Monogeneans on exotic Indian freshwater fish. 8. Co-translocation of Cichlidogyrus tilapiae (Monogenea, Dactylogyridae) with pindani Chindongo socolofi (Cichliformes, Cichlidae): first report of this parasite genus in India within aquarium trade facilities
Amit Tripathi, Chawan Matey, Antoine Pariselle, Maarten P. M. Vanhove

TL;DR
This study reports the first occurrence of the parasite Cichlidogyrus tilapiae in India, found on ornamental cichlid fish in aquarium trade facilities.
Contribution
This is the first report of the parasite genus Cichlidogyrus in India and adds a new host and location to the known distribution of C. tilapiae.
Findings
Cichlidogyrus tilapiae was detected in 22.7% of examined pindani fish in Indian aquarium shops.
Morphological and genetic analyses confirmed the presence of C. tilapiae in India.
The study expands the known host and geographic range of C. tilapiae.
Abstract
The pindani, Chindongo socolofi (Cichliformes, Cichlidae) is a popular freshwater ornamental fish from Lake Malawi in Africa. Although identifying parasites associated with the global ornamental fish trade is critical for developing biosecurity practices, little is known about the parasite fauna of C. socolofi. Therefore, this study sought to determine what monogenean parasites C. socolofi harbours in India. Adult specimens of this host species were collected from various aquarium shops across the country between 2020 and 2022, and their gills were subjected to parasitological examination. Monogeneans were detected in five host specimens (22.7%) with low mean intensities (6.2 ± 3.8). They were identified as Cichlidogyrus tilapiae (Monogenea: Dactylogyridae) based on the presence of the following morphometric characteristics: two pairs of anchors, two auricles on the dorsal bar, a…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1 Figure 1
Figure 1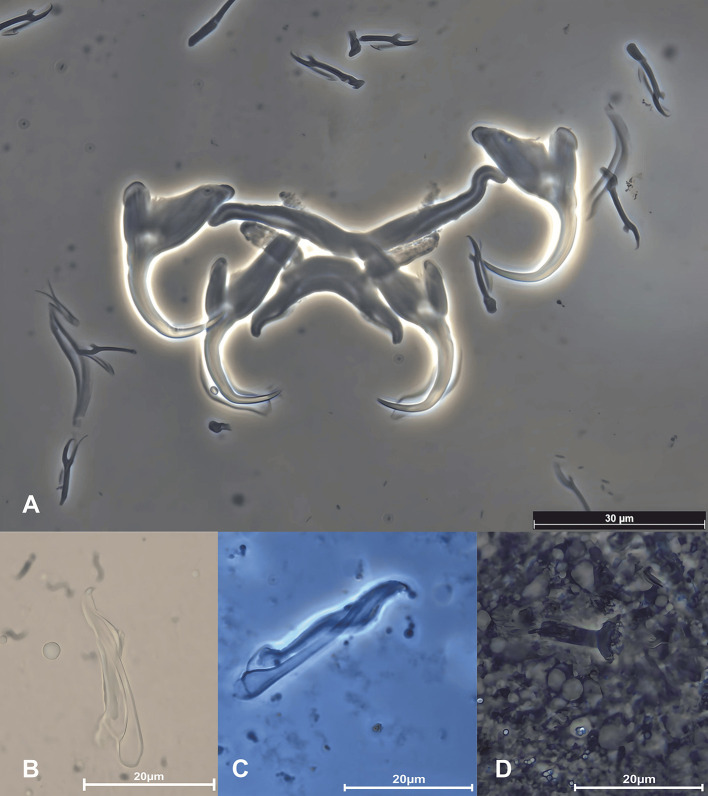 Figure 2
Figure 2 Figure 2
Figure 2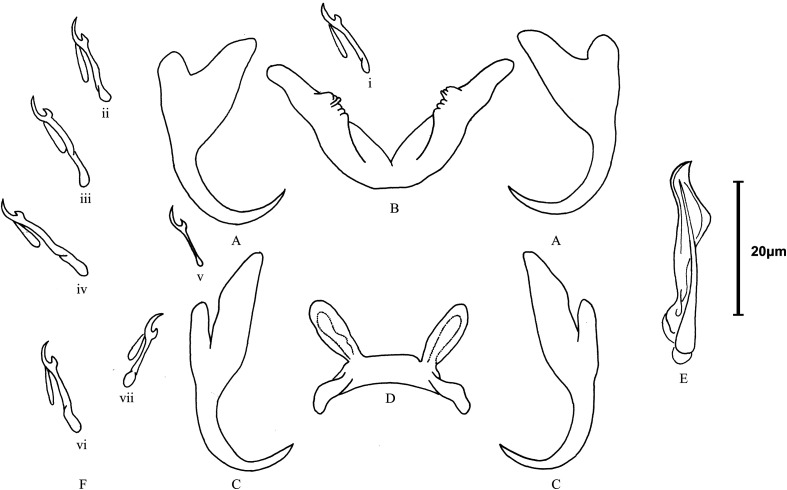 Figure 3
Figure 3 Figure 3
Figure 3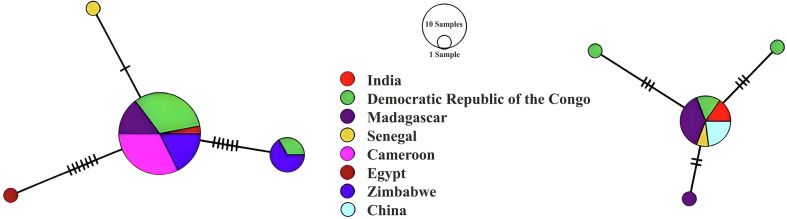 Figure 4
Figure 4 Figure 4
Figure 4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasite Biology and Host Interactions · Fish Biology and Ecology Studies · Myxozoan Parasites in Aquatic Species
Introduction
Cichlidogyrus Paperna, 1960 (Monogenea: Dactylogyridae) is the most species-rich African freshwater monogenean genus [37, 64]. It has 141 valid species [93] naturally parasitising primarily African cichlids (Cichlidae) and a few representatives of Cyprinodontidae Wagner, 1828 (Cyprinodontiformes) and Nandidae Bleeker, 1852 (Anabantiformes) [12, 20]. Some of these species have been identified as potentially pathogenic to fish, especially in aquaculture stocks [31, 56, 69]. Cichlidogyrus species, with few exceptions, are quite host-specific ([64], but also see [40]). An exception is Cichlidogyrus tilapiae Paperna, 1960. Since its first description from the Nile Tilapia, Oreochromis niloticus (Linnaeus 1758) (Cichliformes: Cichlidae) in Israel, C. tilapiae has been recorded in 31 different fish species in 27 countries across five continents, including Asia, Africa, North America, South America, and Australia (Table 1).
