The lack of homozygotes with a large deletion encompassing SPAG1 and POLR2K in primary ciliary dyskinesia patients suggests the lethal effect of the loss of POLR2K protein
Alicja Rabiasz, Monika Drobna-Śledzińska, Patrycja Kaźmierczak, Michał Witt, Ewa Ziętkiewicz

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
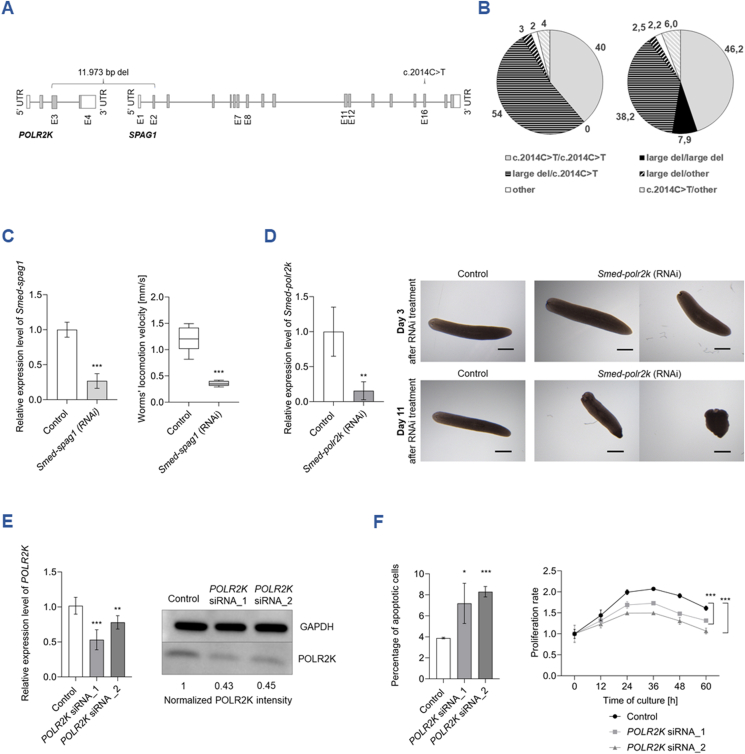 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCystic Fibrosis Research Advances · Neonatal Respiratory Health Research · Respiratory Support and Mechanisms
SPAG1 (sperm-associated antigen 1) is one of over 50 genes whose pathogenic variants underlie primary ciliary dyskinesia (PCD; OMIM244400), an inherited disorder affecting the function of motile cilia,1 highly conserved organelles protruding from the surface of eukaryotic cells. Pathogenic SPAG1 variants impair the assembly of dynein arms, essential elements of cilia. In a cohort of almost 300 affected individuals with the genetic bases of PCD explained during our long-term studies, pathogenic variants in SPAG1 were found in 60 unrelated patients, making it one of the most frequently involved genes in the Polish PCD population. Two predominant, previously reported variants1^,^2 were a nonsense mutation c.2014C>T in exon 16, found in both homozygotes and in compound heterozygotes, and a large 11,973 bp deletion encompassing parts of SPAG1, and of the upstream POLR2K (RNA polymerase II, I and III subunit K). c.2014C>T was found in 27 homozygotes and 31 compound heterozygotes, of whom 28 had the large deletion as the second allele; the large deletion was found in two more compound heterozygotes, accompanied in trans by rare pathogenic variants (Fig. 1A and Tables S1 and 2). Similar to previous reports,1^,^2 the large deletion was repetitively found in compound heterozygosity, but no homozygotes were identified.Figure 1. The elucidation of the role of POLR2K in the pathogenesis of primary ciliary dyskinesia (PCD). (A) Scheme of the localization of pathogenic variants detected in the analyzed group of 60 SPAG1-PCD patients. Two frequent deletion are indicated above the gene scheme: 11,973 bp del: a large 11,973 bp deletion [NM_005034.4:c.61 + 201_NM_003114.5:c.140 + 1169]del, encompassing exons 1 and 2 of SPAG1 (sperm-associated antigen 1) and exons 3 and 4 of POLR2K (RNA polymerase II, I and III subunit K) located upstream of SPAG1 in chromosome 8q22.2 (indicated by a bracket); c.2014C>T in exon 16 (NM_003114.5:c.2014C>T; NP_003105.2:p.Gln672∗). SPAG1 exons with pathogenic variants are indicated by numbers. (B) Distribution of SPAG1 genotypes in the combined group of 103 unrelated Caucasian SPAG1-PCD patients. Left panel: The count of observed genotypes. Right panel: The expected number of genotypes calculated assuming Hardy–Weinberg equilibrium. The expected number of the deletion homozygotes in the joint group of 103 SPAG1-PCD patients raised to eight; with none observed, the difference between observed and expected was statistically significant (p = 0.0066). (C) Analysis of the impact of Smed-spag1 knockdown on S. mediterranea movement. Left: Effectiveness of the dsRNA-mediated Smed-spag1 knockdown. DsRNA-mediated knockdown reduced the relative expression level of Smed-spag1 to 27% compared with control worms treated with dsRNA targeting eGFP (measured by quantitative reverse transcription PCR one day after