Seroprevalence of Rift Valley and Crimean-Congo Hemorrhagic Fever Viruses, Benin, 2022–2023
Nadine Olk, Anges Yadouleton, Olga Quenum, Stephane Sohou, Aime Goundote, Grace Aho Glele Rodrigue, Blaise Guezo-Mevo, Sonia Bedie, Michael Nagel, Petra Emmerich, Benjamin Hounkpatin, Jan Felix Drexler

TL;DR
A study in Benin found low antibody prevalence for two viral diseases but highlights issues with serologic tests in malaria-endemic areas.
Contribution
The study identifies potential false positives in serologic testing due to cross-reactivity with malaria antibodies in endemic regions.
Findings
No patients tested positive for Rift Valley or Crimean-Congo hemorrhagic fever viruses via PCR.
1.1% and 0.3% of patients had IgG antibodies for Rift Valley and Crimean-Congo viruses, respectively.
Malaria antibodies may cause false positives in serologic tests due to cross-reactivity with viral antigens.
Abstract
We screened 650 febrile patients from Benin for Rift Valley fever and Crimean-Congo hemorrhagic fever viruses during 2022–2023. None were positive by reverse transcription PCR; 1.1% and 0.3%, respectively, had virus-specific IgG. False-positive results from malaria-associated antibodies likely reacting with histidine-tagged viral antigens mandate careful validation of serologic tests in malaria-endemic regions.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
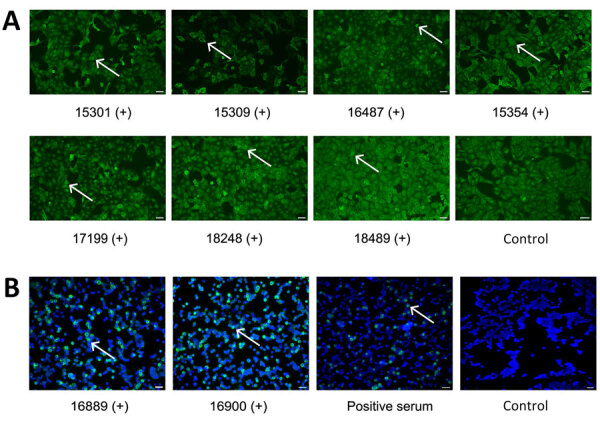 Figure 1
Figure 1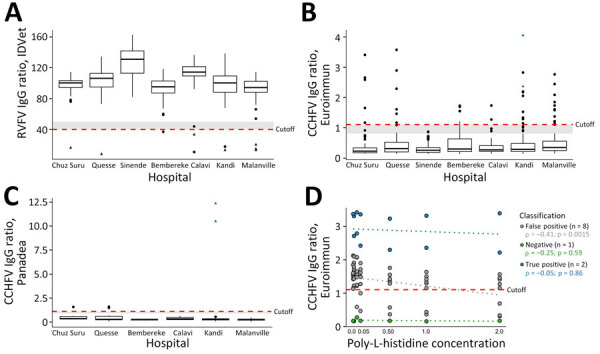 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsViral Infections and Vectors · Viral Infections and Outbreaks Research · Mosquito-borne diseases and control
Rift Valley fever virus (RVFV; family Phenuiviridae) and Crimean-Congo hemorrhagic fever virus (CCHFV; family Nairoviridae) are arthropodborne viruses endemic to Africa and the Arabian Peninsula (1,2) and high-priority pathogens that can cause lethal hemorrhagic fever (2–4) (https://www.who.int/publications/m/item/WHO-BS-2023-2449). In West Africa, RVFV and CCHFV are considered endemic in Senegal and Mauritania (1,2); regional circulation seems likely in Guinea, Burkina Faso, Ghana, and Nigeria (2). In Benin, CCHFV antibodies were reported in humans in 1981, but RVFV and CCHFV epidemiology remains unknown (1,2). Both RVFV and CCHFV infect diverse animals reared as livestock (3). Benin has been undergoing changes in traditional cattle farming, including increased herd sizes and sedentarization (5), which may intensify RVFV and CCHFV circulation. We collected serum samples for routine diagnostic examinations for RVFV and CCHFV in 7 hospitals located across ≈700 km and 3 ecozones in Benin (Appendix Table, Figure).
