Evaluation of Pupal Parasitoids Trichomalopsis ovigastra and Pachycrepoideus vindemiae as Potential Biological Control Agents of Bactrocera dorsalis
Ziwen Teng, Yiting Wang, Minghao Jiang, Yikun Zhang, Xintong Wang, Fanghao Wan, Hongxu Zhou

TL;DR
Researchers found that the parasitoid wasp Trichomalopsis ovigastra is a strong candidate for controlling the invasive fruit fly Bactrocera dorsalis in northern China.
Contribution
The study reveals T. ovigastra's superior parasitism of B. dorsalis and environmental resilience compared to P. vindemiae, suggesting its potential as a biocontrol agent.
Findings
T. ovigastra showed nearly 50% parasitism of B. dorsalis, compared to less than 0.3% by P. vindemiae.
T. ovigastra had shorter developmental times and greater tolerance to environmental stressors like temperature extremes and desiccation.
B. dorsalis has invaded northern China, but no native parasitoids have been reported there, making T. ovigastra a promising local biocontrol candidate.
Abstract
Parasitoid wasps are well-known biocontrol agents, and discovering new species and examining their biological traits are essential to fully exploring their potential for pest management. In northern China, we collected Trichomalopsis ovigastra Sureshan & Narendran (Hymenoptera: Pteromalidae) directly from the field. This parasitoid species had previously only been described morphologically. In this study, we investigated additional biological characteristics of T. ovigastra and