the last administration of dsRNA; n = 4; ∗∗∗p < 0.001). Right: The speed of planarians' locomotion at day 2 after the last administration of dsRNA (1.189 ± 0.371 and 0.354 ± 0.065 mm/s, in controls and Smed-spag1-silenced worms, respectively). Plots show the mean value of biological replicates (n = 10); error bars indicate standard deviation; ∗∗∗p < 0.001. (D) Analysis of the impact of Smed-polr2k knockdown on S. mediterranea's phenotype. Left: Effectiveness of the dsRNA-mediated Smed-polr2k knockdown. Quantitative reverse transcription PCR (measured one day after the last administration of dsRNA) showed a decrease in Smed-polr2k expression level to 14% compared with control worms; plots show the mean value of biological replicates (n = 4); error bars indicate standard deviation; ∗∗p < 0.01. Right: Examples of worm's phenotype abnormalities (deformation of body shape, tissue lysis) after Smed-polr2k knockdown, compared with a control (upper and lower panel: day 3 and 11 after last dsRNA feeding, respectively); the defects finally lead to worms' death. The first changes in the worm's phenotype were observed 3 days after the last dsRNA feeding and affected 54% of the worms; it gradually affected all Smed-polr2k-silenced worms. Scale bar: 1 mm. (E) Efficiency of the siRNAs-mediated POLR2K knockdown in human cells, 48 h post transfection. Left: Quantitative reverse transcription PCR analysis of POLR2K mRNA level. POLR2K siRNA_1 and POLR2K siRNA_2 reduced the relative expression level of POLR2K to ∼54% and ∼78%, respectively (non-targeting siRNA was used as a negative control). Quantitative reverse transcription PCR plots show the mean value of biological replicates (n = 5); error bars indicate standard deviation; ∗∗p < 0.01, ∗∗∗p < 0.001. Right: Western blot analysis of POLR2K; at the protein level, the knockdown efficiency was comparable for both siRNAs; normalized to GAPDH. (F) Analysis of the impact of POLR2K knockdown on vital functions in human cells. Left: Apoptosis assay performed using flow cytometry. Percentage of apoptotic cells: 7.2% and 8.3% 48 h after transfection with POLR2K siRNA_1 and POLR2K siRNA_2, respectively, compared with 3.9% in the negative control. Plots show the mean value of biological replicates (n = 3); error bars indicate standard deviation; ∗p < 0.05, ∗∗∗p < 0.001. Right: Cell proliferation rate (assessed using colorimetric assay) was calculated as the fold change in OD450 for each examined time point in reference to the starting point (0 h); biological replicates, n = 3; ∗∗∗p < 0.001.Figure 1
In our group of 60 SPAG1-PCD patients, ∼4 homozygotes with the large deletion are expected under Hardy–Weinberg equilibrium; however, none was observed. This difference in the predicted and observed distribution of genotypes was not statistically significant (chi-square, p = 0.1663). To achieve the statistical power, we added data for 18 Caucasians reported earlier1^,^2 (8 compound heterozygotes carrying the large deletion), and for 25 SPAG1-PCD German patients (6 c.2014C>T homozygotes and 19 heterozygotes for c.2014C>T and the large deletion; Heymut Omran, personal communication). The expected number of the large deletion homozygotes in the joint group of 103 patients raised to eight (Fig. 1B); with none observed, the unbalanced distribution of the genotypes was statistically significant (p = 0.006).
