Heterologous biosynthesis of cotylenol and concise synthesis of fusicoccane diterpenoids
Ye Yuan, Zhenhua Guan, Xue-Jie Zhang, Nanyu Yao, Wenling Yuan, Yonghui Zhang, Ying Ye, Zheng Xiang

TL;DR
Researchers developed a new method to produce fusicoccane diterpenoids using engineered fungi and chemical steps, enabling easier study of these compounds.
Contribution
A novel biosynthetic and chemical strategy for concise fusicoccane diterpenoid synthesis is introduced.
Findings
Cotylenol was successfully produced in an engineered Aspergillus oryzae strain.
Three fusicoccane diterpenoids were synthesized in 4-5 chemical steps from brassicicene I.
The method provides a foundation for preparing diterpenoids and their analogs for biological studies.
Abstract
A novel strategy for the synthesis of fusicoccane diterpenoids is reported. By harnessing the biosynthetic pathways of brassicicenes and fusicoccins, cotylenol was produced in an engineered Aspergillus oryzae strain. We further achieved the concise synthesis of three fusicoccane diterpenoids, including alterbrassicicene E and brassicicenes A and R in 4 or 5 chemical steps from brassicicene I. This strategy lays the foundation for the preparation of fusicoccane diterpenoids and their analogues for biological studies.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
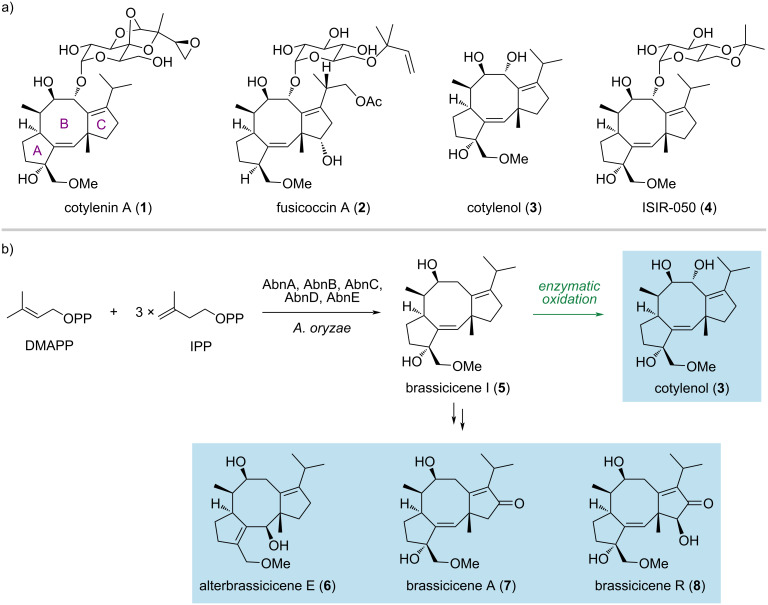 Figure 1
Figure 1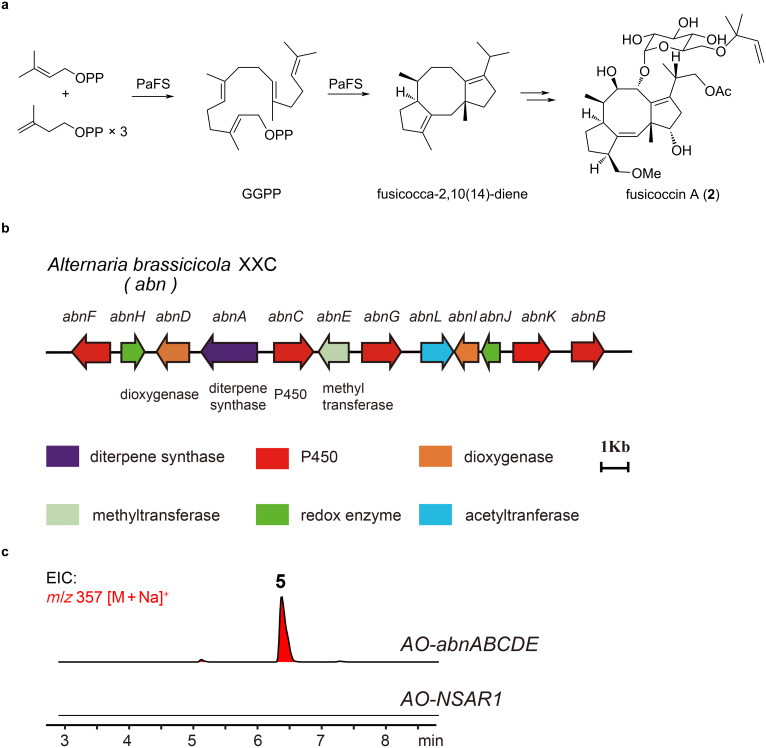 Figure 2
Figure 2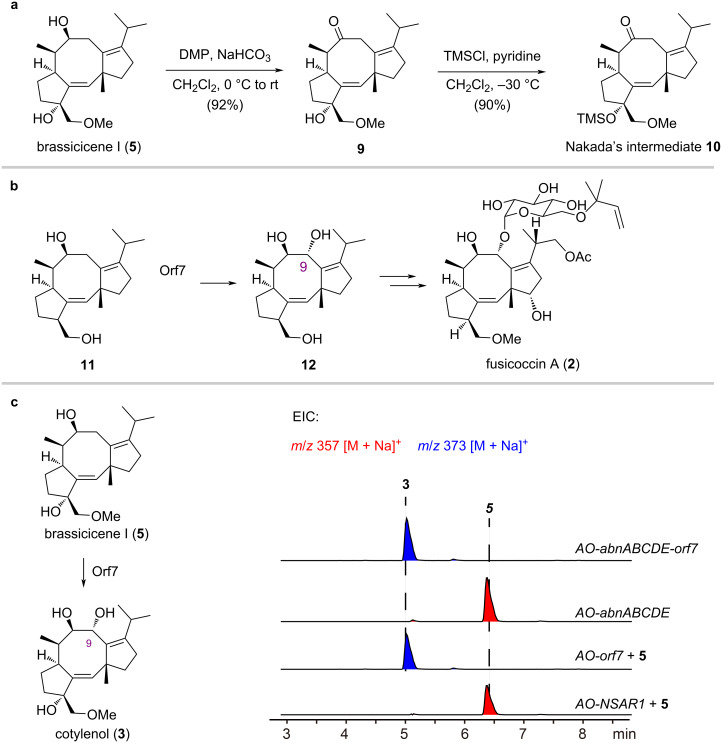 Figure 3
Figure 3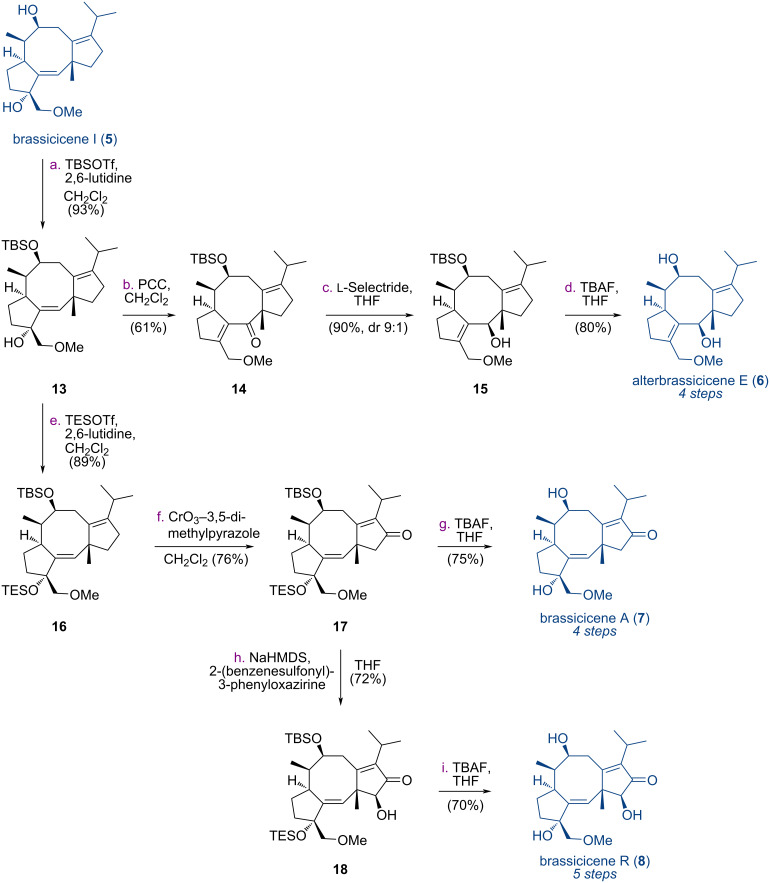 Scheme 1
Scheme 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
Topics14-3-3 protein interactions · Phytochemical compounds biological activities · Ubiquitin and proteasome pathways
Introduction
