Evaluation of the Expression of Virulence Factors of V. cholerae After Interaction With the Human Colon Adenocarcinoma (Caco-2) Cell Line
Mohammadreza Kheradmand, Masoumeh Saberpour, Bita Bakhshi, Mahboube Bahroudi

TL;DR
This study shows how V. cholerae changes its gene expression when interacting with human colon cells, revealing differences between two biotypes that could help in managing cholera.
Contribution
The study compares gene expression changes in two V. cholerae biotypes after interaction with Caco-2 cells, highlighting biotype-specific virulence responses.
Findings
The classical biotype showed a 6-fold increase in ctxAB gene expression compared to the El Tor biotype's 2-fold increase.
The hlyA gene expression increased 13-fold in the classical biotype and 7-fold in the El Tor biotype after Caco-2 interaction.
TcpA gene expression increased significantly in the classical biotype but not in the El Tor biotype when interacting with Caco-2 cells.
Abstract
Cholera remains a global challenge, and understanding how V. cholerae adapts to environmental condition is essential for innovating new management strategies. This research aims to examine the expression of ctxAB, tcpA, and hlyA genes in V. cholerae (El Tor and classical biotypes) after interaction with Caco-2 cells compared to brain heart infusion (BHI) broth. After assaying of viability of Caco-2 cells against V. cholerae at multiplicity of infection (MOI) of 10, 20, 50, and 80, the number of bacteria attached to Caco-2 cells was determined using the adhesion assay. To conduct a valid comparison, an equivalent number of bacteria that attached to Caco-2 cells was inoculated into BHI broth. The expression of ctxAB, tcpA, and hlyA genes in V. cholerae (El Tor and classical biotypes) was assayed using the real-time PCR method. After interaction of Caco-2 cells with V. cholerae, the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
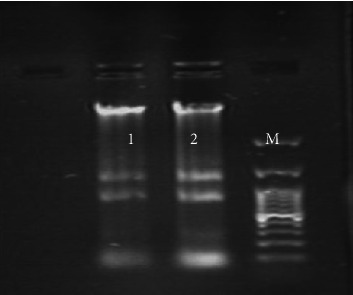 Figure 1
Figure 1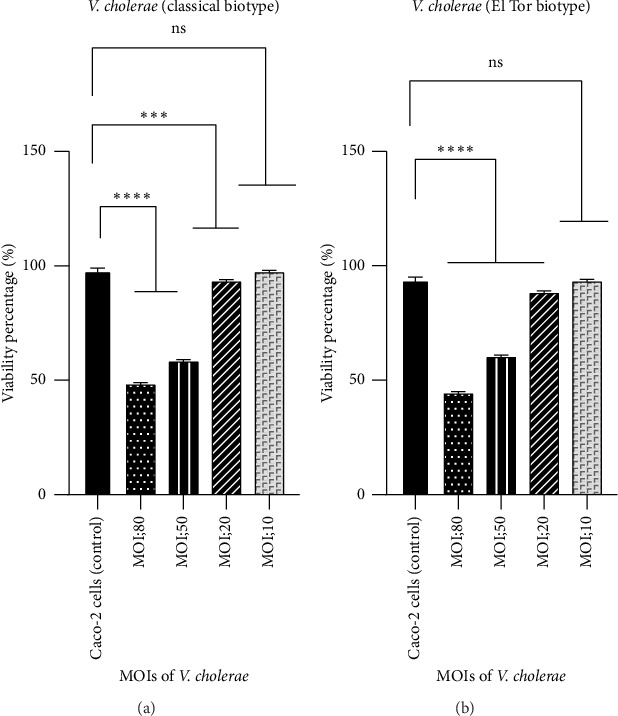 Figure 2
Figure 2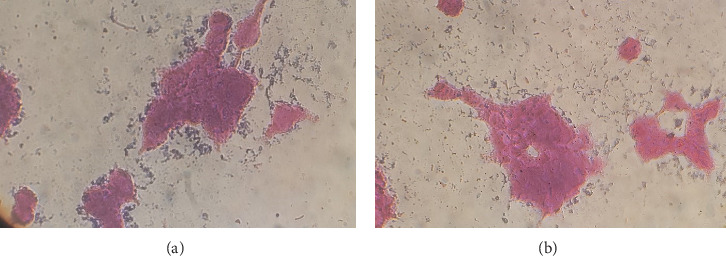 Figure 3
Figure 3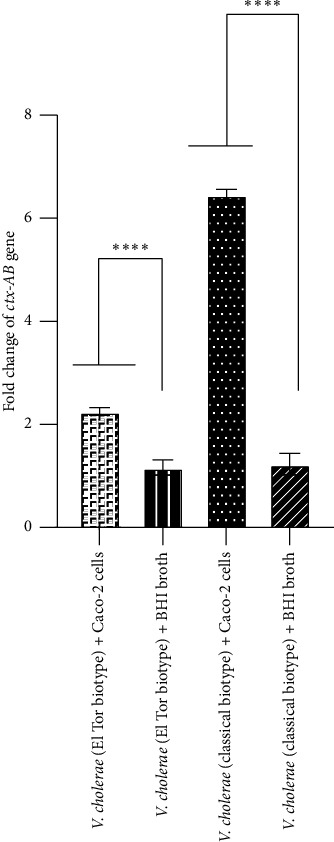 Figure 4
Figure 4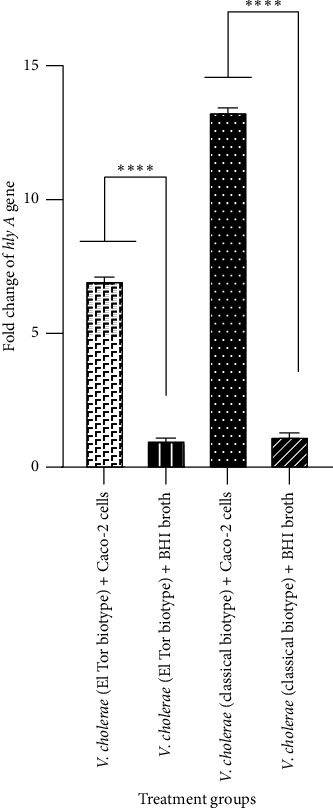 Figure 5
Figure 5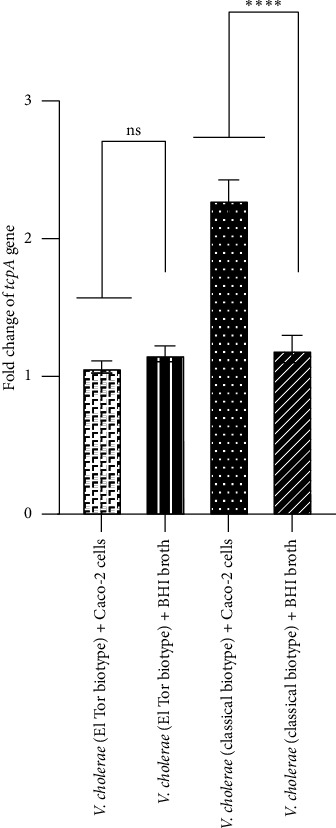 Figure 6
Figure 6- —Tarbiat Modares University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsVibrio bacteria research studies · Bacterial biofilms and quorum sensing · Escherichia coli research studies
1. Introduction