We investigated serum samples from 650 febrile patients (mean age 26.7 [interquartile range 18–34] years; 70.3% female, 29.7% male) who were seen during December 2022–January 2023. We analyzed samples for acute RVFV and CCHFV infection using PCR-based methods and had no positive results (Appendix). However, we detected IgG by using commercially available ELISA kits (RVFV, competitive ELISA; ID.Vet, https://bioadvance.life/en/id-vet-2; CCHFV, indirect ELISA; Euroimmun, https://www.euroimmun.com) with viral nucleoproteins as antigens. We confirmed CCHFV ELISA results by using a CCHFV immune complex capture IgG ELISA (Panadea Diagnostics, https://www.panadea-diagnostics.com) and RVFV and CCHFV ELISA results by indirect IgG immunofluorescence assays (IFAs).
The competitive RVFV IgG ELISA was positive in 10 (1.5%, 95% CI 0.6%–2.5%) samples; 7 were positive by RVFV IFA with high endpoint titers of 1:1,000–12,500 serum dilution (Figure 1, panel A; Figure 2, panel A; Appendix Table). Differential test sensitivity might cause discordant ELISA and IFA results, but ELISA reactivity was not weaker in IFA-negative samples compared with IFA-positive samples (p = 0.83 by Mann-Whitney U test). By indirect ELISA, 40 (6.1%, 95% CI 4.3%–8.0%) samples were positive for CCHFV, but only 5 samples tested positive by immune capture ELISA (Figure 2, panels B, C). Of those 5 samples, we confirmed 2 by CCHFV IFA, with low endpoint titers of 1:20–1:80 (Figure 1, panel B; Appendix Table). IFA-negative samples showed low reactivity in the immune capture ELISA, suggesting differential sensitivity or need to adjust ELISA positivity thresholds (Figure 2, panel C).
Discrepancies among detection rates of the 2 CCHFV ELISA tests and IFA were surprising. Unspecific ELISA reactivity can occur; for example, malaria or herpes virus infection might cause unspecific B-cell stimulation (6,7). Antibodies against Plasmodium falciparum parasites’ histidine-rich proteins occur in ≈25% of people in malaria-endemic areas and decrease sensitivity of rapid diagnostic tests (8). During in vitro antigen production for serologic tests, <6 histidine residues are frequently added to expression constructs for protein purification (9). Increasing externally added histidine concentrations led to significantly decreased CCHFV indirect ELISA reactivity in potentially false-positive samples (ρ = 0.41; p = 0.0015) (Figure 2, panel D). In contrast, reactivities of likely true-positive samples (i.e., confirmed by IFA or immune capture ELISA) and likely true-negative samples were not affected by incremental histidine concentrations. Those data substantiated that antibodies potentially elicited by previous or acute Plasmodium infections targeting histidine-rich epitopes might have interacted with likely histidine-tagged indirect ELISA antigens to cause the observed reactivity pattern, including multiple likely false-positive test results. Other histidine-rich immunogens might also have elicited potentially cross-reactive antibodies, yet malaria-associated immune responses remain the most plausible explanation because of the abundance of malaria in sub-Saharan Africa and a similar rate of potentially false-positive COVID-19 results in a previous serologic study (6). Although the competitive RVFV ELISA and immune complex capture–based CCHFV ELISA might be more specific than indirect ELISA formats, we only considered IFA-positive results for a conservative assessment of the RVFV antibody detection rate of 1.1% (95% CI 0.3%–1.9%; n = 7/650) and of the CCHFV antibody detection rate of 0.3% (95% CI −0.1% to 0.7%; n = 2/650) (Appendix Table).
