Incriminating vectors of deer malaria (Plasmodium odocoilei) at a Florida deer farm
Morgan Rockwell, Samantha M. Wisely, Derrick K. Mathias, Nathan D. Burkett-Cadena

TL;DR
This study identifies potential mosquito vectors of deer malaria in Florida by analyzing mosquito feeding habits and infection rates.
Contribution
The study incriminates specific mosquito species as likely vectors of Plasmodium odocoilei based on field data and infection rates.
Findings
Anopheles quadrimaculatus s.l., An. punctipennis, and An. crucians s.l. specialized in feeding on white-tailed deer and were infected with P. odocoilei.
An. quadrimaculatus s.l. and An. punctipennis had the highest deer blood meal percentages and infection rates, making them likely natural vectors.
Further laboratory studies are needed to confirm the biological transmission capability of these mosquitoes.
Abstract
Plasmodium odocoilei, the only nonhuman Plasmodium parasite of native mammals in North America, infects white-tailed deer (WTD) throughout the eastern USA. Although deer malaria is not a significant cause of disease in healthy deer, infection with P. odocoilei may increase susceptibility to infection with and mortality due to epizootic hemorrhagic disease virus in deer fawns. The incrimination of the vector(s) of deer malaria is an essential step in developing management plans for reducing the incidence of deer malaria. At a deer farm in Gadsden County, FL, with previously documented evidence of deer malaria transmission, mosquitoes were collected using carbon-dioxide-baited light traps, aspirators, and resting shelters. White-tailed deer host use and P. odocoilei infection rates were quantified in potential vector mosquito samples using polymerase chain reaction and Sanger sequencing.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
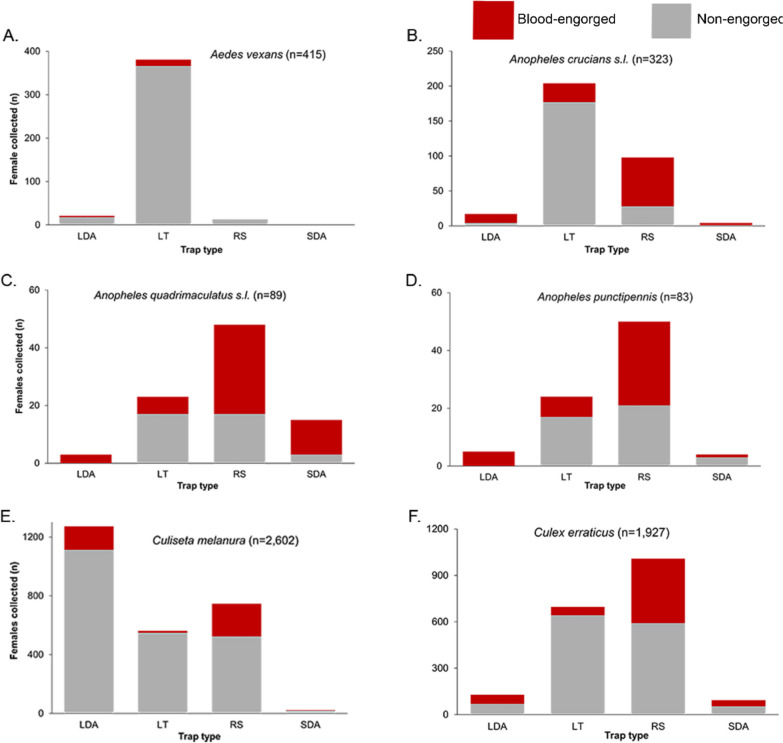 Figure 1
Figure 1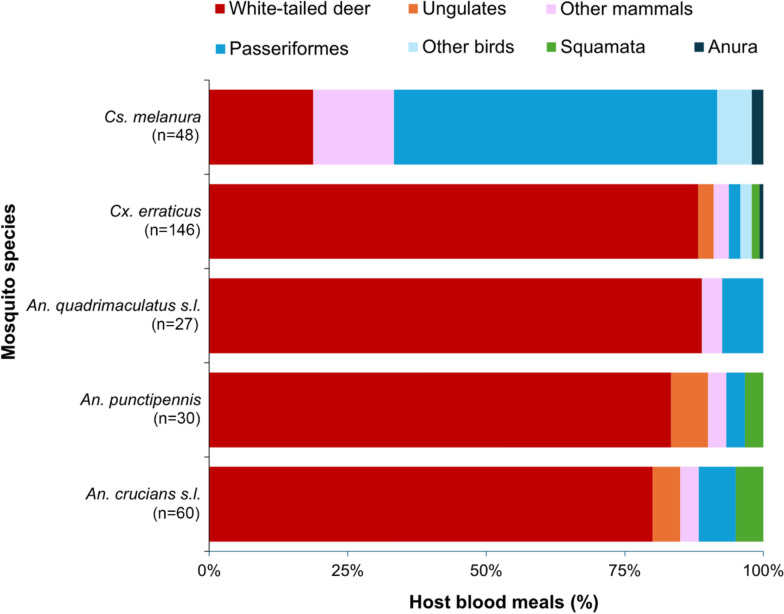 Figure 2
Figure 2 Figure 3
Figure 3- —Florida Legislature
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBird parasitology and diseases · Viral Infections and Vectors · Vector-borne infectious diseases
Background
Deer farming is an important industry in the USA, with annual revenue reaching up to USD 7.9 billion [1]. However, deer farming has been heavily impacted by various vector-borne diseases [2–4]. Deer malaria is the only malaria affecting nonhuman mammals in North America [5, 6]. It is caused by Plasmodium odocoilei, member of a genus of protozoans that infects the red blood cells of their vertebrate hosts and is transmitted primarily by mosquitoes [7]. A survey of several ungulate species across the USA found that P. odocoilei only infects Odocoileus virginianus, white-tailed deer (WTD), which is the most widely distributed wild ungulate in North America [8].