Table 1. Global distribution of Cichlidogyrus tilapiae Paperna, 1960 for 27 countries and 31 host fishes.Country/Host fishReference Australia Oreochromis mossambicus (Peters, 1852)[92] Bangladesh Oreochromis niloticus (Linnaeus, 1758)[17]Oreochromis mossambicus (Peters, 1852)[17] Brazil Coptodon rendalli (Boulenger, 1897)[13]Oreochromis niloticus (Linnaeus, 1758)[10, 91] Burkina Faso Oreochromis niloticus (Linnaeus, 1758)[9]Burundi Oreochromis niloticus (Linnaeus, 1758)[29] Cameroon Chromidotilapia guntheri (Sauvage, 1882)[51]Chromidotilapia linkei Staeck, 1980[51]Coptodon camerunensis (Lönnberg, 1903)[65]Coptodon gutturosa Stiassny, Schliewen and Dominey, 1992[65]Pelmatolapia mariae Boulenger, 1899[65]Tilapia kottae (Lönnberg, 1904) [now Coptodon kottae Lönnberg, 1904][65] China Coptodon zillii (Gervais, 1848)[94]Sarotherodon galilaeus (Linnaeus, 1758)[94]Oreochromis mossambicus (Peters, 1852)[94]Oreochromis niloticus (Linnaeus, 1758)[94] Colombia Oreochromis mossambica (Peters, 1852) [now Oreochromis mossambicus (Peters, 1852)][35] Democratic Republic of the Congo Coptodon tholloni (Sauvage, 1884)[27, 29]Oreochromis macrochir (Boulenger, 1912)[29]Oreochromis mortimeri (Trewavas, 1966)[28]Oreochromis mweruensis Trewavas, 1983[28–30, 32]Oreochromis niloticus (Linnaeus, 1758)[29, 30] Côte d’Ivoire Oreochromis niloticus (Linnaeus, 1758)[5] Cuba Oreochromis aureus (Steindachner, 1864)[49, 68] Egypt Oreochromis niloticus (Linnaeus, 1758)[4, 15] Coptodon zillii (Gervais, 1848)[15] Ghana Oreochromis niloticus (Linnaeus, 1758)[58, 59, 62]Sarotherodon galilaeus (Linnaeus, 1758)[58, 59, 62]Tilapia busumana (Günther, 1903)[59]Hemichromis fasciatus (Peters, 1857)[59]Chromidotilapia guntheri (Sauvage, 1882)[60, 62]Oreochromis aureus (Steindachner, 1864)[62] Israel Oreochromis niloticus (Linnaeus, 1758)[57]Sarotherodon galilaeus (Linnaeus, 1758)[57, 58]Tristramella sacra (Günther, 1865)[57]Tristramella simonis (Günther, 1864)[57] Japan Oreochromis mossambicus (Peters, 1852)[44]Oreochromis niloticus niloticus (Linnaeus, 1758)[44] Kenya Oreochromis niloticus (Linnaeus, 1758)[71]Oreochromis leucostictus (Trewavas, 1933)[71] Madagascar Oreochromis mossambicus (Peters, 1852)[77]Oreochromis niloticus (Linnaeus, 1758)[77]Coptodon rendalli (Boulenger, 1897)[77]Pachypanchax omalonotus (Duméril, 1861)[77]Ptychochromis oligacanthus (Bleeker, 1868)[77]Paratilapia polleni Bleeker, 1868[77]Paretroplus polyactis Bleeker, 1878[77] Malaysia Oreochromis niloticus (Linnaeus, 1758)[41]Red hybrid tilapia (Oreochromis mossambicus × O. niloticus)[41] Mexico Oreochromis aureus (Steindachner. 1864)[26]Oreochromis niloticus (Linnaeus, 1758)[26, 50, 63]Vieja fenestrata (Günther, 1860)[26] Nigeria Chromidotilapia guntheri (Sauvage, 1882)[51] Philippines Oreochromis niloticus (Linnaeus, 1758)[2, 52]Senegal Hemichromis fasciatus Peters, 1857[48]Oreochromis niloticus (Linnaeus, 1758)[48]Sarotherodon galilaeus (Linnaeus, 1758)[48]Coptodon guineensis Günther, 1862[48] South Africa Oreochromis mossambicus (Peters, 1852)[42, 43, 54]Pseudocrenilabrus philander (Weber, 1897)[54] Tanzania Oreochromis urolepis (Norman, 1922)[62] Thailand Oreochromis niloticus (Linnaeus, 1758)[45] O. niloticus x O. mossambicus [45] Uganda Oreochromis niloticus (Linnaeus, 1758)[62]Oreochromis spilurus (Günther, 1894)[62]Oreochromis leucostictus (Trewavas 1933)[61]Haplochromis macrognathus Regan, 1922[61]Oreochromis mossambicus (Peters, 1852)[61] Zimbabwe Oreochromis mortimeri (Trewavas, 1966)[14]
Pindani, Chindongo (Pseudotropheus) socolofi (Johnson, 1974) (Cichliformes, Cichlidae) is native to Lake Malawi in Africa [18, 46], and is available in two colour variants – normal (Blue pindani) and albino (White pindani) [33]. Despite the importance of C. socolofi in the ornamental fish market [74], little is known about its parasite fauna. To our knowledge, only one study on parasitic infections of C. socolofi exists [11]. These researchers studied the parasites of cichlids imported via the aquarium trade in Türkiye and recorded the protozoan parasite Trichodina pediculus Ehrenberg, 1831.
