Genome-Wide Identification and Analysis of bZIP Transcription Factor Gene Family in Broomcorn Millet (Panicum miliaceum L.)
Peipei An, Tianxiang Liu, Zhijie Shui, Panrong Ren, Shan Duan

TL;DR
This study identifies and analyzes 144 bZIP transcription factor genes in broomcorn millet, revealing their roles in stress response and seed germination.
Contribution
The first genome-wide analysis of the bZIP gene family in broomcorn millet, including expression patterns under salt stress.
Findings
144 bZIP transcription factors were identified and grouped into eleven subfamilies.
18 PmbZIP genes showed differential expression during seed germination under salt stress.
Promoter analysis revealed hormone and stress responsiveness in PmbZIP genes.
Abstract
Background: Basic (region) leucine zippers (bZIPs) make up one of the largest families and are some of the most prevalent evolutionarily conserved transcription factors (TFs) in eukaryotic organisms. Plant bZIP family members are involved in seed germination, vegetative growth, flower development, light response, and various biotic/abiotic stress response pathways. Nevertheless, a detailed identification and genome-wide analysis of the bZIP family genes in broomcorn millet have not been conducted. Methods: In this research, we performed genome-wide identification, phylogenetic analysis, cis-elements analysis, and expression pattern analysis. Results: 144 bZIP transcription factors were identified from the P. miliaceum genome and classified into eleven subfamilies using a phylogenetic analysis. Motif and bZIP domain sequence alignment analyses indicated that the members in each subfamily…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
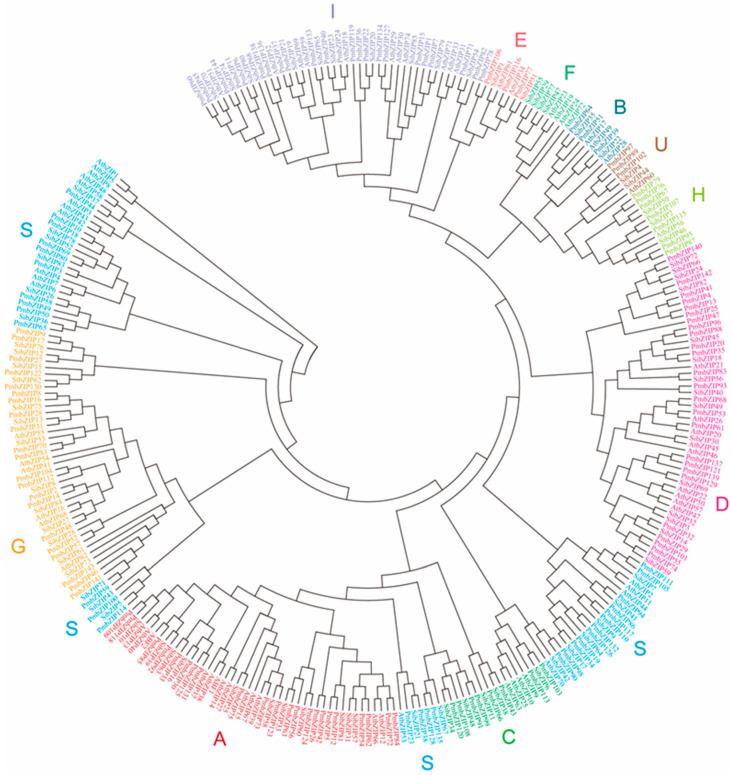 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4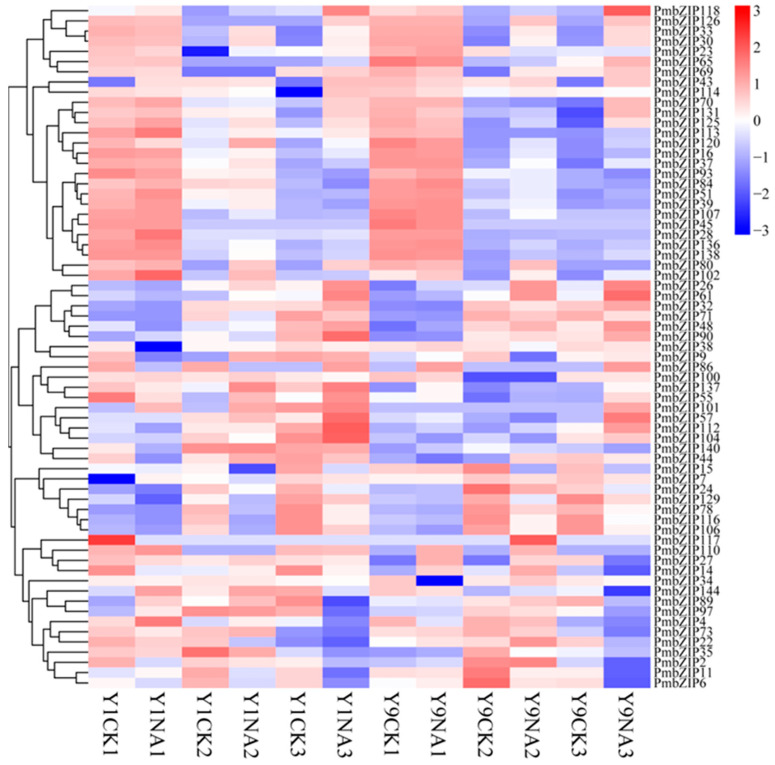 Figure 5
Figure 5- —Doctoral Research Fund of Longdong University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant Gene Expression Analysis · Plant Stress Responses and Tolerance
1. Introduction
Broomcorn millet (Panicum miliaceum L.) is one of the plants that was cultivated the earliest in the world and is also known as millet, common millet, or proso millet. It has been cultivated for more than 10,000 years and played significant roles in food security and cultural history in China [1]. Broomcorn millet has a short growing period and is the most water-efficient; has a high salt tolerance and good nutrient resource usage efficiency; and is high