Shell dissolution rates differ fourfold between mussel species
Rachel R. Carlson, Mazie A. Lewis, Aaron T. Ninokawa, Alisha M. Saley, Tessa M. Hill, Brian Gaylord

TL;DR
This study shows that mussel species differ in how quickly their shells dissolve in acidified ocean conditions, with implications for coastal ecosystems.
Contribution
The study reveals a fourfold difference in shell dissolution rates between two mussel species under ocean acidification.
Findings
M. trossulus has fourfold higher shell dissolution rates than M. californianus under ocean acidification.
Sealing the shell interior significantly reduces dissolution rates in both species.
Freshwater-tolerant mussels may face greater challenges under ocean acidification due to higher abiotic dissolution.
Abstract
Ocean acidification poses a critical threat to marine calcifiers globally and is particularly severe in the California Current System, where ecologically and economically important bivalves experience reduced calcification under climate change. Marine mussels display differential habitat preferences, with species like Mytilus californianus favouring fully saline environments and M. trossulus inhabiting sites with greater freshwater input. Determining abiotic dissolution rates of these species under ocean acidification is essential for predicting future consequences of climate change for coastal populations. We examined shell dissolution rates of mussel congeners under a range of pH (6.5–9.3) and aragonite saturation states (0.1–9.0). We also experimentally quantified the relative importance of dissolution from interior versus exterior shell surfaces. M. trossulus exhibited fourfold…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
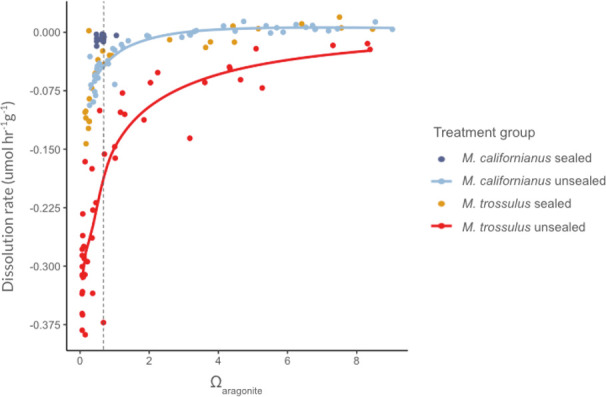 Figure 1
Figure 1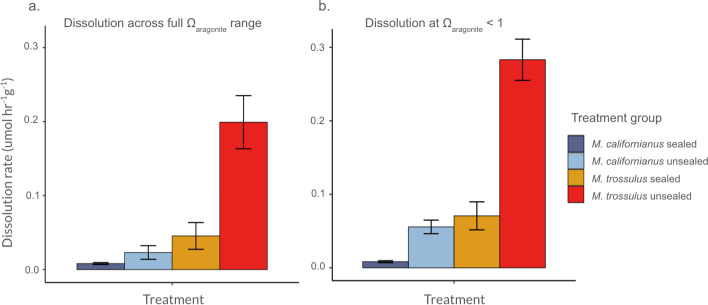 Figure 2
Figure 2|
sample size |
collection site |
mean autumn salinity at collection site |
mean dry weight (g) |
mean length (mm) |
mean treat. time (h) |
maximum dissolution rate (µmol hr−1 g−1) |
dissolution rate at Ω < 1 (µmol h−1 g−1) | |
|---|---|---|---|---|---|---|---|---|
|
|
45 |
Penn Cove, WA, USA |
21.1 ± 5.3 [ |
4.1 ± 1.0 |
58.9 ± 5.2 |
43.2 ± 0.6 |
|
|
|
|
27 |
Penn Cove, WA, USA |
21.1 ± 5.3 [ |
3.6 ± 0.8 |
54.1 ± 4.5 |
43.7 ± 0.8 |
|
|
|
|
46 |
Carmet, CA, USA |
33.2 [ |
14.4 ± 5.4 |
NA |
111.1 ± 2.5 |
|
|
|
|
25 |
Carmet, CA, USA |
33.2 [ |
10.5 ± 1.9 |
54.2 ± 4.2 |
49.2 ± 2.2 |
|
|
|
|
s.e. |
| |
|---|---|---|---|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
intercept |
−3.99 |
0.15 |
<0.0001 |
- —Division of Ocean Scienceshttp://dx.doi.org/10.13039/100000141
- —Chancellor's Postdoctoral Fellowship, University of California
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsOcean Acidification Effects and Responses · Marine Bivalve and Aquaculture Studies · Marine Biology and Ecology Research
Introduction
Ocean acidification is increasing worldwide due to elevated carbon dioxide in the atmosphere, leading to modifications in the ocean carbonate system and, consequently, alterations in the shell structure of calcifying organisms like marine mussels [1–3]. Ocean acidification is particularly severe in the California Current of the northeast Pacific Ocean, where pH is decreasing twice as rapidly as the global average [4–6] and nearshore aragonite saturation state (Ω_aragonite_) is expected to move outside of its current variability envelope by the mid−2030s [7]. Significant changes to calcifying organisms have been documented due to ocean acidification in this region including impaired calcification (e.g. mussel shell thinning [8–10]), modified larval development and reproduction (e.g. [11,12]) and altered behaviour or susceptibility to predation (e.g. [13–15]) that could disrupt species assemblages and trophic structure in critical fisheries [16–19].