Fusicoccanes are a family of 5-8-5 tricyclic diterpenoid natural products that are produced by bacteria, fungi, algae, and plants (Figure 1a) [1–7]. Fusicoccanes possess a broad range of biological activities, including anticancer, anti-inflammatory, antimicrobial, antiparasitic, and plant growth regulating activities. For instance, cotylenin A (1) and fusicoccin A (2) function as molecular glues to stabilize the interactions between 14-3-3 proteins and their binding partners in plant and animal cells [8–12]. It has been reported that cotylenin A and its aglycone, cotylenol (3), induce differentiation in murine and human myeloid leukemia cells [13]. Cotylenin A and fusicoccin A also act synergistically with interferon-α or rapamycin to induce apoptosis in cancer cell lines [14–16]. However, cotylenin A cannot be produced by its natural source, Cladosporium sp. 501-7W, due to the loss of its ability to proliferate during preservation [17]. The important biological activities and complex structures of fusicoccane diterpenoids have inspired several total syntheses, which range between 15 and 29 steps [18–26]. Most of these synthetic approaches rely on similar strategies, i.e., coupling of the A ring and the C ring followed by the formation of the B ring. Additionally, the semisynthesis of analogues of 1 has been reported and led to the discovery of ISIR-050 (4), which shows higher activity than cotylenin A in cell growth inhibition assays and less toxicity in single-agent treatments [27–28]. Recently, Jiang and Renata described a chemoenzymatic approach that combines the skeletal construction by chemical methods and enzymatic C–H oxidations [29]. The synthesis employs a catalytic Nozaki–Hiyama–Kishi reaction and a one-pot Prins cyclization/transannular hydride transfer to construct the 5-8-5 tricyclic scaffold. Enzymatic oxidations were used to install the hydroxy group at the C-3 position. Ten fusicoccanes were synthesized in 8–13 steps each. Despite these efforts, a strategy with limited chemical transformations is highly desirable and should enable the discovery of new fusicoccane derivatives with improved biological activity.