Cholera is a major health challenge with high mortality rates worldwide, especially in developing nation [1]. Cholera is a disease characterized by severe watery diarrhea caused by Vibrio (V). cholerae. It exists in contaminated water and displays diverse pathogenic behaviors under various environmental conditions [2]. After exposure of V. cholerae to unfavorable environments, it modifies its pathogenic behavior by regulating the expression of virulence factors, thereby adapting to adverse conditions and demonstrating increased resistance [3]. So, analyzing of the behavior of this pathogen in different environments and monitoring the expression of its virulence factors help us develop effective strategies for managing this organism. V. cholerae is a gram-negative bacterium with many virulence factors including cholera toxin (CT) (ctxAB), toxin-coregulated pilus (tcpA), hemolysin A (hlyA), zonula occludens toxin (Zot), accessory cholera enterotoxin (Ace), flagella, outer membrane protein U (OmpU), haemagglutinin/protease (HapA), and ToxR-activated gene- A (TagA). The main virulence factors of this bacterium are CT (ctxAB) that produced by a lysogenic phage (CTXΦ), and it alters signaling transduction pathways in the host and leads to diarrhea, toxin-regulated pilus (tcpA) that is a major adhesion factor for colonization of the intestinal epithelium, and hemolysin-encoded by the hlyA gene [4]. These factors are contributed to the high resistance of bacteria in diverse environmental conditions, including aqueous and nonaqueous environments, and adverse environmental conditions such as nutrient deficiencies, alterations in acidity, and fluctuations in temperature [5]. V. cholerae regulates its gene expression in response to adverse conditions. This adaptive process enhances effective colonization, survival, and virulence [3]. Considering the important role of key virulence factors in the persistence of V. cholerae across different environments, assessing the interaction of V. cholerae with various environments is essential. In this study, two different environments including Caco-2 cells and brain heart infusion (BHI) broth medium were used. Caco-2 cells derived from human colon adenocarcinomas have the ability to spontaneously differentiate and form a monolayer in cell culture. After achieving maximum differentiation, these cells display the morphological and functional characteristics of enterocytes, so Caco-2 cells serve as an ideal model system for further evaluation of the pathogenesis of V. cholerae [6]. Moreover, BHI broth medium provides an ideal environment for V. cholerae due to its rich nutrient composition. Few studies exist in this field. For example, interactions of Caco-2 cells with Clostridium perfringens significantly increased the expression of the Clostridium perfringens enterotoxin (Cpe) gene compared