Mussels are important habitat-forming organisms that are highly susceptible to ocean acidification. Mussels generate rugose beds that increase benthic surface area in the rocky intertidal zone, forming climatic niches for epibionts and small benthic organisms [20] and support aquaculture and coastal fisheries (e.g. [21,22]). Many mussel species in the California Current are vulnerable to ocean acidification, posing serious risks to benthic ecosystems [8,23–25]. Two congener mussels, M. trossulus and M. californianus, are common in the California Current yet inhabit divergent habitat niches. M. trossulus is abundant in the Pacific Northwest but rare in California and thrives in regions characterized by high freshwater exposure such as the Puget Sound, and is likewise abundant in the Baltic Sea [26]. In contrast, M. californianus spans from northern Mexico to Alaska and prefers typical marine salinity levels of 32−37 ppt [10,21]. Given that fresh water sources vary in alkalinity, which alters the relationship between dissolved carbon dioxide and pH, M. trossulus and M. californianus may experience differential susceptibility to dissolution when their respective habitats are subjected to increased carbon dioxide.
Phenotypic differences between M. trossulus and M. californianus may also affect vulnerability to ocean acidification. M. californianus was recently characterized as the only Mytilid species to precipitate three mineralogical layers—outer calcite, middle aragonite and inner calcite—whereas other Mytilus species precipitate two layers: an outer layer of prismatic calcite and an inner layer of nacreous aragonite [10]. Inner-shell mineralogy may strongly affect vulnerability to ocean acidification, as the inner shell of mussels may be more susceptible to high pCO_2_ than the outer shell due to the fact that the inner shell lacks a periostracum (protective outer layer [27]). While the outer shell and periostracum are exposed to seawater, the inner shell is exposed to extrapallial fluid that has reduced pH and carbonate, associated with diffusive ejection of metabolic pCO_2_ by mussels that exacerbates acidified conditions at the inner shell compared to seawater [28,29]. Fitzer et al. [30] found that, under conditions of ocean acidification, calcite outer shells in M. edulis became more brittle, while aragonite inner shells became softer and more pliable. Melzner et al. [27] found that M. edulis, which is similar in morphology and habitat to M. trossulus, exhibited high internal shell dissolution under elevated seawater pCO_2_ and low-food treatments, though food subsidies helped mussels overcome high pCO_2_ stress. However, no study to our knowledge has directly compared the effects of ocean acidification on mussel shells with differential shell structure.
In this study, we seek to address the following research questions. First, how do abiotic dissolution rates differ between taxonomically similar calcifiers (congeners) with different mineralogical composition (M. trossulus and M. californianus)? Second, if differences in abiotic dissolution are observed between congeners, to what extent does the shell interior versus exterior contribute to this difference (i.e. which shell surface introduces heightened vulnerability)? (Note: throughout this article, we use ‘abiotic dissolution’ to describe shell loss independent of the biological activity of live mussels or shell-associated microbes.) We discuss possible implications of results for organism vulnerability to ocean acidification. We also contextualize results in the divergent environments (levels of fresh water input) these organisms typically inhabit.
Methods
Mussel collection and preparation
2.1.