The impacts of P. odocoilei on WTD health are poorly understood, but the parasite likely causes subclinical disease [2] and may also increase susceptibility to infection with arboviruses and arbovirus-induced mortality [2, 8]. Plasmodium odocoilei was discovered and then described from the blood of a splenectomized WTD in Texas, USA, during a survey of hemosporidian parasites in deer in 1967 [5, 6]. Some general signs of disease were noted, although they were not definitively attributable to deer malaria [2]. Deer malaria was not reported again until it was detected by researchers investigating transmission ecology of avian malaria in 2016 [9]. During that study, an unidentified malaria parasite was detected in a blood-engorged female of Anopheles punctipennis, a species of mosquito that rarely bites birds [10–14]. Polymerase chain reaction (PCR) and sequencing determined that the An. punctipennis female fed upon a WTD, leading them to suspect and then confirm that the parasite was P. odocoilei [9]. By screening diverse ungulates across the USA, the investigators found a high infection rate of Plasmodium in WTD across the eastern USA (25%, 77/308), but not in the western USA or other ungulate species [9]. Although the direct effects of P. odocoilei on WTD health are rarely reported, a study conducted on a deer farm in northern Florida demonstrated that a substantial percentage of fawns acquired P. odocoilei during their first 8 months of life (21%, 7/33) and documented a potential association between P. odocoilei infection and decreased fawn survival [8]. The study also found a higher infection rate of epizootic hemorrhagic disease virus (EHDV) in fawns infected with Plasmodium (86%, 6/7) compared with fawns that were not infected with P. odocoilei (50%, 13/26). These findings suggested that infection with P. odocoilei may increase the risk of acquiring EHDV and potentially other arboviruses [8].
The vector of P. odocoilei must be incriminated to understand the transmission of the parasite and to develop integrated pest management (IPM) strategies focused on the vector species. The following criteria must be fulfilled before a putative vector species can be incriminated as the vector(s) of P. odocoilei with reasonable certainty: (1) a spatial and temporal relationship between putative vector species and the host must correlate, (2) the vector must be in adequate numbers to transmit the parasite, (3) the putative vector species must be associated with feeding on the host carrying the pathogen, (4) the vector species must be naturally infected with the parasite in the wild, and (5) the putative vector species must be capable of experimentally transmitting the parasite between hosts in laboratory settings [15–18].