This study aimed to establish whether C. socolofi is infected by monogenean parasites and, if so, whether they were co-translocated into India via the ornamental fish trade. We demonstrate the presence of C. tilapiae in post-quarantine populations of C. socolofi collected from Indian aquarium markets. This was accomplished first by morphological characterisation (structure and measurements of the sclerotised parts of the haptor and reproductive organs) and subsequently by molecular characterisation (Sanger sequencing of 18S rRNA gene-ITS1 region and 28S rRNA genes). This paper is part of a series on exotic and/or invasive monogenean parasites imported into India via the ornamental trade [81–87].
Materials and methods
Ethics
This study was approved by the institutional ethics committee of the University of Lucknow under the protocol numbers LU/AEC/ZOO/2019 and 19/I/2024/IAEC/LU.
Sample collection and examination
Between January 2020 and December 2022, 22 specimens of C. socolofi (total weight: 3.12–6.50 g; and total length: 4.5–8.0 cm) (Fig. 1) were collected from aquarium shops in Lucknow, New Delhi, and Kolkata, India. Fish were shipped to the laboratory the same day after they were packaged in polybags containing water and pure oxygen. Individual fish were euthanised with an overdose of tricaine methanesulfonate (MS-222 @ 150 mg/L; Sigma Aldrich Co., St. Louis, MO, USA), followed by exsanguination by the removal of gill arches. Half of the gill arches were initially fixed in hot (60 °C) distilled water to relax and heat-kill the specimens before they were transferred to 4% formalin for microscopy following Kritsky [36]. The other half was preserved in 95% ethanol for genetic analysis. Some of the gill arches were examined fresh with live worms. Monogeneans were later isolated from these gills using fine dissecting needles under a stereomicroscope (Leica Microsystems, Wetzlar, Germany). Fish specimens were identified morphologically with the help of the ICAR-National Bureau of Fish Genetic Resources (ICAR-NBFGR), a premier Indian institute on fish taxonomy, biology, and genomics.
Figure 1. Freshly dead specimens of Chindongo socolofi (Johnson, 1974) examined for the present study. A. Blue pindani, B. White pindani. Photograph by Chawan Matey.
Morphological analysis
Formalin-fixed worms were stained with either Gomori’s trichrome or Borax carmine and mounted in DPX (dibutylphthalate polystyrene xylene) for observing internal anatomy (permanent mounts); others were mounted in glycerine jelly or Hoyer’s medium for the study of sclerotised parts of the haptor and reproductive organs (temporary mounts). Additionally, some ethanol-preserved worms were treated for 20–30 min at 55 °C with 1.0 μL of digestion buffer (0.1 μl of solid tissue buffer and 0.9 μL proteinase K) (Quick DNA^TM^ Miniprep Plus Kit, ZYMO Research, Irvine, CA, USA) to digest the tissues surrounding their sclerotised parts.
The morphology of the sclerotised parts was examined under a light microscope (Leica DM4B) at a magnification of 100×, using an oil immersion lens with phase-contrast (PHA-CO) and differential interference contrast illumination. Photographs and measurements (in micrometres) were obtained using a digital camera (Leica DFC7000 T) and imaging analysis software (LAS X; Leica Microsystems Ltd.) attached to the light microscope. A composite line drawing plate was made from multiple parasite specimens using an Olympus BX-51 microscope drawing tube. Species were identified based on the morphological characters described in previous studies [14, 15, 35, 44, 57, 64]. The terminology and measurement of these characters followed Rahmouni et al. [70]. The prevalence and mean intensity of infection were calculated according to Bush et al. [8].
DNA extraction and amplification
Representative samples of ethanol-preserved specimens were morphologically identified as conspecific to the temporary and permanent mounts before being pooled in two groups (n = 3) by two collection sites (Lucknow and New Delhi) for gDNA isolation using a DNA extraction kit (Extracta DNA Prep for PCR-Tissue, Quantabio, Beverly, MA, USA), according to the manufacturer’s instructions. Partial fragments of 18S ribosomal RNA genes (18S) and internal transcribed spacer 1 (ITS1) clusters were amplified with the primers s1 [78] and ir8 [75]. Meanwhile, those of 28S ribosomal RNA genes were amplified with the primers c1 and d2 [23].