Naturally settled M. californianus were manually collected from Carmet Beach, CA, USA between January 2020 and April 2022 [31]. Salinities at a monitoring station in the vicinity of Carmet Beach exhibit a mean of 33.2 ± 0.4 standard deviation [32]. Mussels were hand-selected at lengths ranging from 30 to 80 mm from the mid-intertidal zone and samples were gathered from the same location to draw from a consistent population and set of environmental parameters that can determine phenotypic traits. Mussels were dissected immediately upon arrival at Bodega Marine Laboratory (BML), CA (<0.5 h transit time) to remove all body tissue. Remaining organic materials (byssal threads, epibionts) were removed by drying shells in an oven for 24 h at 60°C.
M. trossulus of 20–75 mm length (target length consistent with M. californianus shells) were collected from Penn Cove Shellfish Farm in Penn Cove, WA, USA in September and December of 2023. Penn Cove, which uses natural seed, catching and growing wild larvae on rope substrate suspended from rafts, is located in the Salish Sea, which receives significant fresh water from the Skagit River and other waterways in British Columbia and the Pacific Northwest including the Whidbey Basin watershed. Monitoring data at the farm shows a mean autumn salinity of 21.1 ± 5.3 and mean pH of 7.79 ± 0.24 at 1 m depth [33]. Mussels from Penn Cove were maintained in a moist, cool and insulated environment during shipment to BML (< 1 day), and were dissected immediately upon arrival at the lab using methods described above.
To separate contributions of the inner or outer shell surface to dissolution, we coated the inside of a subset of shells of each species with a clear, silicone waterproof sealant (Loctite), applying a single, thin layer to the nacre. A total of 49 M. trossulus shells were sealed and 73 were unsealed; and 74 M. californianus shells were sealed and 46 were unsealed. Sealed M. californianus shells were completed by Saley & Gaylord [34,35] but, given that these authors were attempting to sample across a wide range of periostracum percent cover, we used only shells with a comparable percentage of periostracum to other treatment groups (>50% periostracum; 25 of original 74). Periostracum in Saley & Gaylord [35] was determined through digital photographs analysed in ImageJ (software version 1.52a); periostracum in other treatments was estimated >50% based on visual inspection. The sealant was left to dry for >48 h prior to incubation and all shells were then inspected to ensure full seal and soaked in ambient seawater for 1 h immediately prior to incubation (described below) to allow seawater to saturate any possible air pockets in the sealant. Trials conducted in advance demonstrated that cured sealant had no detectable effect on seawater chemistry [35].
Mussel treatment
2.2.
Abiotic dissolution experiments were conducted on mussel shells between March 2020 and March 2024. All mussel shells were incubated in seawater manipulated to a target aragonite saturation state of Ω_aragonite_ = 0–9 (actual Ω_aragonite_ = 0.05–9.03). Seawater was derived from the Bodega Marine Lab seawater flow-through system, sourced from an intake located 60 m offshore of Horseshoe Cove, CA at a depth of 2 m below mean low tide, and sand filtered to 30 microns. Incubations were conducted in dark conditions at a constant temperature of mean 12.4°C, corresponding to ambient seawater temperature during the season when incubations were conducted. We first added ambient seawater to a 1 l mixing vessel, then added variable doses of sodium hydroxide (NaOH) and hydrochloric acid (HCl) to manipulate pH and Ω_aragonite_. We mixed the treatment water thoroughly and subsampled 150 ml of this water to characterize chemical conditions before incubation (described below). The remaining 850 ml of treatment water was inverted into a glass incubation jar containing a mussel shell and the jar was sealed immediately and placed in a dark incubation chamber. While dark conditions are primarily used to control light conditions in live-mussel experiments, several incubations occurred in tandem with live incubations in darkness for a separate experiment and therefore dark conditions were maintained throughout our experiment for consistency. We removed mussel shells after a target of 42−45 h, though 25% of incubations occurred in tandem with prior experiments and had an incubation period of 108−115 h; we accounted for this difference by normalizing abiotic dissolution by time (dissolution rate). After incubation, a 150 ml subsample was again extracted from jars and used to measure chemical conditions after incubation. The treatment water mass and shell mass were recorded, with water mass derived using the equation (total mass = jar mass + shell mass +water mass).