compared them with those of Pachycrepoideus vindemiae (Rondani) (Hymenoptera: Pteromalidae), a well-known parasitoid wasp of Diptera insects. Our results indicate that T. ovigastra exhibits stronger parasitism of both Drosophila melanogaster Meigen (Diptera: Drosophilidae) and Bactrocera dorsalis (Hendel) (Diptera: Tephritidae), shorter developmental times, and higher resistance to environmental stressors. Given…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
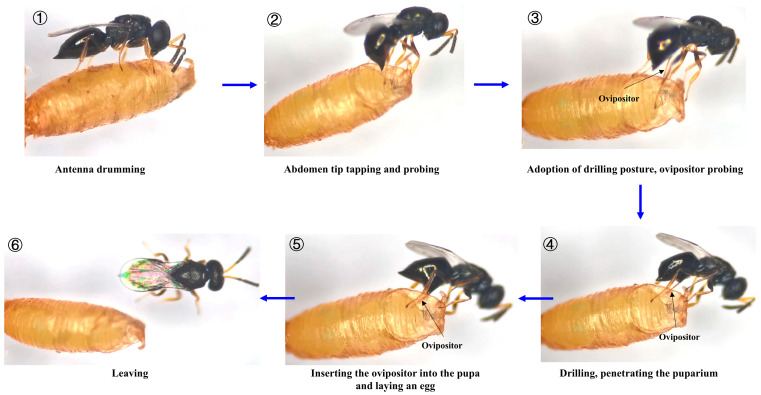 Figure 1
Figure 1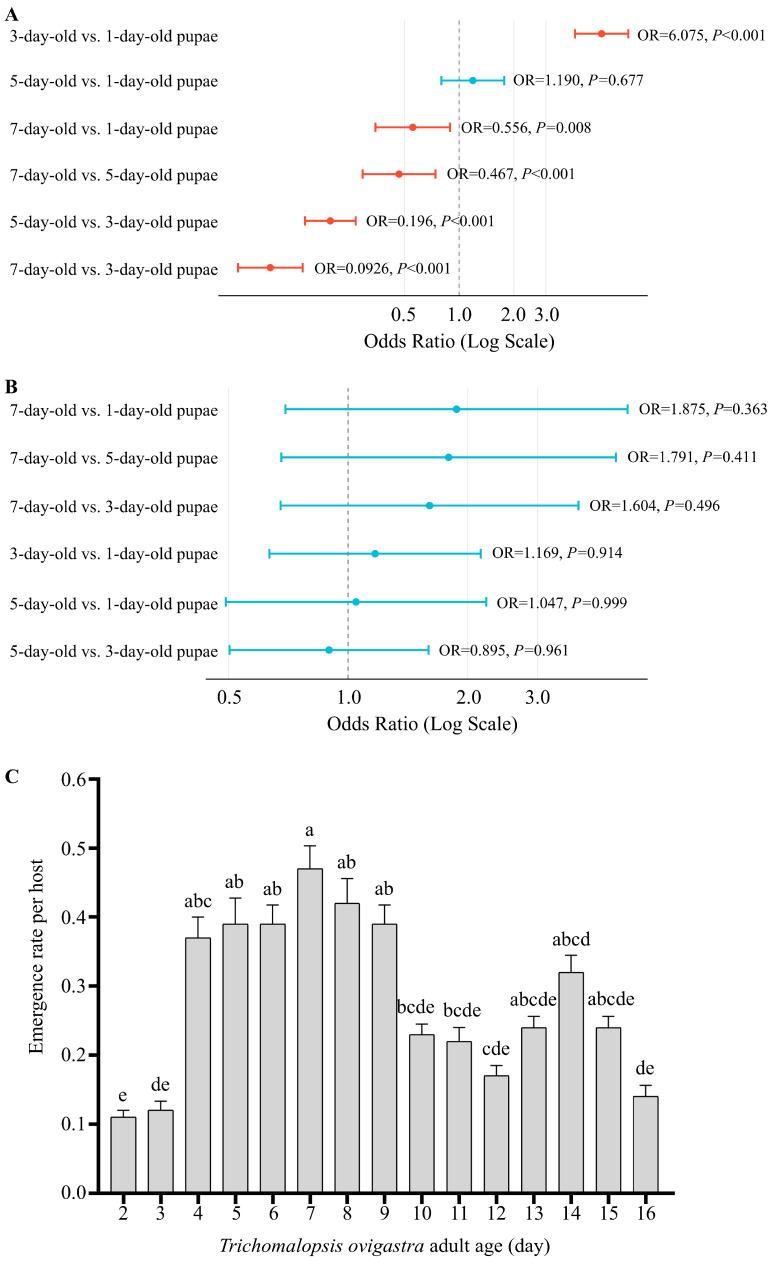 Figure 2
Figure 2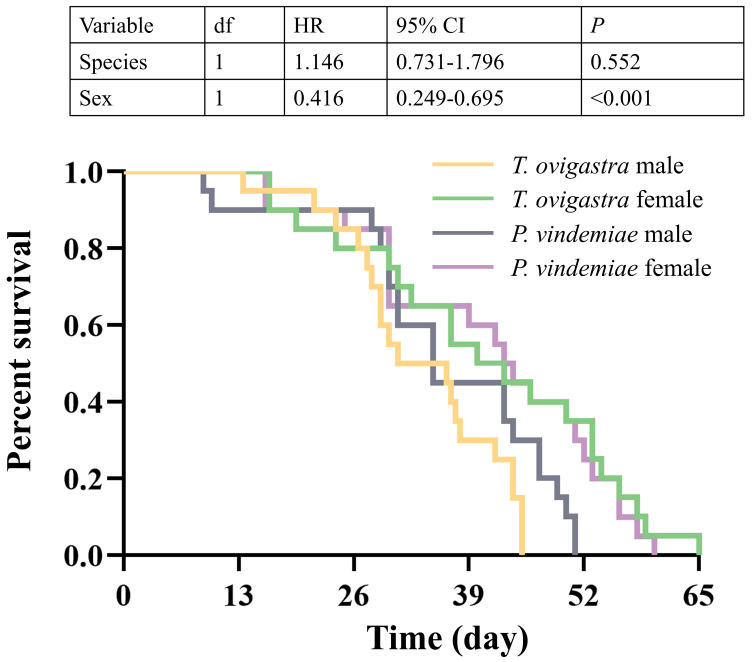 Figure 3
Figure 3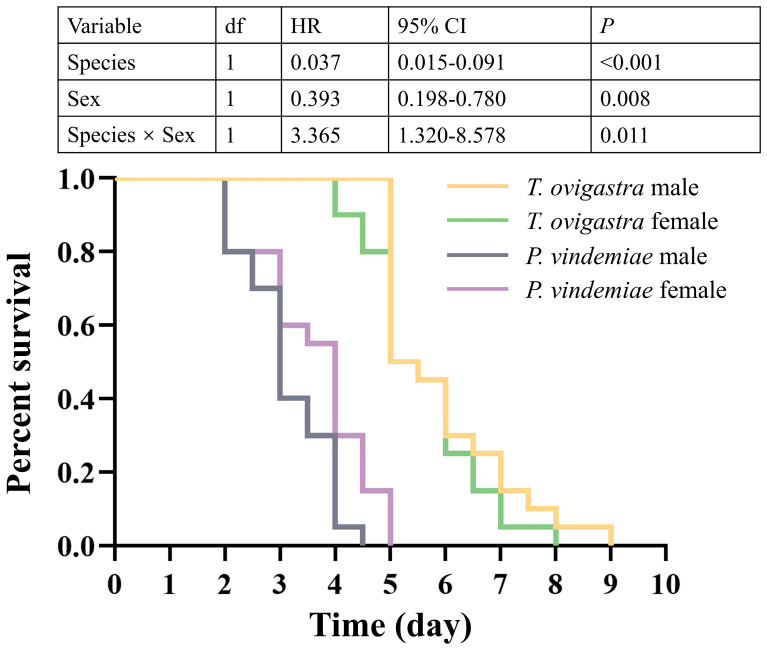 Figure 4
Figure 4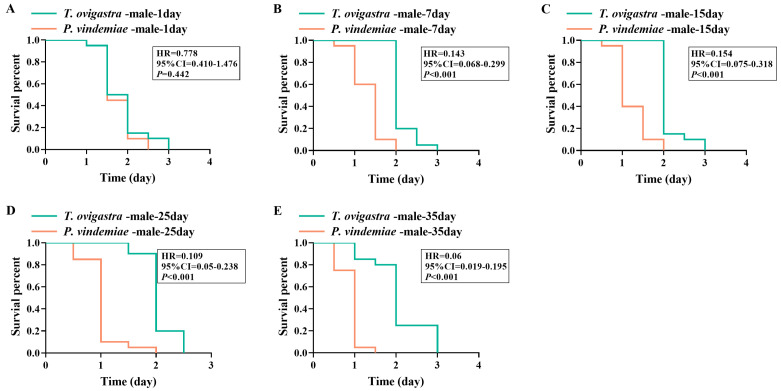 Figure 5
Figure 5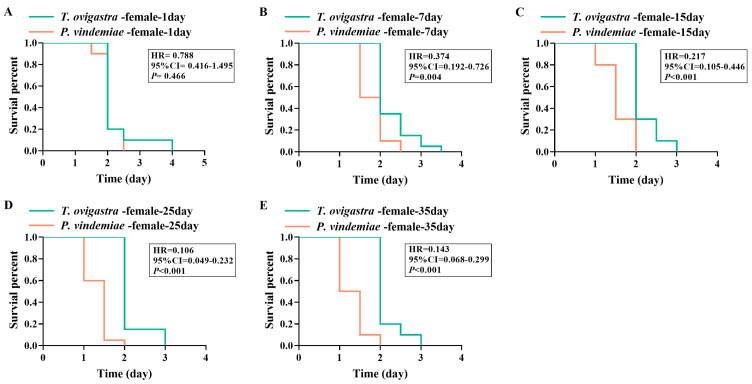 Figure 6
Figure 6- —the Qingdao Natural Science Foundation
- —Qingdao Science and Technology Benefiting the People Demonstration Project
- —Major Scientific and Technological Innovation Projects in Shandong Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect behavior and control techniques · Insect-Plant Interactions and Control · Biological Control of Invasive Species
1. Introduction
The oriental fruit fly Bactrocera dorsalis (Hendel) (Diptera: Tephritidae) is a highly invasive pest species. Under natural field conditions, it has been recorded to have attacked 481 host taxa across 212 genera in 79 families, most notably Moraceae (38 taxa), Rutaceae (37 taxa), Solanaceae (33 