, location, or presence of wing veins have been identified since the dawn of Drosophila genetics (Bridges & Morgan, 1919, 1923; Morgan & Bridges, 1916). These mutations have been instrumental in establishing the formation and differentiation of wing veins as a model for developmental patterning. Vein 3 is essentially established by the hedgehog ( * hh * ) and decapentaplegic ( * dpp * ) signal transduction pathways which activate the expression of a set of transcription factors of the Iroquois Complex (Iro-C) (Gomez-Skarmeta et al., 1996). These pro-veins are then refined by the * dpp * , EGF Receptor , and Notch signaling pathways (Tripathi & Irvine, 2022). A loss of vein phenotype can happen in multiple steps of the vein specification pathway from wing disc patterning, pre-patterning veins, establishing pro-veins, and finally veins.
The tilt mutation was identified by Calvin Bridges in August 1915 and mapped to 0.2 map units to the left of Dichaete (Bridges & Morgan, 1923). The tilt phenotype was described as having wings that were held out and up from the body and a loss of a portion of vein 3. In addition to the loss of vein, tilt mutant wings never had the full complement of the campaniform sensilla normally found on vein 3 (Thompson Jr. et al., 1982). A recent paper (Houtman et al., 2023) describes in detail the three observable phenotypes found in adult * tilt ^1^ * flies: vein loss, sensilla loss, and wing posture defects. Of these three, only the sensilla loss phenotype is penetrant enough to accurately score tilt via a complementation test. One early experiment in mapping tilt placed it to the left (distal) of Minute-h (Mossige, 1938) which is now called M(3)69E , and more recent analysis has identified M(3)69E as * RpS4 * (Marygold et al., 2007). Because we now had a phenotype that could be accurately scored, and a more defined genetic location, we proceeded to map the tilt mutation in an undergraduate lab course. We elected to focus our mapping to the left of * RpS4 * (yellow line in Figure 1E ) and selected deficiency strains between 68C and 69F to test complementation by sensilla loss.
We selected eight deficiency strains and tested complementation with * tilt ^1^ * in mounted wings. Because the position of the sensilla can vary in tilt mutants (Houtman et al., 2023), we counted the anterior crossvein sensilla (ACV) and the total number of the three sensilla on vein 3 (L3-1, L3-2, L3-3) for each wing. Figure 1A- D shows the location of these four sensilla in a wild-type OreR wing at increasing magnifications. We found six deficiency strains complemented * tilt ^1^ * (visualized in Figure 1E in red): Df(3L)ED4475 (n=10, ACV=10/10, L3=32/30); Df(3L)BSC458 (n=20, ACV=20/20, L3=60/60); Df(3L)BSC380 (n=19, ACV=19/19, L3=58/57); Df(3L)ED215 (n=10, ACV=10/10, L3=32/30); Df(3L)Sex204 (n=14, ACV=14/14, L3=45/42); and Df(3L)Exel6117 (n=17, ACV=17, L3=52/51) where n is the number of wings counted, and ACV and L3 are the number of sensilla observed over the expected. Wild-type wings (OregonR) always has an ACV, and will occasionally have a fourth sensilla on vein 3, but never has fewer than three–indicating that wild-type typically has more than expected L3 sensilla (n=56, ACV=56, L3=169/168). We also found two deficiency strains that failed to complement: Df(3)ED4483 (n=21, ACV=7/21, L3=24/63); and Df(3L)BSC413 (n=12, ACV=0/12, L3=6/36) visualized in Figure 1E in green. The results for the non-complementing deficiencies were similar to * tilt ^1^ * homozygotes (n=61, ACV=4/61, L3=45/183). These crosses place the tilt locus between the right breakpoint of Df(3L)BSC380 and the left breakpoint of Df(3L)Exel6117.