Before and after incubation, we measured oxygen, temperature, salinity, total alkalinity and pH in each incubation vessel. Ammonia was measured before incubation from one of four carboys used to dispense ambient seawater for treatments (triplicate ammonia samples per carboy) and after incubation from each incubation vessel. Oxygen and temperature were measured using a PreSens Microx 4 micro-optode; salinity with a Horiba Laqua PC 1100 conductivity probe; and pH with a Horiba Laqua PC 1100 instrument, with all four probes simultaneously submerged in a 150 ml subsample. pH measurements were calibrated on the total scale using simultaneous spectrophotometry on a 3 ml sample within our temperature-controlled room using m-cresol purple dye, which is sensitive to pH changes [36]. Before and after incubation, our 150 ml subsamples were preserved in duplicate, using opaque bottles for alkalinity titration, which occurred within 24 h according to methods described in Ninokawa et al. [31]. Duplicate samples that displayed a standard deviation >10 were discarded, resulting in 45 unsealed and 27 sealed M. trossulus shells and 46 unsealed and 25 sealed M. californianus shells used for analysis (table 1). Though waste excretion was not expected for abiotic shells, we measured ammonia to account for any biologically associated changes in alkalinity (i.e. microbial activity in water or on shells) using a salicylate spectrophotometric assay, as described in Ninokawa et al. [31]. Finally, we calculated abiotic dissolution rate using the ammonia-corrected alkalinity anomaly technique [37], dividing CaCO_3_ dissolution by incubation time and dry shell mass. We used chemical measurements to determine the carbonate chemistry of each incubation including Ω_aragonite_ and Ω_calcite_ using the package seacarb v 3.3.3 in R v 4.3.3 with constants from Lueker et al. [38].
Sealed M. californianus shells were tested during a separate experiment by Saley & Gaylord [34,35] using the same analytical methods but targeting a narrow range of Ω_aragonite _< 1 and using approximately 300 ml of water. Therefore, in statistical analyses focused on comparisons across groups, our full dataset was subsampled to Ω_aragonite < 1 for consistency. We also normalized dissolution (in µmol kg^−1^) by water mass, multiplying alkalinity change by kg treatment water before finding dissolution rate per shell mass (µmol CaCO_3 hr^−1^ g^−1^) across all treatments. Note that shell length but not mass was measured by Saley & Gaylord [35]; we therefore derived shell mass for this group based on the relationship between length and mass in a separate dataset of 558 M. californianus mussels initially measured for Ninokawa et al. [31].
Statistical analysis
2.3.
To evaluate the difference between abiotic dissolution rate of two congener species, we used bootstrapping to compare dissolution rate per shell mass (µmol hr^−1^ g^−1^) by sampling with replacement from M. trossulus and M. californianus shell incubations with 10 000 iterations. Bootstrapping allowed us to estimate 95% confidence intervals (CIs) of dissolution rate from each group, where non-overlapping 95% CIs indicated significant differences between congeners. We conducted this analysis on both our full dataset (full range of Ω_aragonite_) and on a subset corresponding to Ω_aragonite < 1 (net dissolution expected). We recognize that values for saturation state would be different if we used Ω_calcite, but have chosen Ω_aragonite_ as a convenient single index to compare dissolution rates, as these indices only differ by the apparent solubility product constant yet are affected similarly by variations in seawater carbonate ion concentration. Additionally, dissolution can be observed in biogenic carbonates when Ω > 1 due to natural elemental and structural heterogeneity [39] and so we focus on the ability of Ω_aragonite_ to primarily reflect changes in carbonate ion concentrations in this experiment.
To determine the contribution of the inner shell layer to dissolution, we conducted bootstrapping analysis on sealed shells (interior coated with silicone waterproof sealant) versus unsealed shells from each species. Finally, to account for slightly variable Ω_aragonite_ between groups, we used linear regression to examine the influence of Ω_aragonite_, mussel species, treatment (sealed or unsealed) and the interaction between treatment and species on dissolution rate after subsetting data to Ω_aragonite _< 1 and log-transforming our dependent variable to account for strong positive skewness. As a sensitivity test, we assessed the model with and without the removal of three high-leverage observations based on hat values and Cook’s distance. Removing these points did not substantially change model coefficients or statistical significance. Diagnostic plots confirmed that our model met assumptions of homoscedasticity and normality of residuals. Results are reported from the full model using all observations.