taxa), and Cucurbitaceae (29 taxa) [1]. In China, B. dorsalis poses a severe threat to fruit production [2]. For example, in southern China, infestation rates on oranges reached 80–90% [3], and the national economic impacts on the citrus industry have been projected at approximately USD 40 billion [2]. Since its initial detection in Taiwan in 1912, B. dorsalis has steadily spread northward across mainland China [3]. Adults were first recorded on Hainan Island in the 1930s and have since colonized tropical and subtropical zones throughout the country [4]. More recently, B. dorsalis has been reported in temperate northern provinces such as Beijing, Hebei, and Henan [3,4,5]. Northern China, a major fruit-growing region producing peaches, apples, jujubes, and pears [6], now faces mounting risk; in Zhoukoudian (Beijing), plum infestation rates by B. dorsalis reached 10% [3]. To curb its further spread, China has implemented strict quarantine measures—such as trapping surveillance and postharvest thermal treatments—to detect and eliminate B. dorsalis infestations [7,8]. Although mass trapping (pheromone- and bait-based lures) [9], biological control (parasitoids, baculoviruses, and entomopathogenic fungi) [10,11,12], and the sterile insect technique [13] have been trialed, none have been widely adopted. Currently, synthetic insecticides remain the primary management tool, but their overuse has caused high-level resistance in B. dorsalis and has led to more severe outbreaks [14].