The lack of large deletion homozygotes in the combined group of 103 patients suggested a seriously deleterious effect (presumably lethality) of this genotype. Given that the large deletion encompasses the C-terminal part of POLR2K in addition to exons 1 and 2 of SPAG1, we hypothesized that this was due to the lack of functional POLR2K rather than SPAG1 protein. POLR2K represents one of the smallest subunits of RNA polymerase II, shared with two other RNA polymerases; in consequence, POLR2K is involved in the RNA processing machinery and improves RNA polymerase III pre-initiation complex assembly.3 To confirm our hypothesis, we examined the effect of RNA interference-mediated knockdown of POLR2K at two levels: the whole organism (ciliated flatworm, Schmidtea mediterranea), and cellular (human cell line).
S. mediterranea is used as a model organism to study the functionality of embryonic-lethal genes in the adult body and cilia-related genes (worms' locomotion depends on motile cilia covering the ventral side of their body). To compare the phenotypes caused by the deficiency of Smed-spag1 and Smed-polr2k in S. mediterranea, we first performed a Smed-spag1 knockdown. The effectiveness of gene silencing was confirmed using quantitative reverse transcription PCR (Fig. 1C-left). The worms upon knockdown displayed no morphological defects, but moved three times slower than controls, using characteristic inchworming (Fig. 1C-right; supplementary movies 1 and 2).
Supplementary video related to this article can be found at https://doi.org/10.1016/j.gendis.2025.101535
The following are the supplementary data related to this article:VideoS12VideoS1VideoS23VideoS2VideoS34VideoS3
In Smed-*polr2k-*knockdown worms, the relative expression level was also significantly reduced (Fig. 1D-left). In contrast to Smed-spag1-silencing, the knockdown of Smed-polr2k led to severe defects of planarian bodies, including tissue lysis and head regression, which ultimately led to worms' death (Fig. 1D-right). Furthermore, despite the enormous regenerative capacity of planarians, Smed-polr2k deficiency resulted in disruption of the regeneration process (the lack of worms' regrowth after cutting) (Fig. S1). On the other hand, until the point of tissue lysis, no worms' motility impairment was observed (supplementary movie 3), indicating the lack of Smed-polr2k involvement in motile cilia function.
To better understand the phenotype observed in worms and to relate the obtained results to humans, the effect of POLR2K knockdown on the vital cell functions was assessed in the human HEK293T cell line transfected in parallel with two siRNAs targeting different regions of POLR2K exon 3. Quantitative reverse transcription PCR and western blotting confirmed the effectiveness of the gene knockdown at both mRNA and protein levels, respectively (Fig. 1E). POLR2K knockdown resulted in a gentle but statistically significant increase in the number of apoptotic cells (Fig. 1F-left). The cell proliferation was significantly decreased in the POLR2K-silenced cells compared with the control (Fig. 1F-right).
In the presented study, we examined the hypothesis that the discrepancy between the observed and expected distribution of haplotypes in SPAG1-PCD patients (the lack of homozygotes with the large deletion encompassing parts of SPAG1 and POLR2K), reflected a highly deleterious, possibly lethal effect of the loss of a functional POLR2K rather than of SPAG1 protein. A similarly unbalanced distribution of genotypes has been described for PMM2 gene4 (zero observed versus eight expected p.R141H homozygotes in 54 Caucasian patients with carbohydrate-deficient glycoprotein syndrome type 1A). The authors concluded that this distribution was probably caused by selection against the homozygous R141H.4 It should be emphasized that the frequency of homozygous pathogenic variants introducing premature STOP codon located in SPAG1 exon 16 (c.2014C>T) was even higher than expected under equilibrium, indicating the lack of selection against a homozygous deficiency of the wild-type SPAG1.
To provide a molecular context to our observation, we analyzed the impact of POLR2K silencing on phenotypes at the whole organism (planarians) and cellular levels. The dsRNA-mediated knockdown of Smed-spag1 in S. mediterranea did not affect the viability of the organisms, but as expected, impaired worms' movement, confirming the evolutionary conserved role of SPAG1 in the motile cilia function (previously demonstrated using a vertebrate model, zebrafish1). In contrast, silencing of Smed-polr2k resulted in a much more severe phenotype and suggested that Smed-polr2k played an essential role in maintaining body homeostasis by regulating cell turnover, probably through the impact on planarian stem cell proliferation. Our observations are in line with those in other studies where deleterious effects of other essential gene knockdowns on S. mediterranea phenotype have been reported. For example, RNA interference-mediated knockdown of Argonaute RISC catalytic component 2 (Smed-ago2)5 led to regeneration defects or tissue regression in worms, ultimately causing their death, demonstrating that Smed-ago2 is crucial for tissue homeostasis and regeneration and acts through a regulation of adult stem cell proliferation.5
Our results of siRNA-mediated POLR2K knockdown in the human cell culture shed light on the mechanisms of defects observed in the planarian model. They confirmed the essential role of POLR2K in the regulation of vital cell functions, manifested by decreased cell proliferation and increased apoptosis upon its knockdown.