in proteins, certain minerals, and antioxidants compared to most other cereals [2,3,4]. Broomcorn millet is an allotetraploid with 36 chromosomes (2n = 4× = 36) [5], and its genome size is approximated to be ~923 Mb [6]. As a C4 photosynthesis crop that is closely related to the bioenergy crop switchgrass (Panicum virgatum L.), broomcorn millet has been reported to have the maximum water use efficiency (WUE), which may be due to its slow breathing rate, short generation time (about 60–90 days), and high harvest index [7,8,9]. Progress in molecular biology research on broomcorn millet is slow, and only a simple genetic map and a few genetic markers have been found. In 2019, the millet genome was sequenced, and phylogenetic analysis showed that millet divided into broomcorn millet and foxtail millet ~13.1 million years ago (Mya), while broomcorn millet became an allotetraploid ~5.91 million years ago [10]. One study also identified C4 candidate genes in broomcorn millet, and these genes were found to be spread over all three C4 subtypes, thus suggesting that broomcorn millet coexisted in three different carbon fixation pathways [6]. In the immediate future, broomcorn millet will become a crucial crop that can help in diversifying agriculture and promoting a healthier diet for humans. Therefore, searching for and identifying advantageous genes in millet will facilitate further basic scientific research on molecular breeding processes.
Transcription factors (TFs, also called trans-regulatory factors) are proteins with DNA-binding function, and they bind to specific cis-regulatory elements (cis-elements) and directly regulate the transcription of DNA to mRNA [11]. There are more divergent TF families and more unique DNA-binding domains (DBDs) in plant genomes [12], which also play a vital role in the regulation of growth, development, and environmental response. Structurally, TFs are usually classified by their DNA-binding domains: Basic (region) leucine zipper (bZIP) TFs have a basic region that binds DNA in the N-terminal and a leucine zipper dimerization motif in the C-terminal [13]. The basic DNA-binding region is an invariant N-X7-R/K motif, which has asparagine (N) and basic (R/K) residues with exact spacing; meanwhile, the ZIP domain within an alpha helix consists of heptad repeats of leucine (L) or a related hydrophobic amino acid [14]. Charged amino acids produce attractive and repulsive g-e’ pairs to regulate dimerization specificity and bind to DNA [15].