Results
We refer below to dissolution as the inverse of calcification, and therefore, higher dissolution values represent faster dissolution and negative values represent shell precipitation. For both M. californianus and M. trossulus shells, abiotic dissolution rate decreased with an increase in Ω_aragonite_ across all treatment groups where Ω_aragonite_ varied (figure 1). There were distinct differences between abiotic dissolution curves of M. trossulus and M. californianus shells (unsealed), with M. trossulus displaying a maximum dissolution rate of 0.39 µmol hr^−1^ g^−1^ at Ω_aragonite_ = 0.15 and M. californianus displaying a maximum dissolution rate of 0.09 µmol hr^−1^ g^−1^ at Ω_aragonite_ = 0.28 (table 1). That is, M. trossulus abiotic dissolution rate was four times that of M. californianus at the lowest target Ω_aragonite_ in our treatments. In addition, while M. californianus shells approached an asymptotic calcification rate of approximately 0 µmol hr^−1^ g^−1^ at Ω_aragonite_ > 1, M. trossulus shells displayed a calcification rate < 0 across all Ω_aragonite_ for unsealed shells, i.e. no calcification asymptote within the tested Ω_aragonite_ range (maximum Ω_aragonite_ = 9.03).
M. trossulus dissolution (unsealed shells) exceeds dissolution observed in all other treatments. Abiotic dissolution rate is shown across a range of aragonite saturation state (Ωaragonite) for (light blue) unsealed M. californianus, (red) unsealed M. trossulus, (dark blue) sealed M. californianus with an artificially coated interior shell and (yellow) sealed M. trossulus with an artificially sealed interior shell. Units reflect normalization of dissolution rate by shell mass. While the x axis represents Ωaragonite, the dotted line indicates the point where Ωcalcite = 1 for reference. Sealed M. californianus were derived from a separate experiment [34] and thus data are not available across a range of Ωaragonite.
When comparing dissolution rates across groups at Ω_aragonite_ < 1 , there was a significantly higher dissolution rate in M. trossulus (unsealed) than M. californianus (unsealed) shells, with non-overlapping CIs of, respectively, 0.28 ± 0.03 and 0.06 ± 0.01 (figure 2b). This finding was consistent when we conducted bootstrapping with sampling from shells across the full range of Ω_aragonite_ versus only shells at Ω_aragonite_ < 1 (figure 2a,b). When comparing shells with an artificially sealed and unsealed interior surface in each species, we found that unsealed M. trossulus shells had a significantly higher dissolution rate than sealed M. trossulus shells (0.28 ± 0.03 and 0.07 ± 0.02). Likewise, unsealed M. californianus shells had a significantly higher dissolution rate than sealed M. californianus shells (0.06 ± 0.01 and 0.01 ± 0.00; figure 2b). We note that the constrained range in saturation state for sealed M. californianus limits the strength of inference regarding this treatment group. More important is the finding that while M. trossulus shells displayed up to 4x higher abiotic dissolution rate than M. californianus shells, this effect disappeared when the M. trossulus inner shell was artificially sealed.
Unsealed M. trossulus shells exhibit a significantly and fourfold higher abiotic dissolution rate than M. californianus, and a significantly higher dissolution rate than M. trossulus with an artificially sealed inner shell. Figure displays mean abiotic dissolution rate (± 95% confidence interval) across treatment groups based on bootstrapping from within-group dissolution rate for (a) shells treated across the full experimental range of aragonite saturation state (Ωaragonite) and (b) shells treated with Ωaragonite < 1. Units reflect normalization of dissolution rate by shell mass. Note that for sealed M. californianus (dark blue) shells, all but one shell (Ωaragonite = 1.04) were treated with Ωaragonite < 1, and therefore, sealed M. californianus data presented in (a) and (b) are nearly identical. (b) allows for inter-group comparison at Ωaragonite < 1.