In this study, our goal was to partially incriminate vectors of deer malaria, focusing on the presence, host use, and P. odocoilei infection in mosquito species at a deer farm with documented transmission of deer malaria in northern Florida [8]. We determined which mosquito species specialize in feeding on WTD using PCR-based blood meal analysis and Sanger sequencing [19, 20]. We also compared the P. odocoilei infection rates among mosquito species using a nested PCR-assay. These data are essential to incriminate the vector(s) of deer malaria.
Methods
Sampling sites
Mosquitoes were sampled from a northern Florida deer farm in Gadsden County, which is used for breeding and hunting WTD and other cervid and bovine game species [21–23]. The 200 ha property included ten high-fenced pens with improved pastures (8.5 ha), which housed approximately 100 WTD, at a density of approximately 12 WTD/ha. Sick and injured WTD were housed in the “barn pens,” while healthy WTD used for breeding were kept in the “breeding pens.” Free-ranging WTD and other ruminants were located within the preserve, which encompassed the surrounding Gulf Coast Forest and contained around 130–150 WTD (0.74/ha). The landscape of the property consisted of pastures, wetlands, and mixed deciduous/coniferous forests [21, 23, 24]. The farm is fully characterized, including maps, in prior studies [21, 23].
Mosquito sampling, identification, and processing
A variety of sampling methods were employed, including pop-up resting shelters, a large-diameter aspirator, a small-diameter aspirator, and carbon-dioxide-baited Centers for Disease Control and Prevention (CDC) miniature light traps (Model 2836BQ, BioQuip, Rancho Dominguez, CA, USA) to collect a diverse assemblage of mosquito species of different physiological statuses, including blood-engorged and nonengorged females [25]. Mosquitoes were collected daily during each of eight collection trips that lasted for 5 days each (October 2022; May, July, August, September, and October 2023; May and June 2024). For each day, all three collection methods were used at each area (breeding pens, barn pens, and the preserve).
Five to eight resting shelter traps were sampled per area. Resting shelters were set up the day prior to sampling, in shaded areas, with the opening toward the west [25]. Traps were sampled in the morning (8:30–10:30 a.m.), as female blood-engorged mosquitoes often seek refuge, often in cavities of dark, cool places, to digest their blood meal [26, 27]. The large-diameter aspirator consisted of galvanized air duct (20.3 cm in diameter, 61 cm long) with an electric radiator fan 20 cm in diameter powered by a rechargeable 12 V/6Ah gel-sealed battery [24]. The aspirator drew mosquitoes toward the collection cup, which was a screen-bottom 3.7 L plastic food storage container (Rubbermaid, Atlanta, GA, USA). For each site, the large-diameter aspirator was used three times daily for 2-min sessions in the morning (08:30–10:30) within a 200 m radius of the CDC light trap and in low vegetation (grasses, shrubs, and fallen limbs) [28]. The small diameter aspirator consisted of a 10 cm computer fan, attached to a clear plastic tube (120 mm in diameter), powered by a rechargeable 12 V/6Ah gel-sealed battery [25]. The small-diameter aspirator was operated for three 2-min sessions in the morning (8:30–10:30 a.m.) around crevices, burrows, and hollowed trees [25]. At each site, one CDC light trap baited with carbon dioxide was set at dusk (6:30–8:30 p.m.) and collected in the morning (8:30–10:30 a.m.). The light traps were hung above the ground from a 1.21 m shepherd hook. Each light trap was baited with dry ice in an insulated thermos canister that delivered carbon dioxide to the trap through its downward-directed spout. The traps were powered by rechargeable 6 V/12Ah gel-sealed batteries (Model SLAA6-12F, Duracell Ultra, Bethel, Connecticut, USA) [27].
Mosquitoes were identified to species on the basis of morphology using dichotomous keys in Darsie and Ward (2005) and Burkett–Cadena (2013) [29, 30]. Females belonging to the Anopheles quadrimaculatus subgroup (five species) and Anopheles crucians complex (seven species) cannot be reliably differentiated morphologically, and were recorded as An. quadrimaculatus s.l. and An. crucians s.l.
The abdomen of each blood-engorged female was separated from the head–thorax segments using a sterile micropipette tip. The blood meal was smeared onto a Whatman FTA Classic Card to preserve for subsequent blood meal analysis [31, 32]. Up to 25 nonengorged females and head–thorax segments of blood-engorged females of the same species were pooled and processed using nested PCR to detect Plasmodium.