Our serologic data thus substantiated circulation of RVFV and CCHFV in Benin (Appendix Table), albeit at relatively low rates that are largely comparable to neighboring countries (1,2). Livestock rearing in Benin is transitioning to partly sedentary systems with larger cattle herds (5), which highlights the need to continuously monitor RVFV and CCHFV circulation in humans and cattle (10) and support with robust serologic tests validated for specificity in malaria-endemic regions and direct detection of pathogens in arthropod vectors, such as Culex and Aedes mosquitoes for RVFV and Hyalomma ticks for CCHFV.
The main limitation of our study is that it is a nonrepresentative sample. However, including febrile patients from 7 hospitals across 3 ecozones provides broad geographic and ecologic coverage (1).
Beyond surveillance, strategies for future vaccination of livestock and humans will benefit from robust epidemiologic data on RVFV and CCHFV to efficiently use resources across sub-Saharan Africa. Serologic tests relying on tag-free protein production, alternative tags, and careful validation of histidine-tagged antigens for specificity are mandatory for use of antibody tests in malaria-endemic regions.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Tinto B, Quellec J, Cêtre-Sossah C, Dicko A, Salinas S, Simonin Y. Rift Valley fever in West Africa: A zoonotic disease with multiple socio-economic consequences. One Health. 2023;17:100583. 10.1016/j.onehlt.2023.10058337664171 PMC 10474305 · doi ↗ · pubmed ↗
- 2Temur AI, Kuhn JH, Pecor DB, Apanaskevich DA, Keshtkar-Jahromi M. Epidemiology of Crimean-Congo hemorrhagic fever (CCHF) in Africa—underestimated for decades. Am J Trop Med Hyg. 2021;104:1978–90. 10.4269/ajtmh.20-141333900999 PMC 8176481 · doi ↗ · pubmed ↗
- 3Hawman DW, Feldmann H. Crimean-Congo haemorrhagic fever virus. Nat Rev Microbiol. 2023;21:463–77. 10.1038/s 41579-023-00871-936918725 PMC 10013989 · doi ↗ · pubmed ↗
- 4Petrova V, Kristiansen P, Norheim G, Yimer SA. Rift valley fever: diagnostic challenges and investment needs for vaccine development. BMJ Glob Health. 2020;5:e 002694. 10.1136/bmjgh-2020-00269432816810 PMC 7437696 · doi ↗ · pubmed ↗
- 5Houessou SO, Dossa LH, Diogo RVC, Houinato M, Buerkert A, Schlecht E. Change and continuity in traditional cattle farming systems of West African Coast countries: a case study from Benin. Agric Syst. 2019;168:112–22. 10.1016/j.agsy.2018.11.003 · doi ↗
- 6Yadouleton A, Sander AL, Moreira-Soto A, Tchibozo C, Hounkanrin G, Badou Y, et al. Limited specificity of serologic tests for SARS-Co V-2 antibody detection, Benin. Emerg Infect Dis. 2021;27:233–7. 10.3201/eid 2701.20328133261717 PMC 7774555 · doi ↗ · pubmed ↗
- 7Piche-Ovares M, García MP, Moreira-Soto A, Figueroa-Romero MD, Merino-Sarmiento NS, Marcelo-Ñique AI, et al. Rio Mamore hantavirus endemicity, Peruvian Amazon, 2020. Emerg Infect Dis. 2024;30:2532–43. 10.3201/eid 3012.24024939592268 PMC 11616641 · doi ↗ · pubmed ↗
- 8Ho MF, Baker J, Lee N, Luchavez J, Ariey F, Nhem S, et al. Circulating antibodies against Plasmodium falciparum histidine-rich proteins 2 interfere with antigen detection by rapid diagnostic tests. Malar J. 2014;13:480. 10.1186/1475-2875-13-48025481825 PMC 4295572 · doi ↗ · pubmed ↗