Among various transcription factor families in plants, basic leucine zipper (bZIP) proteins represent one of the most conserved and extensively studied groups. They are involved in diverse biological processes, particularly in mediating plant responses to abiotic stresses such as drought, salinity, and cold. bZIP transcription factors are well known for their central role in abscisic acid (ABA)-dependent signaling pathways, especially the PP2C–SnRK2–AREB cascade, which regulates key stress-adaptive responses and seed germination [16]. Compared to other transcription factor families (e.g., zinc finger and bHLH), bZIP genes have been more directly linked to core hormonal and environmental response mechanisms. To date, many plant bZIP transcription factor families have been identified and characterized extensively, including 78 AtbZIPs in Arabidopsis thaliana [13,14], 89 OsbZIPs in rice (Oryza sativa) [17], 187 TabZIPs in wheat (Triticum aestivum) [18], 85 SibZIPs in foxtail millet (Setaria italica), 103 OebZIPs in olive (Olea europaea) [19], 154 PhebZIPs in bamboo (Phyllostachys edulis) [20], 247 BnbZIPs in Brassica napus [21], 50 FvbZIPs in strawberry (Fragaria vesca) [22], 115 ZlbZIPs in Zizania latifolia [23], and 66 ItfbZIPs in sweet potato (Ipomoea trifida) [24]. Numerous studies have shown that different subgroups of bZIP TFs are involved in multiple regulation pathways in plants. bZIP TFs in Group A mainly take part in ABA signaling [25,26,27], abiotic stress responses [28,29,30,31], seed germination [32,33], and plant floral transition control [34,35,36]; bZIP TFs in Groups B and K are important regulators of endoplasmic reticulum (ER) stress response [14,37]; those in Groups C and S regulate sucrose signaling and seed storage protein production [38,39,40]; those in Group D are involved in detoxification processes and pathogen defense responses with salicylic acid (SA) defenses against biotrophic pathogens, as well as the defense of hormones jasmonic acid (JA) and ethylene (ET) against necrotrophies [14,41,42,43,44,45,46]; E-members and M-members might control pollen development [47]; F subfamily members control genes that encode for Zn transporters [14,48,49]; those in Groups G and J have been reported to regulate ER–Golgi transport and pathogen defense [50,51]; H-members are implicated in multiple hormone signaling pathways and development during light regulation [52,53,54]; bZIP TFs in Group I participate in osmosensory responses and root bending regulation [55,56]. bZIP target sequences often contain an ACGT core [57], such as G-box (CACGTG), C-box (GACGTC), A-box (TACGTA), etc. However, one investigator also found that some bZIPs can bind non-ACGT sequences [58,59].
Although the function of bZIP TFs has been reported and their identification and analysis have been conducted in Arabidopsis and many species, their roles in broomcorn millet remain largely unknown. In our previous transcriptome analysis of broomcorn millet (P.miliaceum) under salt stress [60], we observed that a substantial number of bZIP genes were differentially expressed during seed germination. This suggests that bZIP transcription factors may play important regulatory roles in the stress tolerance mechanisms of this highly resilient cereal crop. This study systematically analyzes biological information about bZIP TFs in broomcorn millet, aiming to provide a reference for the identification of various functions.
2. Materials and Methods
2.1. Genome-Wide Identification and Prediction of Physicochemical Properties
Broomcorn millet genomic sequences, coding region sequences (coding sequence, CDS), and protein sequences were downloaded from NCBI under BioProject number PRJNA431363 (https://www.ncbi.nlm.nih.gov/bioproject/?term=PRJNA431363, accessed on 23 June 2022). We referred to the Pfam number (PF00170) of bZIP transcription factors in the Pfam database (http://pfam.xfam.org/, accessed on 23 June 2022) and used the HMMER program based on a Hidden Markov Model (3.3.2) to search for the candidate bZIP proteins in broomcorn millet. The HMM profile was used to perform an hmmscan search against the P.miliaceum protein database with an E-value threshold of 1 × 10^−5^. We used protein sequences with a result greater than “0” as candidate sequences, and then, the candidate gene protein sequence was extracted using TBtools (v1.0692) [61]. Finally, all candidate protein sequences were further detected and identified via CDD (http://www.ncbi.nlm.nih.gov/cdd/, accessed on 22 June 2022) and PFAM, and proteins without bZIP domains were removed. The proteins were named according to the location of the bZIP transcription factor on the chromosome. The theoretical isoelectric point and molecular weight of the bZIP transcription factor proteins in broomcorn millet were calculated using ExPASy—Compute pI/Mw (https://web.expasy.org/compute_pi/, accessed on 25 June 2022).