Model results showed a similar pattern, as species (M. trossulus default) showed a significant relationship with abiotic dissolution rate (β = 1.86 ± 0.12; p < 0.0001; table 2). Treatment (unsealed) also showed a positive and significant relationship with dissolution rate (β = 1.76 ± 0.11; p < 0.0001). In other words, M. trossulus was positively linked to higher dissolution, as was exposure of the interior shell in both species. However, the interaction between treatment and species (unsealed M. trossulus default) had a negative relationship with dissolution rate (β = −0.64 ± 0.16; p < 0.001). In addition, Ω_aragonite_ showed a negative relationship with dissolution rate (β = −1.42 ± 0.22; p < 0.0001), indicating that dissolution increased at low values of Ω_aragonite_ (table 2).
Discussion
Two mussel species with overlapping ranges in the California Current System exhibit strongly different abiotic dissolution rates under conditions simulating ocean acidification. M. trossulus abiotic dissolution exceeds M. californianus dissolution by a factor of four at Ω_aragonite_ approaching 0. This result suggests that M. trossulus, a marine mussel that is frequently found within or near freshwater runoff, may be particularly vulnerable to abiotic dissolution while also susceptible to changes in coastal carbonate chemistry associated with freshwater runoff. While net dissolution in nature is also a function of mussel biological control, including compensatory mechanisms that may result from feeding [40], these results provide an early step towards characterizing calcifier susceptibility to ocean acidification in critical coastal habitats.
Abiotic dissolution of M. trossulus is significantly higher in the inner shell. After artificially sealing the inner shell of M. trossulus, this species showed significantly reduced dissolution relative to unsealed M. trossulus. This pattern is consistent in M. californianus shells, perhaps because the outer shell is protected from dissolution by the periostracum [27,35] and therefore sealing the inner shell prevents substantial dissolution in both species. There was a negative interaction between M. trossulus and treatment (unsealed), suggesting that exposing the inner shell was slightly less deleterious for M. trossulus than M. californianus. However, this result is uncertain given the limited range of Ω_aragonite_ explored for sealed M. californianus shells. In addition, M. trossulus exhibited a higher difference in dissolution rate between sealed and unsealed treatments relative to M. californianus (difference of 0.21 µmol hr^−1^ g^−1^ for M. trossulus and 0.05 µmol hr^−1^ g^−1^ for M. californianus between sealed/unsealed treatments at Ω_aragonite_ < 1), indicating that sealing the interior prevented more overall CaCO_3_ loss in M. trossulus. M. californianus is the only known Mytilus species to contain a separate exterior and interior calcite layer in addition to a third, inner aragonite layer [10], a structure that may lead to less dissolution overall for this species. It is well established that aragonite has a higher solubility than calcite (e.g. [41]), and therefore, the additional calcite layer in M. californianus may confer additional protection from dissolution.
M. trossulus is typically found in the mouths of rivers or other areas with low or highly variable salinity. For example, M. trossulus is common in the Puget Sound and the coastline of the Olympic Peninsula, which receives runoff from the Elwha, Skagit and other large river systems, and represents 70% of animal biomass living on hard sediment in the Baltic Sea, a low salinity system [42]. As ‘wet regions get wetter’ under climate change [43], marine mussels like M. trossulus and M. edulis that have affinities for lower salinity habitats may be particularly susceptible to reduced calcification given the combined effects of ocean acidification and decreased salinity; alternatively, these organisms may be accustomed to salinity and pH flux (e.g. down-regulating metabolism under suboptimal conditions [40,44]). Salinity generally has a positive relationship with calcification given that low salinity from riverine catchments with typical alkalinity is marked by decreased Ca^2+^ and concentration [45]. For example, high-flow systems with high ionic dilution often exhibit low alkalinity [46,47], which Telesca et al. [48] suggest is a driver of thin shell structure in M. trossulus and M. edulis in freshwater-impacted regions [48]. One previous study also showed a negative correlation between salinity and percentage shell aragonite in M. edulis in the freshwater-exposed Puget Sound and San Francisco Bay [49]. However, freshwater alkalinity also depends on local lithology; for example, limestone-dominated watersheds and aquifers may generate higher alkalinity in freshwater systems, especially if chemical weathering is stimulated by acid deposition [50,51].