Polymerase chain reactions were performed in an automated thermal cycler (Himedia Laboratories, Thane, MH, India) with reaction mixtures (final volume 20 μL) containing 4 μL of distilled water, 10 μL of 2× PCR TaqMixture (Himedia Laboratories), 1 μL of 10 pmol/microliter of each primer, and 4 μL of DNA template. The amplification profile for the 18S rRNA gene-ITS1 region was as follows: initial denaturation at 95 °C for 3 min, then 35 cycles of denaturation at 95 °C for 30 s, annealing at 50 °C for 30 s, and extension at 72 °C for 1 min, with a final extension at 72 °C for 7 min. The amplification profile for the 28S ribosomal RNA gene followed Šimková et al. [76]. The size of the PCR products (2 μL) was analysed by electrophoresis in 1.2% agarose gel prepared in 1× TAE buffer, prestained with 0.1 μL/mL 10,000× Sybr Safe in dimethyl sulfoxide (Invitrogen, Waltham, MA, USA), at 90 V for 30 min, and visualised and documented on a Bio-Print gel documentation imaging system (Vilber Lourmat, Collégien, France).
Sequence analysis
The PCR products were purified (on 1.5% agarose using a QIAquick PCR Purification Kit; QIAGEN, Germantown, MA, USA) and Sanger sequenced (on an ABI 3730xL automated sequencer; Applied Biosystems, Foster City, CA, USA) with PCR primers by Eurofins Genomics (Bengaluru, KA, India). SnapGene version 5.3 (https://www.snapgene.com) was used to manually quality-trim the successfully sequenced amplicons. Consensus sequences (18S-ITS1, 942 bp; 28S, 660 bp and 848 bp) were generated using the BioEdit Program [22].
Sequences, together with all sequences from the same markers and species retrieved from NCBI GenBank (Tables 2 and 3) were aligned using ClustalW [24] implemented in MEGA v.7 [38]. To obtain equal lengths for sequence analysis, they were trimmed to 687 bp (18S-ITS1) and 631 bp (28S). A median-joining network [3] was inferred for each marker using PopART [39].
Table 2. Information on Cichlidogyrus tilapiae Paperna, 1960, including hosts, localities, and GenBank accession numbers of their 18S+ITS1 rRNA gene sequences (as retrieved from the NCBI database on December 04, 2024).IsolatesHost fishGeographic locationAccession No.ReferenceAT.CT-2021 Chindongo socolofi India, Asia MZ266637 This studyPPZIM199_1 Coptodon rendalli Zimbabwe, Southern Africa ON819336 [21]PPZIM200_1 Coptodon rendalli Zimbabwe, Southern Africa ON819337 [21]PPZIM200_2 Coptodon rendalli Zimbabwe, Southern Africa ON819338 [21]PPZIM200_3 Coptodon rendalli Zimbabwe, Southern Africa ON819339 [21]PPZIM200_4 Coptodon rendalli Zimbabwe, Southern Africa ON819340 [21]PPZIM056_1 Oreochromis niloticus Zimbabwe, Southern Africa ON819297 [21]PPZIM059_1 Oreochromis niloticus Zimbabwe, Southern Africa ON819298 [21]PPZIM105_1 Oreochromis niloticus Zimbabwe, Southern Africa ON819310 [21]PPZIM117_1 Oreochromis niloticus Zimbabwe, Southern Africa ON819312 [21]PPZIM174_1 Oreochromis cf. mortimeri Zimbabwe, Southern Africa ON819331 [21]PPKAT465_1 Oreochromis niloticus D. R. Congo, Central Africa ON819262 [21] PPKAT482_1 Oreochromis niloticus D. R. Congo, Central Africa ON819266 [21] PPKAT485_2 Oreochromis niloticus D. R. Congo, Central Africa ON819268 [21] PPKAT495_1 Oreochromis niloticus D. R. Congo, Central Africa ON819269 [21] PPKAT997_1 Oreochromis mweruensis D. R. Congo, Central Africa ON819289 [21] PPKAT1074_2 Coptodon rendalli D. R. Congo, Central Africa ON819246 [21]PPKAT1074_3 Coptodon rendalli D. R. Congo, Central Africa ON819247 [21] PPKAT1074_4 Coptodon rendalli D. R. Congo, Central Africa ON819248 [21] PPKAT1039_1 Oreochromis aureus D. R. Congo, Central Africa ON819235 [21] PPKAT1039_2 Oreochromis aureus D. R. Congo, Central Africa ON819236 [21] PPKAT1039_3 Oreochromis aureus D. R. Congo, Central Africa ON819237 [21] PPKAT1039_5 Oreochromis aureus D. R. Congo, Central Africa ON819238 [21] PPKAT1002_1 Oreochromis mweruensis D. R. Congo, Central Africa ON819220 [21] PPCAM347_2 Coptodon guineensis D. R. Congo, Central Africa ON819204 [21] PPCAM334_1 Coptodon guineensis D. R. Congo, Central Africa ON819198 [21] PPCAM339_1 Coptodon guineensis D. R. Congo, Central Africa ON819199 [21] PPCAM340_1 Coptodon guineensis D. R. Congo, Central Africa ON819200 [21] PPCAM340_2 Coptodon guineensis D. R. Congo, Central Africa ON819201 [21] PPCAM342_1 Coptodon guineensis