Selected fusicoccane diterpenoids and overview of this study. (a) Representative members of the fusicoccane diterpenoid family. (b) This work: heterologous biosynthesis of cotylenol (3) in an engineered Aspergillus oryzae strain and concise synthesis of fusicoccane diterpenoids.
Inspired by the biosynthetic machinery of terpenoids, we have reported a hybrid synthetic strategy for accessing bioactive terpenoids by combining enzymatic terpene cyclization and chemical synthesis [30–33]. Briefly, the carbon scaffolds are forged by terpene cyclases, followed by concise chemical transformations to yield the desired natural products. Here, we describe heterologous biosynthesis of cotylenol by engineering the biosynthetic pathway of brassicicenes in Aspergillus oryzae and harnessing the promiscuity of a cytochrome P450 from the biosynthesis of fusicoccin A (Figure 1b). A key intermediate, brassicicenes I (5), was further used to achieve the collective synthesis of alterbrassicicene E (6), brassicicenes A (7) and R (8).
Results and Discussion
Fusicoccanes feature a characteristic dicyclopenta[a,d]cyclooctane (5-8-5) ring system that is biosynthesized from geranylgeranyl pyrophosphate (GGPP) via class I terpene cyclization (Figure 2a). To date, two fusicoccadiene synthases have been identified by the analysis of the brassicicene biosynthesis-related gene cluster (BGC) in Alternaria brassicicola and Pseudocercospora fijiensis [34–35]. The 5-8-5 tricyclic scaffold is transformed into various fusicoccane natural products catalyzed by P450s, dioxygenases, dehydrogenases, and reductases. Therefore, we propose to harness the biosynthetic pathway for brassicicenes, which share the same carbon skeleton and similar oxidation and unsaturation states as cotylenol and cotylenin A [36]. In a previous study, Oikawa and co-workers reported the identification of brassicicene BGC in Pseudocercospora fijiensis [37]. By heterologous expression of this BGC in Aspergillus oryzae, brassicicene I was produced by the transformant AO-bscABCDE at a titer of 5.5 mg/L. Recently, we identified a new BGC for brassicicenes, namely, abn, from the brassicicene-producing strain A. brassicicola XXC (Figure 2b) [38]. We constructed an A. oryzae strain with the homologous gene abnABCDE. As expected, compound 5 was produced at a titer of 8 mg/L (Figure 2c). By co-fermenting with Amberlite XAD-16, an enhanced yield of 30 mg/L was achieved, thus allowing further transformation into other natural products.
Heterologous production of brassicicene I in an engineered A. oryzae strain. (a) Biosynthesis of fusicoccin A in Phomopsis (Fusicoccum) amygdali. (b) Brassicicene BGC in A. brassicicola XXC. (c) Heterologous production of brassicicene I (5) in an engineered AO strain.
We next carried out the formal synthesis of cotylenin A and cotylenol (Figure 3a). Oxidation of brassicicene I with Dess–Martin reagent afforded intermediate 9 in 92% yield. The tertiary hydroxy group of compound 9 was further protected with a TMS group to provide compound 10 in 90% yield, a key intermediate in the synthesis of cotylenol and cotylenin A by Nakada and co-workers [21]. However, installing the C9 hydroxy group requires the use of stoichiometric MoOPH [39], which raises toxicity and safety issues. Therefore, we sought an enzymatic method to selectively oxidize 5 at the C9 position. Dairi and co-workers reported that Orf7 oxidizes compound 11 at the C9 position in the biosynthesis of fusicoccin A (Figure 3b) [40]. Given the structural similarities between compound 5 and compound 11, we hypothesized that Orf7 might also catalyze the hydroxylation of compound 5 at C9. Hence, we fed an A. oryzae strain that expressed the orf7 gene with compound 5. To our delight, compound 3 was obtained successfully (Figure 3c). To stably produce 3 by fermentation, we constructed an A. oryzae strain that integrates abnABCDE with orf7, achieving a yield of 60 mg/kg rice through rice fermentation.