to LB broth medium [7]. Moreover, some researchers focused on evaluating the impact of essential minerals, such as zinc (Zn), selenium (Se), and manganese (Mn) in the Caco-2 cell environment, on the expression of virulence factors of V. cholerae [8]. So far, no research has assessed the expression of virulence genes V. cholerae including ctxAB, tcpA, and hlyA following interaction with Caco-2 cells. Therefore, this study aimed to evaluate of the expression of ctxAB, tcpA, and hlyA genes of V. cholerae (El Tor and classical biotypes) in two conditions of Caco-2 cells (similar to the intestinal environment) and BHI broth (nutrient-rich medium) environments. It will focus on assessing how V. cholerae adapts to Caco-2 cell environments and identifying which pathogenic factors of V. cholerae are effective in adapting to this environment.
2. Materials and Methods
2.1. Growth Condition of V. cholerae
In the current study, two strains of V. cholerae (a classical biotype; ATCC 14035 and a clinical El Tor biotype) were obtained from the laboratory archive in the Microbiology Department of Tarbiat Modares University. These strains were previously confirmed using both phenotypic and molecular techniques. After growing V. cholerae on the surface of thiosulfate citrate bile salts sucrose (TCBS) agar (Merck, Germany), a single colony from each of the strain was initially cultured on BHI agar (Merck, Germany), incubated at 37°C for 24 h, and subsequently subcultured in BHI broth (Merck, Germany) at 37°C for an incubation time of 24 h. For preparing of the bacterial suspension, a volume of 300 μL from the bacterial suspension was transferred into fresh BHI broth medium and incubated at 37°C for 4 h. After 4 h, the concentration of the resulting bacterial suspension was evaluated by measuring its optical density (OD) at 625 nm. OD of the bacterial suspension was determined using the 0.5 McFarland standard, equivalent to 0.1, containing 1.5 × 10^8^ colony forming unit (CFU)/mL. This number of bacteria from each of the two strains was used to inoculate Caco-2 cells [9].
2.2. Preparation of Caco-2 Cells
Caco-2 cells were purchased from the National Center for Genetic and Biological Resources of Iran (IBRC10094). The cells were cultured in a T75 flask (SPL, Korea) containing Dulbecco's Modified Eagle Medium (DMEM) (Gibco, USA) enriched with 10% fetal bovine serum (FBS) (Gibco, USA), 1% antibiotics (penicillin: 100 U/mL and streptomycin: 100 mg/mL) (Gibco, USA), and 2 mM L-glutamine (DNA Biotech, Iran). The cells were then incubated with 5% CO2 at 37°C for 48 h. When the cells reached 80%–90% confluency, they were subcultured into a new flask [10].