Parasitoid wasps play a pivotal role in structuring both natural and managed ecosystems [15] and hold substantial economic value as biocontrol agents against crop pests [16]. To harness their potential, comprehensive evaluations of candidate species’ traits—such as host range [17], parasitism efficacy [18], development and growth [19], stress tolerance [20], and host–parasitoid interactions [21]—are essential, because these parameters critically affect field performance [20,22,23,24].
Recent surveys of fruit fly parasitoids in orchards in northern China showed the occurrence of some resident parasitoid species attacking B. dorsalis. Among these species, Trichomalopsis ovigastra Sureshan & Narendran (Hymenoptera: Pteromalidae) and Pachycrepoideus vindemiae (Rondani) (Hymenoptera: Pteromalidae) were identified. T. ovigastra was first described from specimens collected in Kerala, India [25], but beyond that original record, its distribution and biology remain almost entirely undocumented. Our preliminary laboratory observations indicate that it can parasitize pupae of Drosophila melanogaster Meigen (Diptera: Drosophilidae) and of B. dorsalis. Although direct studies of T. ovigastra are scarce, research on other species within the genus Trichomalopsis may offer valuable insights. For example, Floate et al. have extensively studied T. sarcophagae (Gahan) for its host-searching behavior at different soil depths [26], comparative production on fresh versus freeze-killed hosts [27], cold tolerance [28], and biocontrol performance in cattle feedlots [29]. Other work on Trichomalopsis includes species descriptions [30], morphological and developmental analyses [31,32], parasitoid population surveys [33], host associations [34], and reproductive capacity assays [35]. Importantly, some Trichomalopsis—such as T. apanteloctena (Crawford)—act as facultative hyperparasitoids on hosts such as Cotesia kariyai (Watanabe) (Hymenoptera: Braconidae) [36] and Apanteles sp. (Hymenoptera, Braconidae) [37]. In contrast, P. vindemiae has been more extensively studied. P. vindemiae is a solitary, generalist pupal parasitoid first described in Italy [38] and now distributed in over 60 countries [39]. It has been reported to parasitize more than 60 fly species, including economically important species in the Tephritidae and Drosophilidae families [39,40,41,42]. Recent studies have focused on its morphology [43], parasitic behavior [42], field release [40], and the molecular mechanisms underlying host regulation [44]. It has shown significant potential as a biological control agent against dipteran pests [45] and has been evaluated primarily for controlling stable and house flies [46,47,48]. P. vindemiae can also act as a hyperparasitoid, necessitating caution when deploying it in systems involving other parasitoids [39].
D. melanogaster, which feeds on fermenting fruits, does not pose a significant agricultural threat. However, it serves as an excellent model organism in life sciences research [49]. Consequently, extensive research has been conducted on the mechanisms by which parasitoids regulate D. melanogaster, offering potential insights for applying parasitoids in biological control [50]. Using D. melanogaster as a host, our laboratory has previously studied the circadian activity, identified clock genes of P. vindemiae [42], and developed starvation-resistant lines via multi-generation artificial selection, assessing biological traits, such as starvation resistance, in both selected and non-selected lines [22]. In this study, we selected D. melanogaster as a secondary host model to compare the performance of T. ovigastra with that of the well-established ectoparasitoid P. vindemiae. Moreover, given the recent northward expansion of B. dorsalis into regions lacking native parasitoids, there is an urgent need to identify effective local natural enemies. Based on preliminary observations in our laboratory, we hypothesize that T. ovigastra may exhibit stronger parasitic ability against B. dorsalis than P. vindemiae, though experimental verification is still lacking. Therefore, this study had two objectives: (1) to compare T. ovigastra and P. vindemiae in key biocontrol traits—offspring production, sex ratio, developmental time, adult longevity, and resistance to temperature extremes, starvation, and desiccation—using D. melanogaster as a model host; (2) to assess T. ovigastra’s parasitism performance for B. dorsalis pupae, thereby evaluating its potential as a native biocontrol agent against this invasive pest.