D. R. Congo, Central Africa ON819202 [21] PPCAM347_1 Coptodon guineensis D. R. Congo, Central Africa ON819203 [21] PPCAM037_1 Oreochromis niloticus Cameroon, Central Africa ON819182 [21]PPCAM059_1 Oreochromis niloticus Cameroon, Central Africa ON819186 [21]PPCAM083_1 Oreochromis niloticus Cameroon, Central Africa ON819187 [21]PPCAM088_2 Oreochromis niloticus Cameroon, Central Africa ON819189 [21]RAKAN10–Egypt, North Africa OR793160 UnpublishedKMC43 Oreochromis mossambicus Madagascar, East Africa MH767396 [77]KMC44 Oreochromis niloticus Madagascar, East Africa MH767397 [77]KMC45 Pachypanchax omalonotus Madagascar, East Africa MH767398 [77]KMC46 Ptychochromis oligacanthus Madagascar, East Africa MH767399 [77]KMC47 Paratilapia polleni Madagascar, East Africa MH767400 [77]PC43 Hemichromis fasciatus Senegal, West Africa HE792797 [48]1 Sarotherodon galilaeus Côte d’Ivoire, West Africa AJ920276 [66]2 Sarotherodon galilaeus Côte d’Ivoire, West Africa AJ920277 [66]3 Sarotherodon galilaeus Côte d’Ivoire, West Africa AJ920275 [66]
Table 3. Information on Cichlidogyrus tilapiae Paperna, 1960, including hosts, localities, and GenBank accession numbers of their 28S rRNA gene sequences (as retrieved from the NCBI database on December 04, 2024).IsolatesHost fishGeographic locationAccession No.ReferenceAT-2021 Chindongo socolofi India, Asia MZ265190 This studyCT-28S-CS-NDLS-22 Chindongo socolofi India, Asia PQ675652 This studyB2 Oreochromis mossambicus China, Asia PP448734 Unpublishede2-7 Oreochromis niloticus China, Asia OR557581 UnpublishedE3-3 Oreochromis niloticus China, Asia OR488785 Unpublished208CtilOnilMonzi Oreochromis niloticus D. R. Congo, Central Africa OM720051 [30]217CtilOnilMonzi Oreochromis niloticus D. R. Congo, Central Africa OM720052 [30]238CtilOnilNdim Oreochromis niloticus D. R. Congo, Central Africa OM720053 [30]64CtilOmwKipo Oreochromis mweruensis D. R. Congo, Central Africa OM720054 [30]KMC59 Paratilapia polleni Madagascar, East Africa MH767412 [77]KMC58 Ptychochromis oligacanthus Madagascar, East Africa MH767411 [77]KMC57 Pachypanchax omalonotus Madagascar, East Africa MH767410 [77]KMC56 Oreochromis niloticus Madagascar, East Africa MH767409 [77]KMC55 Oreochromis mossambicus Madagascar, East Africa MH767408 [77]KMC54 Coptodon rendalli Madagascar, East Africa MH767407 [77]– Hemichromis fasciatus Senegal, West Africa HQ010029 [47]
As additional assessment for species-level identification, intraspecific genetic differences (also known as genetic distances) among different geographical isolates of C. tilapiae were computed from the same dataset. This was done using the Kimura two-parameter (K2P) model [34] of nucleotide substitution in MEGA 11 [79], with gaps treated as complete deletions.
Results
Thirty individuals of a single monogenean species – namely Cichlidogyrus tilapiae Paperna, 1960 – were collected from the gills of five specimens of C. socolofi, with a low prevalence (22.72%) and low infection intensities (7–13 worms/fish). Brief morphological and molecular data of this species are presented below.
Taxonomic summary
Phylum Platyhelminthes Minot, 1876
Superclass Neodermata Ehlers, 1985
Class Monogenea van Beneden, 1858
Family Dactylogyridae Yamaguti, 1963
Genus Cichlidogyrus Paperna, 1960
Cichlidogyrus tilapiae Paperna, 1960 (Figs. 2 and 3)
Type host and locality: Nile tilapia Oreochromis niloticus (Linnaeus, 1758) (Cichliformes: Cichlidae); Israel (Paperna 1960).
Figure 2. Photomontage of light microscopy and phase contrast (PHACO) images of Cichlidogyrus tilapiae Paperna, 1960 from Chindongo socolofi (Johnson, 1974). A. anchor-bar complex and hooks, B and C. different configurations of male copulatory organ, D. vagina. Photograph by Amit Tripathi.
Figure 3. Line drawings of sclerotised structures of Cichlidogyrus tilapiae Paperna, 1960 from Chindongo socolofi (Johnson, 1974). A. ventral anchor, B. ventral bar, C. dorsal anchor, D. dorsal bar, E. male copulatory organ, F. hook (pairs i–vii). Scale bar = 20 μm. Figure by Amit Tripathi.
Present host: Chindongo socolofi (Cichliformes, Cichlidae).
Present material and collection date: Aquarium shops in Lucknow (26.8467° N, 80.9462° E), New Delhi (28.6139° N, 77.2090° E), and Kolkata (22.5726° N, 88.3639° E), India; January 2020–December 2022.
Site of infection: Gills.