For any thoracic segment that tested positive for P. odocoilei, the corresponding blood-engorged abdomen was also screened for Plasmodium to determine the developmental stage of the parasite. Detection of Plasmodium in thoracic segments may indicate the presence of infectious sporozoites in the salivary glands [18], whereas positive abdominal segments suggest the presence of either midgut oocysts or Plasmodium-infected erythrocytes derived from the host deer.
Molecular analysis
DNA was extracted from samples using the InstaGene matrix (Biorad, Hercules, CA), following published protocols [31]. For identifying host species, vertebrate host DNA was amplified from the mosquito blood meal using the primer pair H2714 and L2513 (Table 1) targeting mitochondrial 16S rRNA and cytochrome b gene (cytb) of mammals and amphibians [22, 27, 33]. Samples that did not amplify were then processed using the L0 and H1, then L0 and H0 primer pair targeting cytb gene of birds, and then 16L1 and H3056 primer pair targeting the 16S ribosomal RNA (rRNA) of reptiles (Table 1) [34, 35]. A nested PCR assay was used to screen for the presence of P. odocoilei by amplification of Plasmodium DNA using a cytb gene [9]. A total of 2.5 µL of the resulting DNA was then used as a template for 25 µL amplification reactions. PCR amplifications were conducted in a solution containing 12.5 µL Invitrogen Platinum Green Hot Start PCR Master Mix, 0.5 µL of each primer (20 µM), 2.5 µL of DNA template, and 9 µL molecular-grade water. The nested PCR used the amplicon from an initial reaction as a template for the second reaction [36–38]. Table 1 provides primer sequences, target genes, expected amplicon sizes, and cycling conditions for all primers used. Table 1. The primers that were used and their gene targets, sequences, expected amplicon size, and PCR cycle conditionsPrimer nameTarget geneSequenceAmplicon size (bp)Cycle conditions**H2714/L251316S rRNAF: 5’-CTCCATAGGGTCTTCTCGTCTT-3’R: 5’-GCCTGTTTACCAAAAACATCAC-3’20095/240, 95/30 (35), 57/30, 72/30, 72/42016L1/H305616S rRNAF: 5’-CTGACCGTGCAAAGGTAGCGTAATCACT-3’R: 5’-CTCCGGTCTGAACTCAGATCACGTAGG-3’45095/240, 95/30 (35), 62.5/30, 72/30,72/420L0/H0cytbF: 5’-GGACAAATATCATTCTGAGG-3’R: 5’-GGGTGTTCTACTGGTTGGCTTCC-358994/300, 94/30 (35), 55/45, 72/60, 72/420L0/H1cytbF: 5’-GGACAAATATCATTCTGAGG-3’R: 5’-GGGTGGAATGGGATTTTGTC-3’22094/300, 94/30 (35), 65/45, 72/60, 72/420DW2/DW4cytbF: 5’-TAATGCCTAGACGTATTCCTGATTATCCAG-3’R: 5’- TGTTTGCTTGGGAGCTGTAATCATAATGTG-3’80094/240, 94/30 (35), 60/30, 68/50, 68/420DW1/DW3cytbF: 5’-TCAACAATGACTTTATTTGG-3’R: 5’-TGCTGTATCATACCCTAAAG-3’1,00094/60, 94/20 (40), 52/20, 68/30, 68/420^^Cycle conditions: temperature (°C), time (s), and number of cycles (in parentheses) for denaturation, annealing, and extension. Primers H2714/L2513; 16L1/H3056; L0/H0, and L0/H1 were used for host blood meal determination. Primers DW2/DW4 (outer) and DW1/DW6 (inner) were used for P. odocoilei detection. F, forward; R, reverse
Gel electrophoresis was performed to visualize the PCR products [39]. Amplicons from blood meals and Plasmodium infection assays were purified and sequenced using the Sanger method by Eurofins Genomics (Louisville, KY, USA). Nucleotide sequences were entered into the GenBank (National Center for Biotechnology Information) database using the Basic Local Alignment Search Tool (BLASTn). Only sequences with ≥ 95% identity matches determined by BLASTn were accepted for blood meal host and Plasmodium species identification [39].