2. Materials and Methods
2.1. Insect Rearing
The D. melanogaster w^1118^ strain (Bloomington Drosophila Stock Center, Indiana University, Bloomington, IN, USA) was maintained on standard cornmeal-yeast medium. Colonies of P. vindemiae and T. ovigastra were established by parasitizing D. melanogaster pupae. B. dorsalis were sourced from South China Agricultural University and kept in an artificial climate room following the methods described by Zhu et al. [3]. We regularly monitored the survival rate, fecundity, and hatchability to prevent population degradation. Adults were housed in transparent culture tubes (2.5 × 10 cm) with 20% (v/v) sucrose solution. All insects were reared under controlled conditions at 25 °C, with 60 ± 5% relative humidity (RH) and a 12:12 L:D photoperiod.
2.2. Oviposition Behavior of T. ovigastra
Given the limited knowledge on T. ovigastra beyond morphological identification, preliminary observations of its oviposition behavior were conducted to confirm its parasitism of host pupae. A one-day-old, fully mated female wasp with no prior parasitism experience was individually introduced into a transparent Petri dish (8.5 cm diameter × 1.8 cm depth) containing a single two-day-old D. melanogaster pupa. The behavior was recorded under a high-resolution stereomicroscope (SangNond, Shenzhen, China) at 25 °C.
2.3. Offspring Number and Sex Ratio
In each culture tube (2.5 cm in diameter and 10 cm in height), 20 specific-age D. melanogaster pupae or 10 B. dorsalis pupae were introduced, along with a two-day-old, fully mated female wasp with no prior oviposition experience. The pupae ages for D. melanogaster were 0, 1, 2, and 3 days, with 0-day-old referring to pupae within 24 h of pupariation, 1-day-old referring to pupae that had been pupating for 1 day, and so on. For B. dorsalis, the pupae ages were 1, 3, 5, and 7 days. Following an initial 24 h parasitism period, fresh hosts were introduced daily. For D. melanogaster, we tested ten individual female wasps of each species and monitored their parasitism daily for 19 consecutive days (until each wasp reached 20 days of age). For B. dorsalis, we likewise tested ten individual female T. ovigastra and assessed parasitism over a 15-day period (until they reached 16 days of age). After the wasp offspring emerged, the number of males and females was recorded to calculate offspring production and sex ratio. In the initial experiments, we observed that P. vindemiae had a parasitism rate of less than 0.3% on B. dorsalis pupae, which was deemed inefficient and impractical for further analysis. As a result, P. vindemiae was excluded from the trials involving B. dorsalis.
2.4. Developmental Duration
The timing of oviposition was recorded, and adult emergence was monitored at 12 h intervals. The developmental time from egg to adult emergence was calculated separately for males and females. Twenty individuals per sex from each parasitoid species were assessed.
2.5. Longevity
Newly emerged, unmated male and female parasitoids were separated by sex, and each individual was placed into a separate tube. A 20% sucrose solution (Sangon Biotech, Shanghai, China) was provided as the nutritional source. Mortality was recorded every 12 h until all individuals had died. The assays were conducted at 25 °C and 60 ± 5% RH with a 12:12 h light:dark cycle.
2.6. High and Low Temperature Resistance
To assess thermal tolerance, unmated adults of defined ages (1, 15, and 30 days) from each parasitoid species were exposed individually to high (45 °C for 30 min) or low temperature (−10 °C for 2.5 h) conditions. Each sex was tested using 20 individuals. After thermal stress, the insects were returned to 25 °C, 60 ± 5% RH, and 12:12 h light:dark conditions. Survival was assessed 24 h post-treatment.
2.7. Starvation Resistance
Twenty newly emerged, unmated parasitoids from each species were individually maintained with access to water only. Mortality was recorded at 12 h intervals until all individuals died. The test conditions were 25 °C, 60 ± 5% RH, and a 12:12 h light:dark cycle.
2.8. Desiccation Resistance
Unmated wasps of both sexes and selected ages (1, 7, 15, 25, and 35 days) were tested individually. Twenty individuals of each sex per species were placed in separate tubes with 50 silica gel beads (3–5 mm) to induce desiccation. A sponge plug (1.2 cm height × 2.4 cm diameter) was placed above the beads to prevent direct contact with the insects. The tubes were sealed with Parafilm^®^ to reduce the relative humidity below 12% within 15 min. The wasps were held at 25 °C with a 12:12 h light:dark cycle, and mortality was recorded every 12 h.