Infection parameters: Prevalence: 22.72% (5 out of 22 C. socolofi examined); Mean infection intensity: 6.2 ± 3.86 (7–13; n = 5).
Museum material: Five voucher specimens stained with Gomori’s trichrome or Borax carmine and mounted on glass slides in DPX (Smithsonian Institution, USA; USNM 1757684-1757688).
GenBank deposition: 18S-ITS1: 942 bp (MZ266637); 28S: 660 bp (MZ265190), 848 bp (PQ675652).
Morphological data
Cichlidogyrus is distinguished by a vas deferens that does not encircle the intestinal caecum, two pairs of anchors (one dorsal and one ventral), two transversal bars (a dorsal bar with two typical auricles and a V-shaped ventral bar), seven pairs of hooks, a sclerotised or non-sclerotised vagina and a sclerotised male copulatory organ comprising a male copulatory tube and (often but not always) an accessory piece [57, 88].
Our specimens presented nearly identical morphological features of sclerotised parts (both haptoral and reproductive ones) indicated in the original description [55] and subsequent redescriptions or illustrated records [14, 15, 35, 44] of C. tilapiae (Figs. 2 and 3) (Table 4). Only two minor discrepancies were observed in the morphometry of the haptoral armaments. First, our specimens had a slightly longer and deeper outer root of the dorsal anchor (4**–**6 μm), compared to their conspecifics. Second, variations were observed in the ranges of measurements of the ventral bar. Paperna [57], for example, measured the length of the ventral bar to be 34–98 μm, whereas Douëllou [14] measured it to be 26–33 μm (as we did), and Kritsky and Thatcher [35], Ergens [15], and Maneepitaksanti and Nagasawa [44] measured it to be 50–65 μm.
Table 4. Comparative measurements (in μm) of reproductive organs and haptoral armaments of Cichlidogyrus tilapiae Paperna, 1960 from India (present study) and other geographical locations.MeasurementPaperna [57]Kritsky & Thatcher [35]Ergens [15]Douëllou [14]Maneepitaksanti & Nagasawa [44]Present studyCountryIsraelColombiaEgyptZimbabweJapanIndiaReproductive organ Copulatory tube19**–37 2930–3332 (30–36)29 (28–30)32 (25–35) Accessory piece22–3331–33 (31–33)33 (31–37)31 (30–32)Haptoral partsDorsal anchor length26–40 4037–4142 (41–44)36 (35–38)37 (32–42)Dorsal bar length18–38 2827–2929 (28–30)14 (13–15)28 (22–34)Dorsal bar width–––––(4–5)Dorsal anchor inner root11–15–10–1318 (16–19)–16 (14–18)Dorsal anchor outer root4–7–3–64 (3–5)–5 (4–7)Ventral anchor length26–33 3129–3334 (32–36)30 (28–32)33 (31–36)Ventral bar length34–985756–6532 (31–33)54 (50–56)29 (26–32)Ventral bar width–––––5 (4–6)Ventral anchor inner root18–14–1812 (10–14)–15 (13–18)Ventral anchor outer root4–7–3–54 (3–5)–6 (5–7)Auricle length9–19–14–18––12 (11–13)Hook length–15–––11–19Pair I12 (7–17)–18 (17–19)14 (13–14)13 (12–15)(14–15)Pair II15 (13–17)–11(10–11)11 (9–12)10 (8–12)14 (13–15)Pair III16 (13–20)–18 (17–19)15 (13–17)14 (12–17)17 (16–18)Pair IV16 (13–20)–18 (17–19)17 (16–17)14 (12–15)(18–19)Pair V13 (11–15)–18 (17–19)18 (16–19)16 (15–17)(11–12)Pair VI15–18 (17–19)17 (17–18)16 (15–18)(17–18)Pair VII21–18 (17–19)15 (14–16)14 (13–**15)15– shows that these measurement values were not provided by the respective authors.
We were also able to locate the vagina in a single live specimen, which had gone unnoticed in previous studies on C. tilapiae. It resembled a short unsclerotised (muscular?) tube with a funnel-like opening at one end (Fig. 3D). We lost it quickly however, when the vitellaria burst out of the parasite body, killing it. Therefore, our identification of the vagina may not be conclusive and should be reconfirmed. Cichlidogyrus tilapiae has previously been adequately described/redescribed and, thus, does not need to be formally redescribed here.
Molecular data
The partial 18S rRNA gene-ITS1 region (942 bp) and 28S rRNA genes (660 bp and 848 bp) were sequenced from two pools of C. tilapiae specimens collected from C. socolofi in aquarium shops in India. Comparative analysis of these sequences against the NCBI GenBank non-redundant database using the “megablast” algorithm (https://blast.ncbi.nlm.nih.gov/Blast.cgi) revealed “near perfect” matches for 28S rRNA (659/660 bp; 99.85% similar identity with a query coverage of 100%) and 18S rRNA-ITS1 (935/938 bp; 99.68% similar identity with a query coverage of 99%) to C. tilapiae from Paratilapia polleni Bleeker, 1868 (Cichliformes, Cichlidae) in Madagascar deposited in GenBank under the accession numbers MH767412 (28S) and MH767400 (18S-ITS1), respectively [77]. These findings suggest their conspecificity (Tables 5 and 6).