2.3. Evaluation of Cytotoxicity of V. cholerae on Caco-2 Cells
Persistence of Caco-2 cells was separately assessed after exposure to four multiplicities of infection (MOIs; 10, 20,50, and 80) of the V. cholerae strain (classical biotype; ATCC14035) and the V. cholerae strain (El Tor biotype) using MTT solution (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide) (DNA Biotech, Iran). Briefly, 10^4^ cells were seeded in each well of a 96-well culture plate (SPL, Korea), and the plates were incubated overnight at 37°C with 5% CO2. After the cells reached 80% filling, each well was washed three times with sterile phosphate-buffered saline (PBS) 1X. Subsequently, classical and El Tor biotypes of V. cholerae at MOIs 10, 20,50, and 80 were added to the cells and incubated for 2 h at 37°C with 5% CO2 [11]. In the control cells, the medium was not changed. After this time, the medium was aspirated from each well and replaced with DMEM medium containing 10% FBS. Then, the cells were incubated for 3 h at 37°C with 5% CO2. Following this time, the medium was eliminated, and 100 μL of MTT reagent was transferred to each well. Next, the cells were placed in a 37°C incubator containing 5% CO2 for 4 h in the dark. After 4 h, the MTT regent was removed from the cells. Finally, 100 μL of dimethyl sulfoxide (DMSO) (Sigma Aldrich, USA) was transferred to each well to dissolve the formazan. Then, OD of purple formazan was measured at 570 nm using an enzyme-linked immunosorbent assay (ELISA) reader (800 TS, BioTek, Winooski, Vermont, USA). The experiments were repeated three times, and the data are presented as the mean ± SD [10].
2.4. Evaluation Attachment of V. cholerae to Caco-2 Cells
For assessment of attachment of V. cholerae to Caco-2 cells, the 10^5^ cells seeded into a 12-well culture microplate (SPL, Korea) enriched to 10% FBS without antibiotics. After 24 h, when the cells reached to 80%–90% confluency, the medium was removed from each well, and then, the cells were washed three times with sterile PBS (1X) (Gibco, USA). Then, V. cholerae at MOI 10, which lead to the best cell persistence, subjected to each well and incubated at 37°C for 2 h in a 5% CO2 atmosphere. Following this time, each well was washed with sterile PBS (1X) to remove nonadherent bacteria to the cells. Then, 200 μL of 0.1% Triton X100 solution (Sigma-Aldrich, Germany) was added to each well and incubated at 20°C for 15 min. Following cell disruption with 0.1% Triton X-100, the bacterial suspensions were serially diluted (1:1000, 1:10,000, and 1:20,000). Next, 50 μL from each dilution was cultured on three BHI agar plates and then incubated for 24 h at 37°C. The plates were subsequently counted, and the average number of colonies was determined [12, 13].
2.5. Extraction of RNA and Synthesis of Complementary DNA (cDNA)
RNA extraction from Caco-2 cells was performed according to the protocol RNA Miniprep Super Kit (Bio Basic, Canada). For RNA extraction from Caco-2 cells, a 12-well microplate (SPL, Korea) was prepared. Briefly, after counting cells, 3 × 10^5^ cells were seeded into each well of culture plates containing 500 μL of DMEM enriched with 10% FBS and without antibiotics. The cells were incubated with 5% CO2 at 37°C for 24 h. Following this time, when the cells reached 80%–90% confluency, the medium was aspirated, and the cells were washed three times with sterile PBS (1X). Then, the cells were separately treated with both classical and El Tor biotypes of V. cholerae at MOI 10 and incubated overnight at 37°C in an atmosphere containing 5% CO2. Next, the culture medium was removed from each well, and the cells were washed three times with sterile PBS (1X) to remove unattached bacteria from each well. Consequently, the cells and attached bacteria from each well were harvested using RLT solution and transferred into a sterile microtube for RNA extraction [14]. Finally, RNA extraction was performed from treated and untreated (as negative control) Caco-2 cells. For RNA extraction of V. cholerae in BHI broth culture medium after calculating the attachment percentage based on the bacteria attached to the cells, an equivalent number of bacteria attached to Caco-2 cells was inoculated into the BHI broth medium and incubated overnight at 37°C. After 24 h, RNA was extracted from BHI broth medium separately from both classical and El Tor biotypes of V. cholerae. Optical absorbance at 260–280 nm was assayed to determine the concentration and purity of the extracted RNA. RNA integrity was investigated using electrophoresis on an agarose gel (1%), which allowed the observation of medium intensity bands without traces of degradation (Figure 1). Finally, cDNA was synthesized using RNA with a concentration of 1.8-2, according to the manufacturer's protocol of Yekta Tajhiz Azma from Iran [15].