Statistics
A natural vector of deer malaria must take multiple (minimum of two) blood meals from WTD in its lifetime [19]. Therefore, putative vectors must specialize in feeding on this single host species to satisfy the incrimination criteria. We considered at least 75% of blood meals from WTD (three out of four) to constitute WTD host specialization.
The infection rate of each vector species was calculated by taking the number of samples positive for P. odocoilei and dividing them by the total females screened. Each positive pool was assumed to have only one female mosquito positive for P. odocoilei [40]. The “binom.test” function implemented in R Core Team (2024) was used to calculate 95% confidence (CI) intervals using the Clopper–Pearson “exact” interval [41].
Results
Trap type
A total of 5957 female mosquitoes were collected using a combination of trapping methods, which included pop-up resting shelters, a large-diameter aspirator, a small-diameter aspirator, and a carbon-dioxide-baited miniature light trap (Fig. 1). Nine different genera were collected, including mosquitoes from the genera Aedes (8.8%, n = 523), Anopheles (8.5%, n = 509), Culiseta (43.8%, n = 2610), and Culex (36.8%, n = 2190). The most frequently collected species was Culiseta melanura, captured primarily by the large-diameter aspirator (48.9%, n = 1272), of which 12.5% (n = 159) were blood-engorged (Fig. 1). Culex erraticus constituted 32.6% (n = 1927) of total mosquitoes collected (Fig. 1), captured mainly by resting shelters (52.0%, n = 1008), with a large percentage being blood-engorged (41.3%, n = 416). The resting shelter also captured the largest percentage of Anopheles punctipennis (60.2%, n = 83) and Anopheles quadrimaculatus s.l. (53.0%, n = 89) (Fig. 1), with a large percentage being blood-engorged (59.6% and 64.6%, respectively). Most An. crucians s.l. were captured using light traps (62.2%, n = 323); however, the light traps captured a low percentage of blood-engorged An. crucians s.l. females (13.2%).Fig. 1. Relationship between trap type and physiological status for mosquito species at a deer farm in FL, USA. Traps included a large-diameter aspirator (LDA), Centers for Disease Control miniature light traps baited with carbon dioxide (LT), a pop-up resting shelter (RS), and a small diameter aspirator (SDA). A Aedes vexans; B Anopheles crucians s.l.; C Anopheles quadrimaculatus s.l.; D Anopheles punctipennis s.l.; E Culiseta melanura; F Culex erraticus
Blood meal analysis
A diverse assemblage of vertebrates was bitten by the mosquitoes sampled at the deer farm, varying substantially by mosquito species (Supplementary Table S1). A total of 632 blood meals were identified, derived from a wide range of species of the classes Aves (18 species), Mammalia (12 species), Reptilia (1 species), and Amphibia (1 species) (Fig. 1). In general, species of Anopheles took a large fraction (> 80%) of identified blood meals from mammals, particularly WTD (Fig. 2). Anopheles quadrimaculatus s.l. fed primarily on mammals (92.6%, 25/27), including a large percentage of total blood meals from WTD (88.9%; Fig. 2) and one blood meal from eastern cottontail rabbit (Sylvilagus floridanus) (3.7%). Anopheles punctipennis took 93.3% (28/30) of blood meals from mammals, including WTD (83.3%; Fig. 2), axis deer (Axis axis) (3.3%), and fallow deer (Dama dama) (3.3%). Anopheles crucians s.l. took 88.3% (53/59) of blood meals from mammals, with 85.0% of blood meals from Artiodactyla, specifically WTD (80.0%, Fig. 2), cows (Bos taurus) (3.3%), and blackbuck (Antilope cervicapra) (1.7%). Several species of songbird, particularly the northern cardinal (Cardinalis cardinalis), wood thrush (Hylocichla mustelina), red-eyed vireo (Vireo olivaceus), and common yellowthroat (Geothlypis trichas), were bitten occasionally (one blood meal or less per mosquito species) by An. quadrimaculatus s.l., An. punctipennis, and An. crucians s.l. Anopheles punctipennis (n = 1), and Anopheles crucians (n = 3) also bit the green anole lizard Anolis carolinensis (Fig. 2).Fig. 2. Host associations of selected mosquito species from a Florida deer farm. PCR-based blood meal analysis followed by Sanger sequencing was performed to determine the vertebrate host identity. Sample sizes provided in parentheses
Culex erraticus took a large percentage of blood meals from mammals (93.2%, 136/146), mostly from Artiodactyla, including WTD (87.7%, Fig. 2), nilgai (Boselaphus tragocamelus) (1.4%), and fallow deer (1.4%). Culex erraticus fed upon diverse birds (4.7% of total blood meals), including single blood meals from a mallard (Anas platyrhynchos), green heron (Butorides virescens), veery (Catharus fuscescens), yellow-rumped warbler (Setophaga coronata), common yellowthroat, wild turkey (Meleagris gallopavo), and barred owl (Strix varia). Culex erraticus also fed on an amphibian (southern toad [Anaxyrus terrestris]; 0.7%) and one reptile (green anole lizard; 1.4%).