2.2. Phylogeny Analyses and bZIP Domain Amino Acid Sequence Alignment
A.thaliana bZIP TF protein sequences were downloaded from the database TAIR (https://www.arabidopsis.org/index.jsp, accessed on 2 July 2022), and S.italica bZIP TF protein sequences were downloaded from the database Phytozome v12 (http://phytozome.jgi.doe.gov/pz/portal.html, accessed on 2 July 2022). A bootstrap neighbor-joining (NJ) evolutionary tree was created using MEGA 6.06 (https://www.megasoftware.net/, accessed on 7 July 2022) software with 1000 bootstrap replicates based on the sequence alignments. Additionally, the sequence alignment of the bZIP domain from broomcorn millet was performed using Clustal X 1.8.
2.3. Motif and Intron/Exon Gene Structure Analysis
The MEME v5.3.0 online service (http://meme-suite.org/tools/meme, accessed on 13 August 2022) was employed to identify conserved motifs in PmbZIP proteins, using parameters that included a maximum of 12 motifs and an optimal motif width ranging from 6 to 50 amino acids. Afterward, all identified motifs were annotated using InterProScan (http://www.ebi.ac.uk/Tools/pfa/iprscan/, accessed on 16 August 2022), and the gene structure display server program (http://gsds.cbi.pku.edu.cn/, accessed on 30 August 2022) was used to draw the gene structures of PmbZIPs.
2.4. Promoter Analysis
The 2000 base pairs preceding the initiation codon of each PmbZIP gene was obtained. These sequences were analyzed to find cis-elements using the PlantCARE online tool (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 17 October 2022), and the outcomes were visualized with TBtools (v1.0692) [61].
2.5. RNA Isolation and bZIP Gene Expression Analysis
The samples of yumi1 and yumi9, which were grown in 0 mM of NaCl (RO water) and 250 mM NaCl, respectively, were collected at 0 h and 3 h, and dew white seeds were harvested after continuous light conditions commenced. (There were three independent biological replicates for each sample). The samples were swiftly frozen with liquid nitrogen and stored at −80 °C. In total, thirty-six samples were employed for RNA-Seq and differential expression analyses.
For transcriptome analysis, total RNA was isolated from seeds using the RNA prep Pure polysaccharide polyphenol plant total RNA extraction kit (DP441) (TIANGEN, Beijing, China). Using 1% agarose gels tested RNA degradation and contamination, and RNA purity was evaluated using a NanoPhotometer^®^ spectrophotometer (IMPLEN, Munich, Germany). RNA concentration was measured using Qubit^®^ RNA Assay Kit in Qubit^®^ 2.0 Flurometer (Life Technologies, Carlsbad, CA, USA). The Agilent Bioanalyzer 2100 system’s RNA Nano 6000 Assay Kit was used to assess RNA integrity (Agilent Technologies, Santa Clara, CA, USA).
A differential expression analysis of two conditions/groups was performed. The DESeq R package (version 1.10.1) was used to analyze differential expression between two conditions or groups. DESeq offers statistical methods to identify differential expression data, utilizing a model based on the negative binomial distribution. The Benjamini and Hochberg method was used to adjust the resulting p-values to control the false discovery rate, and differentially expressed genes were those with an adjusted p-value under 0.05. The creation of heatmaps and the cluster analysis of PmbZIPs were accomplished using TBtools software [61].