Table 5. Intraspecific genetic distances (Kimura 2-parameter model with partial deletion option) and variations between our samples and conspecific references (most similar BLAST hits) of Cichlidogyrus tilapiae Paperna, 1960 based on 18S rRNA gene-ITS1 region.Sample reference (Host; Geographic location)Identity (%) E-valueGenetic distance Conspecific references (Host; Geographic location)18SITS1Cichlidogyrus tilapiae MZ266637 (Chindongo socolofi; India)99.680.000.0000.000Cichlidogyrus tilapiae MH767400(Paratilapia polleni; Madagascar, East Africa)99.780.000.0000.002Cichlidogyrus tilapiae HE792797(Hemichromis fasciatus; Senegal, West Africa)99.680.000.0000.000Cichlidogyrus tilapiae MH767399(Oreochromis niloticus; Madagascar, East Africa)
Table 6. Intraspecific genetic distances (Kimura 2-parameter model with partial deletion option) and variations between our samples and conspecific references (most similar BLAST hits) of Cichlidogyrus tilapiae Paperna, 1960 based on 28S rRNA gene.Sample reference (Host; Geographic location)Identity (%) E-valueGenetic distanceConspecific references (Host; Geographic location)Cichlidogyrus tilapiae MZ265190 (Chindongo socolofi; India)99.850.000.000Cichlidogyrus tilapiae MH767412(Paratilapia polleni; Madagascar, East Africa)99.850.000.000Cichlidogyrus tilapiae HQ010029(Hemichromis fasciatus; Senegal, West Africa)99.850.000.000Cichlidogyrus tilapiae MH767409(Oreochromis niloticus; Madagascar, East Africa)
The intraspecific genetic distances for 28S rRNA and 18S rRNA genes between four geographic isolates of C. tilapiae from different hosts and geographical locations were determined at 0%, indicating their conspecificity (Tables 5 and 6). The genetic distance for the ITS1 sequence, another marker with higher variability, was also determined between 0 and 0.002% (Table 5).
The haplotype network indicated that Indian haplotypes, for both markers, were widespread, and shared with conspecifics from both native and introduced populations (Fig. 4).
Figure 4. Median-joining haplotype networks based on a 687 bp fragment of small subunit rDNA and the first Internal Transcribed Spacer (left) and 631 bp of large subunit rDNA (right) from the newly sequenced individuals of Cichlidogyrus tilapiae from India, aligned with all previously published sequences from this species. Genotypes are represented by circles, with the size of the circle correlating with the number of isolates displaying the respective genotype. Colours denote the countries of sampling localities. Genotypes are connected by lines indicating the number of mutations between them.
Discussion
This paper is the first to document the presence of a member of Cichlidogyrus in India. In addition, C. socolofi has been identified as a new host species for C. tilapiae. This is also the first time a species of Cichlidogyrus is formally reported from a Malawi cichlid; members of the genus are known to occur on the lake’s cichlids but were mentioned without species-level identification [6]. Although there are a few variations in morphometrical data between different geographic isolates of C. tilapiae, we do not consider these differences to merit species-level separation. These variations may be attributable to differences in the host species [55], environmental factors [7], developmental stages [85, 90], individual variations within the species, or even the different fixation [16] or measuring methods employed thus far. For instance, just as we did, Douëllou [14] measured only one branch of the ventral bar following established norms of measurements for Cichlidogyrus [70]. Meanwhile, Kritsky and Thatcher [35] and Ergens [15] measured the total length in a “straight line extending between the two most distant parts”. Unfortunately, Paperna [57] and Maneepitaksanti and Nagasawa [44] did not specify their measurement methods.
While many publications have recorded only the occurrence of C. tilapiae without providing any morphometric data, those that have provided such data have shown a few variations in the sclerotised parts. For example, Paperna [57] found that both pairs of anchors were “about the same length”. However, all subsequent investigations have clearly shown that dorsal anchors are slightly larger than the ventral anchors. Paperna [57] also described and illustrated an accessory piece that terminated in a “bent bifurcated tip”, but this bifurcation has not been observed in any other studies. Furthermore, Ergens [15] and Douëllou [14] noted a small “groove on the base of dorsal anchor”, that no other researchers have reported. Ergens [15] also illustrated a small sliver-like structure on the outer roots of the ventral anchor, which has not been described or illustrated by other researchers.
Nonetheless, the distinctive morphology of the male copulatory organ, which lacks a heel and has a hook-shaped terminal end of the accessory piece, is consistent enough in all illustrations of C. tilapiae to be considered the most reliable diagnostic trait for identifying this species. This aligns with the notion that identification of Cichlidogyrus species is primarily based on the morphology of the reproductive hard parts [89].
Curiously, the 18S rRNA gene-ITS1 region and 28S rRNA gene sequences of C. tilapiae found in India differed from their conspecific references by only 3 bp and 1 bp, respectively (see above). Different phenotypes of C. tilapiae did not cluster monophyletically in the recent morphology-based phylogeny [51]. Therefore, we speculate that either C. tilapiae comprises a species complex of morphologically variable but closely related lineages [66] or that there are geographical variants of a single species.
The haplotype networks (Fig. 4) indicate that the haplotype of C. tilapiae found in India occurs widely throughout native and introduced host and parasite populations. Other markers than the ones used here, for example a sequence fragment of the cytochrome c oxidase subunit 1 gene, allow higher resolution distinction between populations of C. tilapiae [30] and may allow the identification of native and (co-)introduced strains of cichlid parasites [21].