The combination of low salinity and low alkalinity in some freshwater regions may compromise mussel calcification under climate change; however, we speculate that in many cases fresh water provides ample food subsidies to help M. trossulus overcome limitations on calcification. Our study measured abiotic dissolution (i.e. it did not account for biogenic plasticity in response to stress). Feeding has been shown to significantly reduce nacre dissolution under high pCO_2_ conditions [27] and there is high biological control over calcification in the inner shell based on the mussel energy budget. Food subsidies may otherwise contribute to compensatory mechanisms under ocean acidification (e.g. increasing protective membranes and accelerating calcium and proton pumps [52]). In previous studies, pH and salinity have had variable effects on the periostracum, suggesting that mussels acclimatize to low pH and salinity in the coastal zone [53].
It is uncertain whether food subsidies are sufficient to compensate for both ocean acidification under climate change and ‘business as usual’ threats to calcification in fresh water-exposed areas (low salinity, low riverine alkalinity), and experiments testing interactive effects of salinity, pCO_2_ and feeding are needed to test realistic portfolios of stress facing nearshore calcifiers. In addition, the ecological implications of compensatory mechanisms under high-food treatments require further study. For example, Telesca et al. [48] showed that, as salinity decreased, wild-collected mussels exhibited thinner shells with a thicker periostracum and an increased proportion of calcite and organic matrix, a structure that protects mussels from dissolution but increases vulnerability to predators. M. californianus shells had mean dry weights that were three times higher than M. trossulus shell weights, indicating thicker shells associated with an additional calcite layer (table 1). If increasing shell thickness occurs through a calcite layer, this might reduce vulnerability to predators in M. californianus indicating a compensatory mechanism combating both predation and ocean acidification. Moreover, large size classes of M. californianus have demonstrated a positive relationship between seawater temperature and percentage aragonite in shells [49]. Further research is needed to determine how compensating for climate stress (ocean acidification and thermal stress) affects trophic interactions and performance across multiple life history stages and size classes in calcifiers. In addition, policy analyses similar to those initiated in tropical systems (e.g. [54]) are needed to identify local- and regional-scale regulations (e.g. water quality or land-use measures) that may alleviate multiple stressors on freshwater-exposed temperate species under climate change.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Doney SC, Balch WM, Fabry VJ, Feely RA. 2009 Ocean acidification: a critical emerging problem for the ocean sciences. Oceanography 22, 16–25. (10.5670/oceanog.2009.93) · doi ↗
- 2Kroeker KJ, Kordas RL, Crim RN, Singh GG. 2010 Meta‐analysis reveals negative yet variable effects of ocean acidification on marine organisms. Ecol. Lett. 13, 1419–1434. (10.1111/j.1461-0248.2010.01518.x)20958904 · doi ↗ · pubmed ↗
- 3Chan F et al. 2017 Persistent spatial structuring of coastal ocean acidification in the California current system. Sci. Rep. 7, 2526. (10.1038/s 41598-017-02777-y)28566727 PMC 5451383 · doi ↗ · pubmed ↗
- 4Gruber N, Hauri C, Lachkar Z, Loher D, Frölicher TL, Plattner GK. 2012 Rapid progression of ocean acidification in the California current system. Science 337, 220–223. (10.1126/science.1216773)22700658 · doi ↗ · pubmed ↗
- 5Osborne EB, Thunell RC, Gruber N, Feely RA, Benitez-Nelson CR. 2020 Decadal variability in twentieth-century ocean acidification in the California current ecosystem. Nat. Geosci. 13, 43–49. (10.1038/s 41561-019-0499-z) · doi ↗
- 6Wootton JT, Pfister CA, Forester JD. 2008 Dynamic patterns and ecological impacts of declining ocean p H in a high-resolution multi-year dataset. Proc. Natl Acad. Sci. USA 105, 18848–18853. (10.1073/pnas.0810079105)19033205 PMC 2596240 · doi ↗ · pubmed ↗
- 7Hauri C et al. 2013 Spatiotemporal variability and long-term trends of ocean acidification in the California current system. Biogeosciences 10, 193–216. (10.5194/bg-10-193-2013) · doi ↗
- 8Gaylord B, Hill TM, Sanford E, Lenz EA, Jacobs LA, Sato KN, Russell AD, Hettinger A. 2011 Functional impacts of ocean acidification in an ecologically critical foundation species. J. Exp. Biol. 214, 2586–2594. (10.1242/jeb.055939)21753053 · doi ↗ · pubmed ↗