Culiseta melanura mainly fed upon birds (64.6%, 31/48), which included red-eyed vireo (16.7%), wood thrush (14.6%), northern cardinal (6.3%), veery (4.2%), ruby-crowned kinglet (Corthylio calendula) (4.2%), yellow-rumped warbler (2.1%), yellow-billed cuckoo (Coccyzus americanus) (2.1%), white-eyed vireo (Vireo griseus) (2.1%), turkey vulture (Cathartes aura) (2.1%), tufted titmouse (Baeolophus bicolor) (2.1%), Swainson’s thrush (Catharus ustulatus) (2.1%), Carolina wren (Thryothorus ludovicianus) (2.1%), and barred owl (2.1%). A lower percentage of Cs. melanura blood meals were derived from mammals (33.3%, 16/48), including blood meals from WTD (18.8%), and single blood meals from five other mammal species (Fig. 2).
Plasmodium odocoilei infection rate
Anopheles quadrimaculatus s.l., An. punctipennis, and An. crucians s.l. were the only species found infected with P. odocoilei at the deer farm (Table 2). The highest overall P. odocoilei infection rate was observed in An. quadrimaculatus s.l. (4.1%; Clopper–Pearson exact 95% CI 4.0–4.1%), followed by An. punctipennis (3.1%; Clopper–Pearson exact 95% CI 3.1–3.2%) and then An. crucians s.l. (0.47%; Clopper–Pearson exact 95% CI 0.46–0.48%). Two other species, Cx. erraticus and Cs. melanura, had large sample sizes screened for the parasite (n = 1457 and n = 1741, respectively), but P. odocoilei was not detected in those species. Plasmodium odocoilei was detected in An. quadrimaculatus s.l., at all sampling sites, with the highest infection rate observed at the barn pens (6.7%), followed by breeding pens (3.7%) and preserve (3.3%). In contrast, the highest infection rate for An. punctipennis was observed at the barn pens (6.7%), followed by breeding pens (3.2%), but was not detected at the preserve (0/15 screened). For An. crucians s.l., P. odocoilei was only detected at the preserve (1.1%), not at the breeding pens (n = 37 screened) or barn pens (n = 70 screened). Plasmodium sequences showed a high similarity to available P. odocoilei sequences in GenBank (range = 98.7–100%; mean = 99.9%), suggesting that the Plasmodium spp. detected were indeed P. odocoilei. For most of the mosquito pools (six of seven) that tested positive for P. odocoilei, a blood meal of a female that was included in the pool also tested positive for the parasite. Two An. punctipennis pools tested positive for P. odocoilei; however, only one of the corresponding blood meals was found to be positive. Two An. quadrimaculatus s.l. pools were positive for P. odocoilei, and positive blood meals were detected for both pools. For the single An. crucians s.l. pool that tested positive, a positive blood meal was also detected. Table 2. Plasmodium odocoilei in mosquitoes from a deer farm in Gadsden County, Florida, USA (2022–2023)SpeciesP. odocoileiBreeding pensBarn pensPreserveNA**Total (CI)Anopheles crucians s.l.Females screened37709511213Positive00101Infection rate (%)001.100.47 (0.46–0.48)Anopheles punctipennisFemales screened311515364Positive11002Infection rate (%)3.26.7003.1 (3.1–3.2)Anopheles quadrimaculatus s.l.Females screened271530274Positive11103Infection rate (%)3.76. 73.304.1 (4.0–4.1)Culex erraticusFemales screened274802328531457Positive00000Infection rate (%)00000Culiseta melanuraFemales screened399286972841741Positive00000Infection rate (%)00000^^Sites marked as “NA” indicate that study locations were not recorded. Mosquitoes were sampled in three areas (breeding pens, barn pens, and preserve). Pools consisted of 25 females or fewer, screened by nested PCR-based assay targeting the cytb gene of Haemosporida followed by Sanger sequencing of amplicons
Discussion