Figure 1F shows a higher resolution map of the region that, when deleted, fails to complement tilt . Within this region are three protein coding genes: sosondowah ( * sowah * ), araucan ( * ara * ), and * caup olican * ( * caup * ). The transcription factors * ara * and * caup * are redundant homeodomain proteins of the Iroquois Complex which help specify cell fates in many tissues in Drosophila development, including veins 3 and 5 in the wing (Gomez-Skarmeta et al., 1996). * sowah * is a predicted cell adhesion protein whose large introns house the regulatory regions that drive * ara * and * caup * expression in vein 3 (Maeso et al., 2012). We obtained smaller deficiencies throughout the * sowah * , * ara * , and * caup * genes, and tested their complementation with tilt by examining mounted wings and scoring in the same way ( Figure 1F ). We found that Df(3L)sowah-EGP1 complemented tilt (n=19, ACV=19/19, L3=58/57). Df(3L)sowah-EGP1 deletes the 3' end of sowah, and completely removes CG4096, Pmm2, and CG34242. This indicates that tilt is not a loss of function allele of * sowah * . We found that deficiencies that removed the transcripts of both * ara * and * caup * failed to complement tilt : first, Df(3L)iro-EGP6 (n=18, ACV=7/18, L3=3/54) deletes the 3' end of * ara * and all of * caup * and second, Df(3L)iro-DMF3 (n=23, ACV=7/23, L3=23/69) that deletes about half of * ara * and all of * caup * and extends off the map for ~44 kb. Two deficiencies that remove regions of * ara/ caup * regulatory regions also fail to complement tilt : first, Df(3L)sowah-EGP3 (n=17, ACV=3/17, L3=2/51) removes most of * sowah * , CR43612, and the 5' end of * ara * ; and second, Df(3L)sowah-EGP2 (n=32, ACV=3/32, L3=2/96) removes CR43612 and a large portion of sowah, but does not remove either * ara * or * caup * transcription units. Because we could rule out sowah by complementation, and because * ara * and * caup * are redundant, we hypothesized the lesion in * tilt ^1^ * would be in the regulatory regions found within the * sowah * introns, which is roughly the breakpoints of Df(3L)sowah-EGP2. In addition to the loss of campaniform sensilla, the Df/ * tt ^1^ * lines that fail to complement tilt frequently have the wing posture defect found in tilt homozygotes and the very rare loss of wing vein 3 as seen in Houtman et al., 2023.
Having narrowed the tilt locus to the regulatory region of Iro-C within of * sowah * , we performed complementation tests with eight publicly available transposon insertions. Figure 1G shows the * sowah * region that fails to complement tilt with the appropriate deficiency strains at the bottom. Just above the deficiencies are the positions of the transposon insertions tested. Of these eight insertions, seven fully complemented tilt : * sowah ^f01127^ * (n=16, ACV=16/16, L3=52/48); * sowah ^e01289^ * (n=10, ACV=10/10; L3=31/30); * sowah ^MI01804^ * (n=10, ACV=10/10; L3=32/30); * sowah ^MI02324^ * (n=10; ACV=10/10; L3=34/30); * sowah ^MB00901^ * (n=9; ACV=9/9; L3=31/27); * sowah ^LL05518^ * (n=10; ACV=10/10; L3=31/30); and * sowah ^MB01147^ * (n=10; ACV=10/10; L3=31/30). One insertion, * sowah ^f05010^ * (shown in green) failed to complement tilt (n=27, ACV=4/27, L3=2/81). In addition to failing to complement tilt by the campaniform sensilla assay, * tilt ^1^ * / * sowah ^f05010^ * shows a strong held-out wing phenotype exactly as shown in * tilt ^1^ * homozygotes (Houtman et al., 2023).