2.9. Statistical Analysis
All statistical analyses were performed using R software (version 4.4.3) [51]. Offspring number and sex ratio were analyzed using generalized linear mixed models (GLMMs) with binomial distribution, considering parent ID as a random effect and parasitoid species as a fixed effect. The glmer function in the lme4 package (version 1.1.36) was used [52]. Visualizations were generated using ggplot2 (version 3.5.1). Cox regressions, implemented using the coxph function from the survival package (version 3.8.3) [53], were used to analyze developmental duration, lifespan, and survival under starvation and desiccation. To identify optimal covariates (species, sex, and interaction), models were compared based on Akaike Information Criterion (AIC), Bayesian Information Criterion (BIC), and likelihood ratio tests. The best-fitting model was selected using the lowest AIC/BIC and significant likelihood improvements. Survival curves were plotted with the ggsurvplot function from survminer (version 0.5.0) [54]. This study presents only the results of the best-fit model. Binary logistic regression using the glm function from stats (version 4.4.2) [51] was applied to analyze thermal resistance data. Model selection followed the same approach as above, evaluating AIC, BIC, and likelihood ratio tests to determine the most appropriate predictors (species, sex, and interaction).
3. Results
3.1. Oviposition Behavior of T. ovigastra
The oviposition process of T. ovigastra, from antenna drumming to post-oviposition departure, was divided into six sequential phases: ① Antenna drumming: upon locating a host pupa, the female examined its surface by tapping it with her antennae. ② Abdomen tip tapping and probing: after antennal examination, the female curved her abdomen, gently probing the puparium surface with the tip. ③ Adoption of drilling posture and ovipositor probing: maintaining a curved posture, the female extended her ovipositor in preparation for penetration. ④ Drilling and penetrating the puparium: gradual rotational movements of the ovipositor characterized the initial drilling, transitioning to forceful thrusting or rocking motions to complete penetration. ⑤ Inserting the ovipositor into the pupa and laying an egg: the ovipositor was fully inserted into the host, where it was maneuvered internally for egg deposition. After laying the egg, the ovipositor was withdrawn. ⑥ Leaving: following oviposition, the female briefly touched the puparium with her antennae while walking over it, then moved away from the host (Figure 1).
3.2. Offspring Number and Sex Ratio
When parasitizing D. melanogaster pupae of different ages, T. ovigastra consistently produced more offspring than P. vindemiae. Specifically, each T. ovigastra female yielded mean offspring counts of 148.4, 234.8, 224.9, and 151.3 from 0-, 1-, 2-, and 3-day-old pupae, respectively, whereas each P. vindemiae female produced 79.1, 134.9, 138.9, and 84.7 offspring under the same conditions. Despite these differences in fecundity, the offspring sex ratio remained constant at 0.6 for both species across all pupal ages. For each parasitoid, the number of emerged offspring and sex ratios did not differ significantly across host pupal ages (Table 1).
To further evaluate the parasitism of the two species over time, we applied GLMMs with a binomial distribution, comparing offspring emergence across adult wasp ages. The results showed that from days 6, 8–11, and 13–20, T. ovigastra produced significantly more offspring than P. vindemiae, while no differences were found at other ages (Table 2 and Table S1).
In the preliminary experiments, we found that P. vindemiae exhibited a parasitism rate below 0.3% on B. dorsalis pupae, making it unsuitable for subsequent analyses due to extremely low efficiency and lack of practical value. Consequently, we excluded P. vindemiae from the trials involving B. dorsalis. When T. ovigastra parasitized B. dorsalis pupae of different ages, significant differences in offspring emergence were observed. Except for the comparison between 5-day-old and 1-day-old pupae, where no significant differences were observed, all other age comparisons revealed significant differences. The highest emergence rate was recorded in 3-day-old pupae (Figure 2A and Table S2). However, the offspring sex ratio remained consistent across all pupal ages (Figure 2B and Table S2). In addition, parasitism by T. ovigastra differed significantly among individuals aged 2 to 16 days, with those at the younger and older ends of this age range exhibiting reduced parasitism efficiency (Figure 2C).