2.6. Evaluation of the Expression of Virulence Factors of V. cholerae After Interaction With the Caco-2 Cell Line Using the Real-Time PCR Method
In the current study, the expression of ctxAB, tcpA, and hlyA genes of V. cholerae was evaluated under two conditions of Caco-2 cell lines and BHI broth environments using the real-time PCR method (ABI System, Applied Biosystems, USA). Briefly, the final reaction volume was 20 μL, containing 1 μL of reverse primer, 1 μL of forward primer, 6 μL of RNase-free water, 2 μL of cDNA, and 10 μL of SYBR Green master mix (Amplicon, Denmark). The primer sequences are detailed in Table 1. The 16s rRNA gene was used as an internal reference in this study. To determine the average ∆CT of each target gene, the CT value of the target genes was subtracted from the average CT of the internal control gene (16s rRNA). Next, the average ∆CT of the control group (classical and El Tor biotypes of V. cholerae in BHI broth medium) was subtracted from the average ∆CT of target genes to calculate the ∆∆CT value. Subsequently, relative quantification was carried out using standard 2^−ΔΔCT^ analysis. Real-time PCR experiments were carried out in triplicate [15]. The primer sequences are listed in Table 1.
2.7. Statistical Analysis
GraphPad Prism version 8 was used for evaluating data using one-way ANOVA and the Bonferroni test. Significance was determined at a p value < 0.05. The results are shown as mean ± SD from triplicate experiments.
3. Results
3.1. Persistence of Caco-2 Cells After Interaction With V. cholerae
In this study, five groups were investigated including untreated Caco-2 cells (as negative control) and treated cells with V. cholerae at MOI of 10, 20, 50, and 80. As shown in Figure 2, the viability of Caco-2 cells following interaction with V. cholerae at MOI of 10, 20, 50, and 80 were 50%, 60%, 91.7%, and 96.6%, respectively. The results indicated that the highest number of live cells was detected after exposure to MOI = 10. So, MOI of 10 as optimal MOI was used.
3.2. Evaluation of Attachment of V. cholerae to Caco-2 Cells
Because V. cholerae does not penetrate inside the cell, we assessed the number of bacteria attached to Caco-2 cells. The equivalent number of V. cholerae attached to Caco-2 cells was inoculated into BHI broth medium to maintain constant conditions, especially in terms of the bacterial count in both environments. The highest stability of cells was detected at MOI 10, and the attachment test was performed at this MOI. The counts from dilutions (10^−3^ 10^−4^, and 2 × 10^−4^) were 26 × 10^4^ for the classical biotype of V. cholerae and 22.6 × 10^4^ for the El Tor biotype of V. cholerae. Therefore, the optimal count from dilution 10^−4^ was then selected for BHI broth inoculation. As shown in Table 2, the percentages of bacteria attached at MOI of 10 in a dilution 10^−4^ to Caco-2 cells in the classical and El Tor biotypes of V. cholerae were 53% and 44%, respectively (Figures 3(a) and 3(b)).
3.3. Examination of the Expression of ctxAB, tcpA, and hlyA V. cholerae Genes in Caco-2 Cells and BHI Broth Medium
In the current study, the expression of V. cholerae genes including ctxAB, tcpA, and hlyA after interaction of Caco-2 cells with both El Tor and classical biotypes of V. cholerae strains was assayed than their expression in BHI broth medium (as a control). Our findings indicated that, after interaction of Caco-2 cells with the El Tor biotype of V. cholerae, fold changes of ctxAB, tcpA, and hlyA genes were of 2-, 1.02-, and 7-fold, respectively, while fold changes of these genes in the classical biotype of V. cholerae were 6-, 2-, and 13-fold, respectively. As shown in Figures 4 and 5, there was a significant increase in the expression levels of ctxAB and hlyA genes in both El Tor and classical biotypes of V. cholerae after interaction with Caco-2 cells compared to those in BHI broth medium. As shown in Figure 6, after interaction of Caco-2 cells with the classical biotype of V. cholerae, a significant increase was detected in the expression of the gene tcpA, while no significant increase was detected in the expression of this gene in the El Tor biotype. Moreover, after interaction of Caco-2 cells with the classical biotype of V. cholerae, fold changes of the genes ctxAB, tcpA, and hlyA were higher compared to the El Tor biotype of V. cholerae. After interaction of Caco-2 cells with the El Tor biotype of V. cholerae, the expression of the gene tcpA was a minimal increase (1.02-fold) than ctxAB and hlyA genes, while the expression of the gene hlyA in the classical biotype V. cholerae was the highest increase (13-fold) compared to the genes ctxAB and tcpA (Figure 6). These findings indicated that Caco-2 cell is an enhancer of the expression of the genes ctxAB, tcpA, and hlyA in the classical biotype of V. cholerae, while it is an enhancer of the genes ctxAB and hlyA in the El Tor biotype of V. cholerae. The enhancing effect of Caco-2 cells was more significant on the expression of the genes ctxAB, tcpA, and hlyA in the classical biotype compared to the El Tor biotype of V. cholerae. However, the enhancing effect of Caco-2 cells on the expression of the genes ctxAB, tcpA, and hlyA in the classical biotype is higher compared to the El Tor biotype of V. cholerae.