3. Results
3.1. Identification of PmbZIPs in Broomcorn Millet
To identify bZIP genes in the complete P. miliaceum genome, the Hidden Markov Model (HMM) profile file of the bZIP domain (PF00170) was exploited as a query file for a search across the P. miliaceum protein sequence data, and the Pfam and CDD databases were used to confirm the presence of the complete bZIP domain. As shown in Table 1, we identified 144 PmbZIP genes in the P. miliaceum genome after removing redundant sequences and designated them as PmbZIP1 to PmbZIP144 according to their chromosome locus. Moreover, the physical and chemical properties of the 144 PmbZIPs were analyzed, such as amino acid (aa) length, molecular weight (MW), and protein isoelectric (PI) points. Chromosomal localization shows that there are 14 PmbZIPs in Chr5, which has the most bZIP genes, but Chr15, Chr16, and Chr17 only have 2 PmbZIPs. Genome sequence analyses were conducted and showed that PmbZIPs ranged from 369 base pairs (bp, PmbZIP100, PmbZIP114) to 12,828 bp (PmbZIP45). Protein sequence analyses showed that the PmbZIPs ranged from 78 aa (PmbZIP59) to 650 aa (PmbZIP51) in length. The predicted MWs ranged from 9.19 kDa (PmbZIP59) to 68.3 kDa (PmbZIP51), and the PIs ranged from 4.52 (PmbZIP102) to 11.57 (PmbZIP43).
3.2. Phylogenetic and Sequence Conservation Analysis of PmbZIPs
A phylogenetic analysis was performed with all 144 identified PmbZIP proteins, as well as 75 Arabidopsis and 78 foxtail millet bZIP family members (Figure 1 and Figure S1), using the neighbor-joining (NJ) algorithm in MEGA software (MEGA6.06). The 144 PmbZIP genes were divided into ten groups (designated as A to E, G, H, I, S, and U) according to the subfamilies classified for Arabidopsis. Based on phylogenetic relationships, Group S contains 29 members and is the largest subfamily, and the smallest subfamilies are Groups B, E, and U, with only 3 members in each. It is interesting to note that no PmbZIP member was assigned to Group F. A total of 22 PmbZIPs were classified as belonging to Group A, 25 to Group D, 23 to Group I, 21 to Group G, 8 to Group C, and 7 to Group H.
3.3. Motif and Structural Analysis of PmbZIPs
To investigate the protein sequence features of PmbZIPs, 12 different motifs were identified in PmbZIPs (Figure 2), with lengths ranging from 21 to 50 aa. The phylogenetic analysis showed that the same clusters of PmbZIPs had similar conserved domain compositions, and obvious differences between different groups were also found. Motif 1, as a “basic region” of the bZIP domain, existed in all PmbZIPs. Additionally, motifs 4 and 7, as two different “leucine zippers” of the bZIP domain, existed in Groups B, C, E, G, I, and S and Groups A, D, H, and U, respectively. Specifically, the PmbZIPs in Group D contain the most motif types, including motifs 1, 2, 3, 5, 7, 8, and 11. However, only motif 1 was present in PmbZIP135, PmbZIP128, PmbZIP38, and PmbZIP23 in Group S. This may be the reason why different subfamilies have different functions.
The structural diversity of the PmbZIP family was analyzed in terms of the exon/intron arrangement of the coding sequences. The number of introns in PmbZIPs ranged from zero to twelve. The detailed gene structure of PmbZIPs is pictured in Figure 3. Twelve introns were identified in PmbZIP104 and PmbZIP112, whereas 24 PmbZIPs were identified as intronless. Subgroups G and D generally contain more than seven introns, while subgroup S often has no introns. Most closely related PmbZIPs in the same class or subfamily share a similar gene structure in terms of the number of introns.
3.4. Promoter Analysis of PmbZIPs
To predict the biological function of PmbZIPs, 2000 bp upstream sequences from the translation start sites of PmbZIPs were analyzed using the PlantCARE database (Figure 4). The promoter of each PmbZIP consists of several cis-acting elements, such as phytohormone-responsive elements, MYB binding sites, light-responsive elements, anoxic-specific inducibility elements, abiotic stress-responsive elements, defense- and stress-responsive elements, seed-specific regulation elements, and root-specific elements. As illustrated in Table 2, a light-responsive element was identified in the promoters of all PmbZIPs. An abscisic acid-responsive element, methyl jasmonate (MeJA)-responsive element, and meristem expression element were identified in the promoters of 135, 128, and 99 PmbZIP genes, respectively. The promoters of 66 PmbZIPs contained an auxin-responsive element, 75 PmbZIPs contained a gibberellin-responsive element, and 55 PmbZIPs contained a salicylic acid-responsive element. Additionally, an MYB binding site, defense- and stress-responsive elements, a low-temperature-responsive element, and a zein metabolism regulation element were all found in 97, 33, 64, and 46 PmbZIPs, respectively. In total, PmbZIP143 promoters contained 73 (maximum) cis-elements, which included 34 light-responsive elements, 17 abscisic acid-responsive elements, 14 MeJA-responsive elements, 7 anoxic-specific inducibility elements, and 1 meristem expression element. However, PmbZIP21 promoters only contained 11 cis-elements. These findings demonstrate that PmbZIPs might be associated with various transcriptional regulations involving development, hormones, and stress responses.