The objectives of this study were to determine which mosquito species specialize in feeding on WTD and to quantify the infection rate of P. odocoilei in female mosquitoes, thereby partially incriminating vector(s) of deer malaria. Culiseta melanura, Cx. erraticus, An. punctipennis, An. quadrimaculatus s.l., and An. crucians s.l. were present in areas inhabited by WTD at the farm. However, only An. punctipennis, An. quadrimaculatus s.l., and An. crucians s.l. specialized in feeding on WTD and were found to be infected with P. odocoilei. These results indicate that the putative vectors for P. odocoilei include An. quadrimaculatus s.l., An. punctipennis, and to a lesser extent, An. crucians s.l..
The possibility of An. quadrimaculatus s.l. being a vector for deer malaria is supported by its high specialization in feeding on WTD (88.9%) and relatively high P. odocoilei infection rate (4.1%). Other studies across North America have shown that An. quadrimaculatus takes a relatively high percentage of its blood meals (average of 60.0% across studies) from WTD (Table 3). Of the 12 available studies, 5 found that An. quadrimaculatus specialized on WTD, taking 75–100% of total blood meals from this single host species [11–14, 42–50]. These studies varied considerably in location, from zoos and urban residences to forests and swamps, potentially explaining some of the variation in the feeding patterns of An. quadrimaculatus. The detection of Plasmodium parasites in both the head–thorax segments and the corresponding blood meals of An. quadrimaculatus s.l. suggests that these females may have been infected with both sporozoites and gametocytes. Overall, evidence of An. quadrimaculatus s.l. specializing in feeding on WTD, its natural infection with P. odocoilei, and the potential presence of both gametocytes and sporozoites support its role as a potential vector of deer malaria. Table 3. Summary of published blood meal analysis studies, with respect to feeding on white-tailed deerLocationSettingCiteAn. crucians s.l.An. punctipennis**An. quadrimaculatus s.l.Cs. melanura**Cx. erraticus(%) WTD (n)(%) WTD (n)(%) WTD (n)(%) WTD (n)(%) WTD (n)CanadaUrban/forest[14]–32 (21)21 (14)11 (9)–NYForest[13]–96.3*** (107)97.7*** (131)––CTUrban[12]–90.9*** (11)75.0*** (8)––NJRural[43]72.0 (50)60.0 (20)83.9*** (415)4.4 (68)–NJForest[13]–85.8*** (127)96.9*** (288)––NCRural[44]25.0 (4)50.0 (16)7.7 (13)0 (51)0 (13)NCUrban[45]––42.9 (550)–27.2 (169)TNZoo[46]–20.0 (5)0 (7)–5.9 (17)TNUrban[43]––14.2 (169)–10.0 (50)TNUrban[47]–70.0 (10)11.6 (249)–0.4 (454)TNForest[11]–29.0 (34)40 (112)0 (3)60.0 (82)ALRural[50]–32.4 (34)––8.0 (25)FLForest[48]22.7 (422)–55.0 (20)––FLSwamp[42]28.6 (21)–––19.7 (66)FLSwamp[49]54.5 (22)–44.1 (34)1.7 (60)19.2 (500)FLRural[51]11.1 (9)0 (5)100*** (1)0 (57)0 (24)FLRural[34]–––0 (233)–FLRural[56]0.5 (213)Total28.8 (528)71.3 (390)60.0 (2011)1.0 (694)15.3 (1400)Asterisks denote > 75% of feeding on white-tailed deer (WTD)
Anopheles punctipennis is also a likely vector of P. odocoilei, owing to specialization in feeding on WTD in this study (83.3%) and natural infection (3.1%) with P. odocoilei in field-caught samples. Other studies showed that An. punctipennis also takes a relatively high percentage of blood meals from WTD in nature (average of 71.3%). Of 12 available blood meal analysis studies, 3 reported that An. punctipennis is specialized in feeding on WTD, taking 75–100% of total blood meals from this single host species [11–14, 43–51]. The detection P. odocoilei in the head–thorax and the corresponding blood meals of An. punctipennis suggest that sporozoites and gametocytes could be present. The absence of P. odocoilei in blood meals from females whose bodies were pooled suggests that some females harbored sporozoite stage, potentially indicating infectious vectors. The likelihood of An. punctipennis being a vector for P. odocoilei is also supported by a study from Washington, DC, [9] showing a similar infection rate of P. odocoilei in An. punctipennis (5.0%) as in our study (3.1%).