3.3. Developmental Duration and Longevity
T. ovigastra completed development from egg to adult emergence significantly faster than P. vindemiae. The females of both species exhibited prolonged developmental durations compared to the males (Table 3). No significant interspecific differences in adult longevity were detected between T. ovigastra and P. vindemiae (Figure 3).
3.4. High and Low Temperature Resistance
In the low-temperature resistance trials, no significant difference was found between the two parasitoids at day 1 post-emergence. However, at 15 and 30 days of age, T. ovigastra displayed significantly greater survival than P. vindemiae. In contrast, T. ovigastra consistently outperformed P. vindemiae at all the tested ages (1, 15, and 30 days) under high-temperature stress (Table 4).
3.5. Starvation Resistance
T. ovigastra exhibited significantly higher resistance to starvation than P. vindemiae (Figure 4). Moreover, no sex-related differences in starvation tolerance were observed in T. ovigastra, while in P. vindemiae, the females survived significantly longer than the males (Figure 4 and Table S3).
3.6. Desiccation Resistance
We assessed the desiccation tolerance across sexes and ages for both parasitoid species. On day 1, there was no difference in desiccation resistance between the two species’ male wasps (Figure 5A), nor between their females (Figure 6A). However, at all other tested ages, T. ovigastra exhibited significantly higher resistance than P. vindemiae, regardless of sex (Figure 5B–E and Figure 6B–E).
4. Discussion
Biological control constitutes a vital component of invasive pest management [55]. One strategy introduces and establishes natural enemies from the pest’s native range [56,57], but stringent regulations on importing non-native species often hinder its implementation [58]. An alternative and more straightforward approach enhances populations of resident natural enemies in newly invaded regions [58]. However, using parasitoids as an example, this strategy faces challenges as local parasitoids need varying amounts of time to adapt to new hosts, depending on their level of plasticity [59,60]. In our study, while T. ovigastra exhibited significantly higher parasitism than P. vindemiae on both hosts, its maximum emergence rate from B. dorsalis pupae remained below 50%. Such suboptimal parasitism likely reflects multiple ecological and physiological constraints, including incomplete host adaptation. Future studies should continue to measure T. ovigastra’s field parasitism rate on B. dorsalis to evaluate its host adaptation and pinpoint the factors limiting its efficacy.
Measuring offspring production over a limited interval provides an incomplete picture of a parasitoid’s reproductive potential compared to lifetime fecundity assessments [58]. For example, P. vindemiae maintains egg production throughout its adult life via both initial egg loads and ongoing oogenesis, whereas some species, such as Trichopria drosophilae Perkins (Hymenoptera: Diapriidae), cease oviposition midway through their lifespan due to arrested egg maturation [58,61]. Stacconi et al. (2017) suggested that these contrasting reproductive strategies may differentially influence the realized fecundity under field conditions, especially when the host availability fluctuates, potentially rendering T. drosophilae egg-limited in high-host environments [58]. Such insights could guide the selection of parasitoid species based on target pest densities. In this study, we counted offspring produced by parasitoids during the first 19 days on D. melanogaster and 15 days on B. dorsalis. Although T. ovigastra outperformed P. vindemiae between days 2 and 20, further studies are needed to determine the full duration and pattern of its oviposition period.
The number of offspring a parasitoid produces is strongly correlated with female longevity and, by extension, with adult nutritional intake [62,63,64]. Studies have demonstrated that Ascogaster quadridentata (Wesmael) (Hymenoptera: Braconidae), when provided with specific sugar or floral resources, experienced an extended lifespan. Moreover, the total number of parasitized host larvae per female was significantly correlated with the observed lifespan [62]. Hosts themselves can serve as important nutritional sources: host-feeding by Eupelmus vuilletti (Crawford) (Hymenoptera: Eupelmidae) boosts both egg production and survival [65], and female P. vindemiae that engage in host-feeding live significantly longer than those deprived of hosts [58]. It is hypothesized that females deprived of hosts may experience nutrient deficiencies, as they are unable to access the essential nutrients typically obtained through host-feeding [58]. We also observed host-feeding in T. ovigastra, but our longevity assays used only 20% sucrose solution. Future research should quantify survival when hosts are available.