3.5. Expression Analysis of PmbZIPs in Seed Germination Under Salt Stress
To explore the expression patterns of these millet bZIP genes, we used RNA-seq data of yumi1 (Y1) and yumi9 (Y9) under salt stress in the seed germination stage. Based on the millet RNA-seq data, 67 bZIP genes were detected in all three sampling stages at the gene level (Figure 5). This suggests that nearly half of bZIP genes are broadly expressed during millet germination and development. In addition, the fact that 18 bZIP genes have different expression levels suggests that these genes were induced by salt and have a vital function in tolerance responses. The heatmap analysis found that PmbZIP6, PmbZIP11, PmbZIP15, PmbZIP71, PmbZIP78, PmbZIP89, and PmbZIP97 are downregulated salinity-responsive genes (SRGs). Upregulated SRGs include PmbZIP26, PmbZIP30, PmbZIP33, PmbZIP65, PmbZIP70, PmbZIP104, PmbZIP107, PmbZIP113, PmbZIP118, PmbZIP125, and PmbZIP131. Interestingly, the differential expression of most SRGs occurs when water is imbibed for 3 h or during radicle protrusion (RAP) in the seed envelope stage, thus indicating that the seeds reinitiate metabolic processes and stress response in this period.
4. Discussion
In the immediate future, broomcorn millet will become a crucial crop that can help in diversifying agriculture and promoting a healthier diet for humans. Several plant bZIP transcription factor families have been identified and characterized extensively, which play a vital role in the regulation of growth, development, and environmental response. However, these gene families have not been reported in broomcorn millet. Therefore, searching for and identifying advantageous bZIP genes in millet will facilitate further basic scientific research on molecular breeding processes.
4.1. Characterization of Broomcorn Millet bZIP Gene Family
The genes involved in genome replication events can evolve into genes with new functions, which play an important role in expanding genome content and diversifying gene functions [62]. Previous research showed that the emergence of the broomcorn millet genome was the result of the ~5.6 MYA hybridization of two closely related genomes. Most Panicum species are polyploids native to the tropical/semi-arid regions of the world. Many gene families in the millet genome are double copies, most of which are retained by the parental species and single-copy genes of the parental species [6]. Since there are 78 bZIP genes in the foxtail millet genome, 144 bZIP genes were predicted to be in the P. miliaceum genome. In addition, many subfamily bZIP genes are double copies in broomcorn millet, except those in Groups B, E, F, and U; this is consistent with previous research. After genome duplication, nonfunctionalization (duplicated genes are silenced), subfunctionalization (function is partitioned between the new paralogs), and neofunctionalization (duplicated genes gain new functions) generally take place [63,64,65]. In this study, we found that Group F genes could have been lost or had changed functions during their evolution, thus suggesting that there was extensive gene loss during genome duplication.
A phylogenetic analysis of the PmbZIP gene family revealed that subfamilies A, D, and S contain a relatively large number of members that form well-supported and tightly clustered groups, thus suggesting potential functional conservation and lineage-specific expansion. Notably, the S subfamily appears to be divided into four smaller clades, which are interspersed among the members of the G, A, and C subfamilies. This branching pattern suggests that during evolution, some ancestral S-type bZIP members may have undergone functional divergence and structural differentiation, thus giving rise to new regulatory subgroups such as G, A, and C. These findings support the hypothesis that bZIP subfamily diversification has been driven by both sequence divergence and the acquisition of specialized regulatory roles in different stress and developmental contexts.