With an identified molecular region of the * tilt ^1^ * mutation due to its failure to complement * sowah ^f05010^ * and Df(3L)sowah-EGP2, we attempted to identify the lesion in the tilt genome by using whole genome sequencing by Oxford Nanopore long-read technology. After de-novo assembly, and by aligning the * tilt ^1^ * sequence reads to the Drosophila reference genome, we have identified a number of changes found in the * tilt ^1^ * sequence that are not found in the reference genome. The thick black bar in Figure 1G shows the region between the left breakpoint of EGP2 and the transcription start site of * sowah * that was examined in detail. We found three retrotransposons within the Df(3L)sowah-EGP2 region: a Burdock element of 6,376 bp, a 1360 element of 1,474 bp, and a copia element of 5,113 bp. In addition, we found a fourth retrotransposon insertion just outside the region, a gtwin element of 6,870 bp. Finally, we found a deletion of 165 bp, 39 smaller deletions between 1 and 30 bp, 45 insertions between 1 and 22 bp, and 447 single nucleotide polymorphisms in the region shown in the bar. We did not find any changes in the * tilt ^1^ * genome that might be an obvious choice to be the causative mutation.
Because the * tilt ^1^ * allele was a spontaneous mutation it seems likely that it would be a single genomic lesion that would affect both hinge and vein expression of the Iroquois Complex. Any SNP or indel would be unlikely to alter expression from both the hinge and vein 3 enhancers as they are under different upstream regulatory signaling pathways (Ayala-Camargo et al., 2013; Hatini et al., 2013; Johnstone et al., 2013) (Crozatier et al., 2004). We hypothesized an alternative mechanism that might affect a larger regulatory region. The * sowah ^f05010^ * transposon insertion is a PBac{WH} vector that contains a suppressor of Hairy wing ( su(Hw) ) binding site, sometimes called a gypsy insulator element (Thibault et al., 2004). This sequence binds the su(Hw) transcriptional insulator (Parkhurst et al., 1988), and might cause the vein 3 enhancers and wing hinge enhancers of * ara * and * caup * found in the * sowah * intron to be separated from the * ara * and * caup * promoters by putting them into different Topological Associated Domains (TADs) (Maeso et al., 2012; Peterson et al., 2021). Wing discs of * sowah ^f05010^ * homozygous larvae show a loss of both * ara * and * caup * expression in vein 3 and hinge region (Maeso et al., 2012). The gtwin retrotransposon found in * tilt ^1^ * is related to the gypsy retrotransposon (Kotnova et al., 2006; Ludwig & Loreto, 2007), and also contains su(Hw) binding sites and binds su(Hw) (Gerland et al., 2017). Thus, although the gtwin does not lie within the Df(3L)sowah-EGP2 region, it may keep the vein 3 and wing hinge enhancers from interacting with the * ara * and * caup * promoters. The Burdock, copia, and 1360 elements do not contain su(Hw) binding sites.
To test whether the * tilt ^1^ * campaniform sensilla phenotype was related to the su(Hw) binding sites found in the gtwin retrotransposon insertion, we made flies homozygous for both * tilt ^1^ * and * su(Hw) ^8^ * . The * su(Hw) ^8^ * allele is adult viable and has been shown to suppress gypsy insertion phenotypes in alleles of yellow ( * y * ), cut ( * ct * ), and forked ( * f * ) (Georgiev & Kozycina, 1996) (Guida et al., 2016; Li, Heng, 2018). In * tilt ^1^ * , * su(Hw) ^8^ * mounted wings, we found complete suppression of the tilt phenotype (n=45, ACV=45/45, L3=150/135) compared to * tilt ^1^ * alone (n=61, ACV=4/61, L3=45/183) or w; * su(Hw) ^8^ * alone (n=40, ACV=40/40, L3=123/120). In addition, we did not observe any * tilt ^1^ * , * su(Hw) ^8^ * flies with the wing posture defect that is found in 50% of * tilt ^1^ * homozygotes (Houtman et al., 2023). This indicates that the tilt phenotype in both the hinge and vein is dependent on su(Hw) .