The survival ability of parasitoids under unfavorable environmental conditions is also critical for their field biocontrol efficacy. For instance, releasing parasitoids at low pest densities is essential to prevent rapid growth of the target pest population, as delayed releases may hinder effective control. However, the absence of sufficient hosts can result in parasitoid extinction due to starvation. Thus, the starvation resistance of parasitoids is crucial for their survival and their ability to locate and parasitize hosts [66]. In this study, the laboratory data suggest that T. ovigastra may tolerate a wider range of temperatures and lower humidity levels compared to P. vindemiae. Consequently, these findings indicate that T. ovigastra may be more suitable for controlling B. dorsalis.
In conclusion, although this study provides valuable data on the parasitism, development, and stress tolerance of T. ovigastra, demonstrating its potential for biological control, several critical knowledge gaps remain. For instance, whether T. ovigastra exhibits facultative hyperparasitism—similar to P. vindemiae—is an urgent area for investigation. Notably, other species in the same genus, such as T. apanteloctena, are known to be idiobiont ecto-hyperparasitoids of prepupae or pupae of various Cotesia species [67]. Historical biocontrol programs reveal that hyperparasitism can undermine parasitoid efficacy [68]; for instance, C. rubecula (Marshall) experienced hyperparasitism rates rising from 37.9% in 1987 to 100% in August 1988, correlating with a failure to establish the agent in subsequent seasons [69]. Moreover, facultative hyperparasitoids that preferentially attack primary parasitoids over pest populations can be detrimental to control outcomes [70,71]. Therefore, determining whether T. ovigastra exhibits hyperparasitism and quantifying its host-selection preferences are essential steps. Additionally, research into its host-range breadth, diapause regulation, scalable rearing protocols, optimized release strategies, and molecular mechanisms of host manipulation will be indispensable for the safe and effective application of T. ovigastra in biocontrol programs.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Liquido N.J. Mcquate G.T. Birnbaum A.L. Hanlin M.A. Nakamichi K.A. Inskeep J.R. Ching A.J. Marnell S.A. Kurashima R.S. A Review of Recorded Host Plants of Oriental Fruit Fly, Bactrocera (Bactrocera) dorsalis (Hendel) (Diptera: Tephritidae), Version 3.0. USDA CPHST Online Database 2017 Available online: https://coffhi.cphst.org/(accessed on 23 June 2025)
- 2Zhao Z. Carey J.R. Li Z. The global epidemic of Bactrocera pests: Mixed-species invasions and risk assessment Annu. Rev. Entomol.2023113410.1146/annurev-ento-012723-10265837708416 · doi ↗ · pubmed ↗
- 3Zhu Y.F. Tan X.M. Qi F.J. Teng Z.W. Fan Y.J. Shang M.Q. Lu Z.Z. Wan F.H. Zhou H.X. The host shift of Bactrocera dorsalis: Early warning of the risk of damage to the fruit industry in northern China Entomol. Gen.20224269169910.1127/entomologia/2022/1453 · doi ↗
- 4Zeng Y. Reddy G.V.P. Li Z. Qin Y. Wang Y. Pan X. Jiang F. Gao F. Zhao Z.H. Global distribution and invasion pattern of oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae)J. Appl. Entomol.201914316517610.1111/jen.12582 · doi ↗
- 5Liang L. Li Y. Risk Analysis of Bactrocera dorsalis (Hendel) in Shaanxi Province Shaanxi For. Sci. Technol.201724648 Available online: https://www.cnki.net/(accessed on 23 June 2025)(In Chinese)
- 6Zhu Y.F. Qi F.J. Tan X.M. Zhang T. Teng Z.W. Fan Y.J. Wan F.H. Zhou H.X. Use of age-stage, two-sex life table to compare the fitness of Bactrocera dorsalis (Diptera: Tephritidae) on northern and southern host fruits in China Insects 20221325810.3390/insects 1303025835323555 PMC 8954565 · doi ↗ · pubmed ↗
- 7Jaffar S. Rizvi S.A.H. Lu Y. Understanding the invasion, ecological adaptations, and management strategies of Bactrocera dorsalis in China: A review Horticulturae 20239100410.3390/horticulturae 9091004 · doi ↗
- 8Fang Y. Kang F. Zhan G. Ma C. Li Y. Wang L. Wei Y. Gao X. Li Z. Wang Y. The effects of a cold disinfestation on Bactrocera dorsalis survival and navel orange quality Insects 20191045210.3390/insects 1012045231847197 PMC 6955761 · doi ↗ · pubmed ↗