Among these subfamilies, Group A bZIP genes stand out due to their well-established roles in abscisic acid (ABA)-mediated stress responses. In our study, PmbZIP30 (PmABI5) and PmbZIP131, both belonging to Group A, were significantly induced by ABA during seed development in broomcorn millet [60]. This is consistent with findings in Arabidopsis and rice, where homologous Group A bZIPs (e.g., ABI5) act as core components in the ABA signaling pathway. Furthermore, cross-species evidence supports the involvement of Group A bZIPs in the conserved PP2C–SnRK2–AREB signaling module, which regulates plant responses to drought and other abiotic stresses. The promoter architecture and expression dynamics of Group A PmbZIP genes in this study reinforce the notion that their functional roles in ABA-dependent stress signaling are evolutionarily conserved. These results not only provide insight into the diversification and conservation of bZIP transcription factors in broomcorn millet but also highlight the adaptive significance of Group A members in stress tolerance.
4.2. Structural Analysis of PmbZIPs
The detailed understanding of the functional domain of A. thaliana and S. italica bZIPs enabled us to analyze similar domains within the broomcorn millet bZIP gene family. In this research, all 144 PmbZIP proteins contain the necessary basic domain, which provides the structural basis for their conserved function. Moreover, different subgroups have different ZIP motifs, which makes their functions differ. Groups A, D, H, and U have type I ZIP domains (motif 7, K-L-X7-R), and the main function of these subfamily genes is the regulation of biological and abiotic stresses. However, Groups B, C, E, G, I, and S have type II ZIP domains (motif 4, V-L-X8-R) that are involved in carbohydrate biosynthesis, post-transcriptional inhibition, development, and hormone synthesis. It has been reported that intron retention regulates protein isoform production, RNA stability and translation efficiency, and the rapid induction of expression via the post-transcriptional splicing of retained introns [66]. An analysis of the bZIP gene structure revealed that most intronless PmbZIPs occurred in Groups S and C, and a similar observation was reported for banana (Musa spp. L.) and switchgrass (P. virgatum L.) [67,68]. The PmbZIPs of subgroups A, C, and I, with relatively fewer introns, were associated with stress responses. This conclusion aligns with the results of the present and past studies [60].
4.3. Cis-Element Analysis in the Promoters of PmbZIPs
The cis-elements of different transcription factors have different functions. The number and form of cis-elements in promoter regions could play an essential role in the regulation of gene expression. The results illustrate that abiotic stress-related cis-elements, including abscisic acid-responsive elements, anoxic-specific inducibility elements, low-temperature-responsive elements, MeJA-responsive elements, MYB binding sites, salicylic acid-responsive elements, and wound-responsive elements, are major regulatory elements in the PmbZIP promoters activated by ABA, NaCl, or other forms of abiotic stress. In addition, many development-related cis-elements were also found, such as auxin-responsive elements, endosperm expression elements, gibberellin-responsive elements, root-specific elements, seed-specific regulation elements, and meristem expression elements. These findings suggest that light-responsive elements, abscisic acid-responsive elements, and MeJA-responsive elements play a vital role in transcriptional regulation in broomcorn millet. PmbZIP promoters present a lot of stress-responsive cis-elements and hormone response cis-elements, which indicates their potential roles in the response to stress and pathogen infections. Consequently, PmbZIPs are often taken as candidate genes to understand the responses to biotic stresses and plant development.