4. Discussion
Cholera is a global health challenge, especially in developing nations such as Iran [18]. It is caused by V. cholerae and leads to severe diarrhea [19]. V. cholerae inhabits aquatic environments and, under unfavorable conditions, regulates its virulence factors to adapt with the environment [20]. Therefore, a comprehensive understanding from virulence factors and behaviors of V. cholerae in diverse environmental conditions will facilitate more effective management of this global challenge. Few studies have assessed the effect of Caco-2 cells on the expression of virulence factors in other bacteria. For example, interactions of Caco-2 cells with Clostridium perfringens significantly increased the expression of the Cpe gene, compared to LB broth medium. They found that Caco-2 cells secrete a substance that suppresses the expression of the Cpe gene [7]. Moreover, bacteria have different behaviors in unfavorable conditions, they can impact on the expression of virulence factors, and they can modify the pathogenic ability to adapt to the environment. Key virulence factors of V. cholerae are CT, toxin-coregulated pilus (tcpA), and hemolysin A (hlyA) [21]. This study was designed to evaluate the effects of both Caco-2 cells (similar to the intestinal environment) and BHI broth (a suitable laboratory medium) on the expression of the genes ctxAB, tcpA, and hlyA of V. cholerae. To conduct a valid comparison in this study, after counting the bacteria attached to the Caco-2 cells, this number was inoculated into the BHI broth medium. The main findings of this study following interaction of V. cholerae with Caco-2 cells include the following: i) there was a significant increase in the expression levels of ctxAB and hlyA genes in both El Tor and classical biotypes of V. cholerae compared to those in BHI broth medium; ii) the expression of the gene tcpA was increased in the classical biotype of V. cholerae, while no significant increase was detected in the expression of this gene in the El Tor biotype of V. cholerae; iii) higher fold changes of ctxAB, tcpA, and hlyA genes were detected in the classical biotype compared to the El Tor biotype of V. cholerae; iv) the expression of the gene hlyA in the classical biotype of V. cholerae was the highest increase (13-fold) compared to ctxAB and tcpA genes; and v) no significant increase in the expression of the tcpA gene in the El Tor biotype of V. cholerae was detected (1.02-fold) compared to ctxAB and hlyA genes.
The results demonstrated that after interaction of the Caco-2 cell with V. cholerae, fold changes of the genes ctxAB, tcpA, and hlyA in the El Tor biotype of V. cholerae were lower than fold change in the classical biotype of V. cholerae. Previous studies indicated that the hlyA gene, similar to ctxAB and tcpA genes of V. cholerae, is repressed by the quorum sensing (QS) regulated transcription factor including HapR. The process of repression occurs on two distinct levels: transcriptional, independent of HapR, and posttranslational, mediated by HapR [22]. Briefly, the hly A gene in the El Tor biotype of V. cholerae is controlled by the QS system that associated with HapR, FUR, and hlyA proteins. HapR binds to the promoters of hlyA, hlyU, and FUR for suppressing of the gene hly A [22]. On the other hand, ctxAB and tcpA genes are controlled by the ToxR system, while the El Tor biotype does not express ToxR regulon in the Caco-2 cell environment. Therefore, ctxAB and tcpA genes are strictly regulated by both HapR and ToxR systems, while hlyA is regulated exclusively by the HapR system [23]. It suggested that after interaction of V. cholerae with Caco-2 cells, the secretory factors produced by the cells led to more suppress of the QS system in V. cholerae, so after interaction with Caco-2 cells, fold change of the gene hlyA in both biotypes is significantly higher than that of the genes ctxAB and tcpA. However, the fold change of the hlyA gene is lower in the El Tor biotype compared to the classical biotype of V. cholerae. The QS system, mediated by HapR, appears to more strongly repress the expression of the gene hlyA in the El Tor biotype compared to the classical biotype of V. cholerae in the Caco-2 cell environment [23]. Therefore, it is possible that the control of the expression of virulence factors in the El Tor biotype of V. cholerae be highly complex, potentially involving factors beyond the Tox R regulator. Despite the widely accepted notion that gene expression and protein expression are analogous, a direct correlation between the two is not always evident. In fact, while quantitative assessments of mRNA levels for a specific gene can offer insight into the amount of protein, this correlation is not always observed. A plethora of regulatory mechanisms exist between the process of RNA polymerase traversing the gene and the formation of mRNA and the subsequent protein output from the ribosome. The objective of this study was to determine the expression of ctxAB, tcpA, and hlyA genes in V. cholerae (El Tor and classical biotypes) after interaction with Caco-2 cells compared to BHI broth. Consequently, it can be deduced that the interpretation of results derived from gene expression analysis should be approached with caution.