Anopheles crucians s.l. may serve as a vector for P. odocoilei. However, owing to its relatively low P. odocoilei infection rate in our study (0.47%), this species likely does not play a major role in transmission. Although An. crucians took an average of 28.8% of its total blood meals from WTD in other studies (Table 3), this species was not observed to specialize in feeding on WTD in the seven available studies [43–45, 49, 50]. The low infection rate of P. odocoilei, low percentage of blood meals from WTD, and the Plasmodium-infected blood meal suggest that An. crucians s.l. plays a minor role in the transmission of deer malaria.
Other common species of mosquitoes observed in our study, particularly Cx. erraticus and Cs. melanura, are unlikely vectors of P. odocoilei. Although Cx. erraticus showed a high specialization in feeding on WTD in our study (87.7%), it did not specialize in feeding on WTD in nine other studies, where an average of 15.3% of its blood meals came from WTD [11, 42–47, 49–51]. In contrast, Cs. melanura mainly fed on songbirds on the deer farm in our study. Other studies found that Cs. melanura does not specialize in feeding on WTD either (1.0% on average) [11, 14, 34, 42–44, 49, 51]. Neither Cx. erraticus nor Cs. melanura were found infected with P. odocoilei, despite over 1000 females of each species being screened for the parasite. Given the low percentage of blood meals from WTD and lack of infection, both species are not likely vectors of deer malaria.
This study was conducted at a single deer farm in Gadsden County, which may limit the application of findings to other regions. In addition, females of the An. quadrimaculatus complex and An. crucians complex are difficult to reliably identify morphologically, so the dominant member within each group is unclear. Future research could use DNA barcoding to determine species composition at the deer farm and assess whether host use or P. odocoilei infection rates vary among members of each group.
Despite our efforts to separate thoraxes and abdomens when processing samples, PCR-based detection of P. odocoilei is unable to specifically determine the stage of the parasite. Detection of P. odocoilei in the thorax could be attributed to the amplification of infected blood meal residue, although this is unlikely owing to degradation by mosquito digestive enzymes [52, 53]. Alternatively, it is possible that PCR amplification represented underdeveloped oocysts or nonviable sporozoites present in the hemocoel [53, 54]. Future studies could perform dissections and microscopy, as well as develop molecular markers to target parasite stage-specific proteins, to clarify whether or not PCR positives represent infectious mosquitoes [52, 53]. Despite these limitations in the collection and blood meal analysis, the study successfully identified mosquitoes specializing in feeding on WTD at the study site and detected the P. odocoilei in wild-collected adult females of three mosquito species.
Conclusions
This study is important for partially incriminating the vectors of P. odocoilei and understanding the transmission of deer malaria. Anopheles quadrimaculatus s.l. and An. punctipennis better met criteria for vector incrimination than An. crucians s.l.. Their relative importance as vectors of P. odocoilei likely varies with geographic region and habitat. Anopheles punctipennis is more common in temperate woodlands, while Anopheles quadrimaculatus s.l. is associated with forested wetlands [29]. Future studies should evaluate the vector competence of An. quadrimaculatus s.l.., An. punctipennis, and An. crucians s.l. for transmitting P. odocoilei in laboratory settings. If vector competence is confirmed, IPM strategies that focus on the vector species can be developed to decrease the risk of infection in WTD, particularly on deer farms [55].
Supplementary information
Supplementary material 1.