4.4. PmbZIP Involvement in Development and Stress Response
Previous reports have revealed that bZIP TFs function in many stress responses and development by regulating diverse biochemical and physiological pathways [17,69,70,71]. bZIP transcription factors possess different characteristics in different species. The overexpression of TabZIP15 in wheat can enhance root length and fresh weight during salt stress, thus suggesting that the TabZIP15 gene is involved in the regulation of wheat salt stress tolerance [72]. The homozygous T-DNA insertional mutants Osabf1-1 and Osabf1-2 are more sensitive in response to drought and salinity treatments than wild-type plants, and the OsNAC, OsLEA3, and OsABA45 genes are significantly suppressed in Osabf1 mutants. Hence, OsABF1 likely plays a positive role as an ABA-responsive transcription factor in abiotic stress signaling [26]. We incorporated and highlighted experimental data from our previous work, wherein PmbZIP30 was overexpressed in rice. The transgenic rice lines exhibited significantly shorter seed germination times compared to the wild-type, thus indicating a positive regulatory role of PmbZIP30 in seed germination and stress response. This functional validation supports the biological relevance of our bioinformatic findings (Figure S2). The overexpression of StbZIP65 in Arabidopsis enhanced salt tolerance [73]. However, some research shows that GmbZIP19 expression is significantly induced by ABA (abscisic acid), JA (jasmonic acid), and SA (salicylic acid) but is reduced under salt and drought stress conditions, thus suggesting that GmbZIP19 is a positive regulator of pathogen resistance and a negative regulator of salt and drought stress tolerance [74]. TabZIP6 can bind to the promoters of CBFs and decrease the expression of downstream COR genes in TabZIP6-overexpressing Arabidopsis seedlings; therefore, TabZIP6 is a negative regulator in the cold stress response [75].
In this study, a total of 144 PmbZIP genes were identified in the P. miliaceum genome and classified into eleven subfamilies based on phylogenetic relationships. Conserved motif and domain structure analyses revealed that the members within each subfamily exhibit high sequence conservation. The promoter analysis indicated that PmbZIP genes may be involved in multiple hormone signaling pathways and environmental stress responses, as reflected by the presence of diverse cis-acting regulatory elements, including MYB binding sites. The transcriptome analysis further showed that 18 PmbZIP genes were differentially expressed during seed germination under salt stress, thus suggesting their potential regulatory roles in abiotic stress adaptation. These findings provide a valuable resource for understanding the functional roles of bZIP transcription factors in broomcorn millet and lay the foundation for future stress resilience breeding and gene function studies.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Lu H. Zhang J. Liu K.-B. Wu N. Li Y. Zhou K. Ye M. Zhang T. Zhang H. Yang X. Earliest domestication of common millet (Panicum miliaceum) in East Asia extended to 10,000 years ago Proc. Natl. Acad. Sci. USA 20091067367737210.1073/pnas.090015810619383791 PMC 2678631 · doi ↗ · pubmed ↗
- 2Yue H. Wang M. Liu S. Du X. Song W. Nie X. Transcriptome-wide identification and expression profiles of the WRKY transcription factor family in Broomcorn millet (Panicum miliaceum L.)BMC Genom.20161734310.1186/s 12864-016-2677-327165545 PMC 4862231 · doi ↗ · pubmed ↗
- 3Rajput S.G. Santra D.K. Schnable J.J.M.B. Mapping QT Ls for morpho-agronomic traits in proso millet (Panicum miliaceum L.)Mol. Breed.2016363710.1007/s 11032-016-0460-4 · doi ↗
- 4Hunt H.V. Badakshi F. Romanova O. Howe C.J. Jones M.K. Heslop-Harrison J.S. Reticulate evolution in Panicum (Poaceae): The origin of tetraploid broomcorn millet, P. miliaceum J. Exp. Bot.2014653165317510.1093/jxb/eru 16124723408 PMC 4071833 · doi ↗ · pubmed ↗
- 5Wang H. Wang J. Chen C. Chen L. Li M. Qin H. Tian X. Hou S. Yang X. Jian J. A complete reference genome of broomcorn millet Sci. Data 20241165710.1038/s 41597-024-03489-538906866 PMC 11192726 · doi ↗ · pubmed ↗
- 6Zou C. Li L. Miki D. Li D. Tang Q. Xiao L. Rajput S. Deng P. Peng L. Jia W. The genome of broomcorn millet Nat. Commun.20191043610.1038/s 41467-019-08409-530683860 PMC 6347628 · doi ↗ · pubmed ↗
- 7Baltensperger D.D. Foxtail and Proso Millet Progress in New Crops ASHS Press Alexandria, VA, USA 1996
- 8Washburn J.D. Schnable J.C. Davidse G. Pires J.C. Phylogeny and photosynthesis of the grass tribe Paniceae Am. J. Bot.20151021493150510.3732/ajb.150022226373976 · doi ↗ · pubmed ↗