Indian scenario
Since nothing is known about the monogenean fauna of C. socolofi in the wild, we cannot ascertain whether it is a natural host of C. tilapiae or whether it acquired it from other cichlids cohabiting in aquarium conditions. However, it is highly likely that C. socolofi is a regular host for C. tilapiae because the latter was consistently recovered over space (Lucknow, New Delhi, and Kolkata) and time (January 2020–December 2022). Chindongo socolofi is currently maintaining its self-sustaining populations in the country’s aquacultural ponds and has yet to be recorded in the wild. The potential negative impact of C. socolofi on India’s environment and/or economy will therefore depend on its ability to successfully establish, dominate, and expand in Indian waters. It has previously been hypothesised that the invasion success of a fish is linked to, amongst other things, favourable environmental conditions in the new habitat that are comparable to those in its native ranges (climate match theory; [1, 25]), and to the enemy release hypothesis [80]. Chindongo socolofi may be considered a potentially invasive fish species in this context because the climatic conditions in India, particularly in South India, are similar to those found in the native range of C. socolofi (southeastern Africa), including a tropical climate and a temperature range of 24–26 °C [18].
Should C. socolofi become invasive in Indian waters, the low host specificity of C. tilapiae (see above), combined with the native fish species’ lack of protective immunity against exotic parasites [72], could pose a serious biological invasion challenge. It is worth noting that C. tilapiae has already demonstrated its ability to switch from introduced cichlids to native hosts in destination environments, such as Vieja fenestrata (Günther, 1860) (Cichliformes, Cichlidae) (syn. = Paraneetroplus fenestratus) in Mexico [19], and Coptodon tholloni (Sauvage 1884) (Cichliformes, Cichlidae) in the Lower Congo Basin [29], and even non-cichlid hosts: Pachypanchax omalonotus (Duméril, 1861) (Cyprinodontiformes: Aplocheilidae) in Madagascar [77]. In fact, tilapia-infecting monogeneans have been proposed as the most ubiquitous tropical freshwater fish parasites globally, with C. tilapiae being one of the species most frequently reported as co-introduced with translocated tilapias [73]. However, to the best of our knowledge, this is the first report of this parasite from the ornamental fish trade.
The presence of C. tilapiae on C. socolofi highlights an additional challenge in India namely, illegal ornamental fish trafficking. The “Guidelines for import of ornamental fishes into India” [53] includes an “indicative list” of 92 exotic ornamental fish species that the Government of India has approved for import. Although C. socolofi is not on this list, it is widely available in Indian domestic trade ([67], this report).
Clearly, the fish were acquired illegally via international smuggling. Unfortunately, the “Guidelines” makes no clear or implicit declaration prohibiting the import of ornamental fish that are not on the “indicative list”, nor does it suggest that violators will face prosecution or even a fine. It simply states that “the import permit shall be cancelled forthwith and all the specimens imported destroyed without any notice to or permission of the importer”.
Given that the issue at hand involves not only fish trafficking but also the trafficking of accompanying (unidentified and often overlooked) parasites, merely cancelling import permits is a minor step toward protecting biodiversity and deterring traffickers. In reality, illegally imported exotic species are more likely to introduce parasites and diseases into the country because they bypass the import risk analysis and quarantine procedures of the importing country. Therefore, we recommend that the sale of a non-permitted ornamental fish species be treated as a criminal offence comparable to wildlife smuggling and implementing heavy penalties for this crime.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Agostinho AA, Suzuki HI, Fugi R, Alves DC, Tonella LH, Espindola LA. 2015. Ecological and life history traits of Hemiodus orthonops in the invasion process: looking for clues at home. Hydrobiologia, 746, 415–430.
- 2Arthur JR, Lumanlan-Mayo S. 1997. Checklist of the parasites of fishes of the Philippines. Rome: FAO.
- 3Bandelt H, Forster P, Röhl A. 1999. Median-joining networks for inferring intraspecific phylogenies. Molecular Biology and Evolution, 16, 37–48.10331250 10.1093/oxfordjournals.molbev.a 026036 · doi ↗ · pubmed ↗
- 4Bayoumy EM, El-Monem SA. 2012. Functional adaptation of branchial and stomach dactylogyrid monogenean: Cichlidogyrus and Enterogyrus isolated from Oreochromis niloticus, Proceedings of the 5th Global Fisheries & Aquaculture Research Conference, Egypt, 1 – 3 October, p. 353–360.
- 5Blahoua GK, Yao SS, Etile RND, N’Douba V. 2016. Distribution of gill Monogenean parasites from Oreochromis niloticus (Linnaeus, 1758) in man-made Lake Ayame I, Côte d’Ivoire. African Journal of Agricultural Research, 11, 117–129.
- 6Blais J, Rico C, van Oosterhout C, Cable J, Turner GF, Bernatchez L. 2007. MHC adaptive divergence between closely related and sympatric African cichlids. P Lo S One, 2, e 734.17710134 10.1371/journal.pone.0000734 PMC 1939875 · doi ↗ · pubmed ↗
- 7Brazenor AK, Saunders RJ, Miller TL, Hutson KS. 2018. Morphological variation in the cosmopolitan fish parasite Neobenedenia girellae (Capsalidae: Monogenea). International Journal of Parasitology, 48, 125–134.29122607 10.1016/j.ijpara.2017.07.009 · doi ↗ · pubmed ↗
- 8Bush AO, Lafferty KD, Lotz JM, Shostak AW. 1997. Parasitology meets ecology on its own terms: Margolis et al. revisited. Journal of Parasitology, 83, 575–583.9267395 · pubmed ↗