Despite the repetitive reports suggesting POLR2K's importance in many cellular processes (additional discussion), no direct experimental proof of phenotype changes after POLR2K knockdown has been published. Our findings confirm that POLR2K is necessary for the functioning of living organisms and suggest that its influence on cell turnover is exerted through the regulation of cell proliferation and apoptosis. Moreover, our study provides evidence that the essential role of POLR2K in vital cell functions is evolutionarily conserved. Finally, these results, together with the earlier studies regarding POLR2K as a hub gene involved in many essential interactions, support our starting hypothesis that the lack of homozygotes of the large deletion involving SPAG1 and POLR2K genes is caused by the lethal impact of the POLR2K deficiency; at the same time, it indicates that POLR2K is not directly involved in the PCD pathogenesis. Our findings suggest that potential mRNA-based therapy for SPAG1-PCD patients with large deletion should be designed to restore SPAG1, and not POLR2K function.
Ethics declaration
The diagnostics were conducted according to the recommendation of the World Medical Association. Written informed consent was obtained from the participating subjects or their guardians. Procedures representing part of the diagnostic scheme were approved by the institutional committee (the Ethics Committee of the Medical University in Poznań, 960/11, 435/13, and 381/22).
Funding
This research was funded by the 10.13039/501100004442National Science Centre, Poland (No. 2018/29/N/NZ5/00810 to A.R.; 2018/31/B/NZ2/03248 to E.Z.) and by the Institute of Human Genetics PAS, Poland (Minigrant No. 2021/01 to A.R.).
CRediT authorship contribution statement
Alicja Rabiasz: Writing – review & editing, Writing – original draft, Visualization, Validation, Methodology, Investigation, Funding acquisition, Formal analysis, Data curation, Conceptualization. Monika Drobna-Śledzińska: Writing – review & editing, Methodology, Investigation, Formal analysis. Patrycja Kaźmierczak: Writing – review & editing, Investigation. Michał Witt: Writing – review & editing, Supervision. Ewa Ziętkiewicz: Writing – review & editing, Writing – original draft, Supervision, Investigation, Funding acquisition, Formal analysis, Data curation, Conceptualization.
Data availability
The data that support the findings of this study are available from the corresponding authors upon reasonable request.
Conflict of interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Knowles M.R.Ostrowski L.E.Loges N.T.Mutations in SPAG 1 cause primary ciliary dyskinesia associated with defective outer and inner dynein arms Am J Hum Genet 93420137117202405511210.1016/j.ajhg.2013.07.025PMC 3791252 · doi ↗ · pubmed ↗
- 2Djakow J.KramnáL.DušátkováL.An effective combination of Sanger and next generation sequencing in diagnostics of primary ciliary dyskinesia Pediatr Pulmonol 51520164985092622829910.1002/ppul.23261 · doi ↗ · pubmed ↗
- 3Li Q.Xu Z.Fang F.Shen Y.Lei H.Shen X.Identification of key pathways, genes and immune cell infiltration in hypoxia of high-altitude acclimatization via meta-analysis and integrated bioinformatics analysis Front Genet 142023105537210.3389/fgene.2023.1055372 PMC 1008002337035734 · doi ↗ · pubmed ↗
- 4Matthijs G.Schollen E.Van Schaftingen E.Cassiman J.J.Jaeken J.Lack of homozygotes for the most frequent disease allele in carbohydrate-deficient glycoprotein syndrome type 1A Am J Hum Genet 6231998542550949726010.1086/301763 PMC 1376957 · doi ↗ · pubmed ↗
- 5Li Y.Q.Zeng A.Han X.S.Argonaute-2 regulates the proliferation of adult stem cells in planarian Cell Res 21122011175017542189418910.1038/cr.2011.151PMC 3357986 · doi ↗ · pubmed ↗