However, further research is required in this field, and it is recommended to examine other virulence factors of V. cholerae in both Caco-2 cells and BHI broth medium.
5. Conclusion
In conclusion, this research suggests that changes in environmental conditions can influence the pathogenicity of V. cholerae which ultimately leads to the organism's adaptation to the environmental conditions. Considering different effects of Caco-2 cells on the expression of ctxAB, tcpA, and hlyA genes in the El Tor and classical biotypes of V. cholerae, it is possible for the involvement of another regulatory system, potentially beyond HapR and ToxR, in modulating these genes. Therefore, gaining more understanding of how V. cholerae adapts to adverse conditions by modulating virulence genes can provide novel insight for disease management and prevention strategies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Legros D. Global Cholera Epidemiology: Opportunities to Reduce the Burden of Cholera by 2030 Journal of Infectious Diseases 2018218 suppl_3S 137S 1403018410210.1093/infdis/jiy 486PMC 6207143 · doi ↗ · pubmed ↗
- 2Das B. Verma J. Kumar P. Ghosh A. Ramamurthy T. Antibiotic Resistance in Vibrio cholerae: Understanding the Ecology of Resistance Genes and Mechanisms Vaccine 202038 A 83A 923127287010.1016/j.vaccine.2019.06.031 · doi ↗ · pubmed ↗
- 3Hsiao A. Zhu J. Pathogenicity and Virulence Regulation of Vibrio cholerae at the Interface of Host-Gut Microbiome Interactions Virulence 2020111158215993317231410.1080/21505594.2020.1845039 PMC 7671094 · doi ↗ · pubmed ↗
- 4Montero D. A. Vidal R. M. Velasco J. Vibrio cholerae, Classification, Pathogenesis, Immune Response, and Trends in Vaccine Development Frontiers of Medicine 202310 p. 115575110.3389/fmed.2023.1155751 PMC 1019618737215733 · doi ↗ · pubmed ↗
- 5Tsou A. Frey E. Hsiao A. Liu Z. Zhu J. Coordinated Regulation of Virulence by Quorum Sensing and Motility Pathways during the Initial Stages of Vibrio cholerae Infection Communicative and Integrative Biology 20081142441970478710.4161/cib.1.1.6662 PMC 2633796 · doi ↗ · pubmed ↗
- 6Sun H. Chow E. C. Liu S. Du Y. Pang K. S. The Caco-2 Cell Monolayer: Usefulness and Limitations Expert Opinion on Drug Metabolism and Toxicology 2008443954111843334410.1517/17425255.4.4.395 · doi ↗ · pubmed ↗
- 7Wang C. Defoirdt T. Rajkovic A. The Effect of Caco-2 Cells on Sporulation and Enterotoxin Expression by Foodborne Clostridium perfringens Pathogens 2024136 p. 43310.3390/pathogens 13060433 PMC 1120655038921731 · doi ↗ · pubmed ↗
- 8Bhattaram V. Upadhyay A. Yin H.-B. Mooyottu S. Venkitanarayanan K. Effect of Dietary Minerals on Virulence Attributes of Vibrio cholerae Frontiers in Microbiology 20178 p. 9112857998310.3389/fmicb.2017.00911 PMC 5437166 · doi ↗ · pubmed ↗