Synthesis of cotylenol (3). (a) Synthesis of Nakada’s intermediate 10 from 5. (b) Orf7 catalyzes the oxidation of 11 in the biosynthesis of fusicoccin A (2). (c) LC–MS analysis of the production of 3 through AO-abnABCDE+orf7 heterologous expression or AO-orf7 biotransformation.
We next targeted alterbrassicicene E (6), brassicicenes A (7) and R (8) (Scheme 1). The secondary hydroxy group of brassicicene I was selectively TBS-protected in the presence of TBSOTf and 2,6-lutidine to give compound 13 in 93% yield. Then, compound 13 underwent oxidative rearrangement with PCC to afford ketone 14 in 61% yield. Under Luche reduction conditions, compound 15 and its diastereomer were obtained in a total yield of 90% at a ratio of 1:0.7. To improve the diastereoselectivity, we examined other reduction conditions and found that ʟ-Selectride afforded compound 15 in 90% yield with a dr of 9:1. Upon desilylation with TBAF, compound 15 was converted into alterbrassicicene E (6) in 80% yield. To synthesize brassicicenes A (7) and R (8), the tertiary hydroxy group of compound 13 was protected with a TES group to furnish compound 16 in 89% yield. By screening several conditions, we found that allylic oxidation of compound 16 could be achieved in the presence of chromium trioxide–3,5-dimethylpyrazole complex [41] to provide compound 17 in 76% yield. After deprotection of the TBS and TES groups with TBAF, brassicicene A (7) was obtained in 75% yield. Compound 17 was subjected to α-hydroxylation from the less-hindered convex face using Davis’s oxaziridine [25], furnishing intermediate 18 in 72% yield. After deprotection of the TBS and TES groups, brassicicene R (8) was obtained in 70% yield. Therefore, alterbrassicicene E (6) and brassicicenes A (7) and R (8) were synthesized from brassicicene I over 4 or 5 chemical steps.
Synthesis of alterbrassicicene E (6) and brassicicenes A (7) and R (8) from brassicicene I (5).
Conclusion
In summary, the diverse biological activities and complex structures of fusicoccane diterpenoids have stimulated multiple elegant chemical syntheses. In contrast to these approaches, we harnessed the biosynthetic machinery of brassicicenes to produce brassicicene I in an engineered A. oryzae strain. Brassicicene I was further oxidized by a cytochrome P450 from the biosynthesis of fusicoccin A, thus leading to total biosynthesis of cotylenol in A. oryzae. Three fusicoccane diterpenoids, including alterbrassicicene E and brassicicenes A and R, were efficiently synthesized from brassicicene I in 4 or 5 chemical steps. This work lays the foundation for the preparation of fusicoccane natural products and exploration of their biological activities.
Supporting Information
File 1Experimental data and copies of spectra.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1de Boer A H de Vries-van Leeuwen I J Trends Plant Sci 20121736036810.1016/j.tplants.2012.02.00722465041 · doi ↗ · pubmed ↗
- 2Ballio A Chain E B De Leo P Erlanger B F Mauri M Tonolo A Nature 196420329710.1038/203297 a 0 · doi ↗
- 3Sassa T Agric Biol Chem 1971351415141810.1271/bbb 1961.35.1415 · doi ↗
- 4Sassa T Ooi T Nukina M Ikeda M Kato N Biosci, Biotechnol, Biochem 1998621815181810.1271/bbb.62.181527392691 · doi ↗ · pubmed ↗
- 5Tang Y Xue Y Du G Wang J Liu J Sun B Li X-N Yao G Luo Z Zhang Y Angew Chem, Int Ed 201655124069407310.1002/anie.20160031326916098 · doi ↗ · pubmed ↗
- 6Li F Lin S Zhang S Pan L Chai C Su J-C Yang B Liu J Wang J Hu Z J Nat Prod 20208361931193810.1021/acs.jnatprod.0c 0016532520548 · doi ↗ · pubmed ↗
- 7Zhou P Zhang X Dai C Yan S Wei M Feng W Li Q Liu J Zhu H Hu Z J Org Chem 2022877333734110.1021/acs.joc.2c 0052835588185 · doi ↗ · pubmed ↗
- 8Ohkanda J Chem Lett 202150576710.1246/cl.200670 · doi ↗