In summary, we have mapped the * tilt ^1^ * allele to the wing enhancers for the Iroquois Complex that are found within the * sowah * gene. Though we cannot rule out that any number of mutations within the region might contribute to the phenotype, we found that the tilt phenotype is dependent on a functional su(Hw) gene, implying that the gtwin retrotransposon is the causative lesion in the tilt genome.
Methods
** D. melanogaster stocks and genetics: **
Fly stocks and crosses were maintained on cornmeal molasses agar food (Archon Scientific). Crosses were performed at 25° C, and males were used for all analyses. Adult wing imaging was done exactly as described in Houtman et al., 2023. Images shown are Z-stacks with image depths of between 21 mm ( Figure 1A ) and 11 mm ( Figure 1D ) created by Helicon Focus software.
Whole Genome Sequencing:
Genomic DNA was isolated using a Qiagen DNeasy Blood and Tissue Kit (#69504) and delivered to the Duke University Sequencing and Genomic Technologies Shared Resource where it was sequenced using a FLO-Min112 Flow Cell on a GridIon sequencer. Approximately 4.13 Gb of sequences were produced. Sequence was assembled using minimap2 (Li, H., 2018) and visualized using Tablet (Milne et al., 2013) and changes to the tilt genome were verified by eye for the ~30 kb region shown in Figure 1C . Sequence Reads deposited in SRA with a BioSample accession of SAMN41913838 : Run SRR29467516 from the Sequence Read Archive (SRA). Available at https://trace.ncbi.nlm.nih.gov/Traces/?view=run_browser&acc=SRR29467516
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ayala-Camargo A Anderson AM Amoyel M Rodrigues AB Flaherty MS Bach EA 2013823 JAK/STAT signaling is required for hinge growth and patterning in the Drosophila wing disc.Dev Biol 38220012-160641342610.1016/j.ydbio.2013.08.01623978534 PMC 3795806 · doi ↗ · pubmed ↗
- 2Bridges, C. B., & Morgan, T. H. (1919). Contributions to the genetics of Drosophila melanogaster . Carnegie Institute of Washington Publication, 278 , 123–304.
- 3Bridges, C. B., & Morgan, T. H. (1923). The third-chromosome group of mutant characters of Drosophila melanogaster . Carnegie Institute of Washington Publication, 327 , 1–251.
- 4Carrasco-Rando M Tutor AS Prieto-Sánchez S González-Pérez E Barrios N Letizia A Martín P Campuzano S Ruiz-Gómez M 2011721 Drosophila araucan and caupolican integrate intrinsic and signalling inputs for the acquisition by muscle progenitors of the lateral transverse fate.P Lo S Genet 771553-7390 e 1002186 e 100218610.1371/journal.pgen.100218621811416 PMC 3141015 · doi ↗ · pubmed ↗
- 5Crozatier Michèle Glise Bruno Vincent Alain 2004101 Patterns in evolution: veins of the Drosophila wing Trends in Genetics 20100168-952549850510.1016/j.tig.2004.07.01315363904 · doi ↗ · pubmed ↗
- 6Georgiev P Kozycina M 199621 Interaction between mutations in the suppressor of Hairy wing and modifier of mdg 4 genes of Drosophila melanogaster affecting the phenotype of gypsy-induced mutations.Genetics 14220016-673142543610.1093/genetics/142.2.4258852842 PMC 1206977 · doi ↗ · pubmed ↗
- 7Gerland TA Sun B Smialowski P Lukacs A Thomae AW Imhof A 2017216 The Drosophila speciation factor HMR localizes to genomic insulator sites.P Lo S One 122e 0171798 e 017179810.1371/journal.pone.017179828207793 PMC 5312933 · doi ↗ · pubmed ↗
- 8Gomez-Skarmeta JL Diez del Corral R de la Calle-Mustienes E Ferré-MarcóD Modolell J 199645 Araucan and caupolican, two members of the novel iroquois complex, encode homeoproteins that control proneural and vein-forming genes.Cell 8510092-86749510510.1016/s 0092-8674(00)81085-58620542 · doi ↗ · pubmed ↗