Mouse PrimPol Outperforms Its Human Counterpart as a Robust DNA Primase
Gustavo Carvalho, Susana Guerra, María I. Martínez-Jiménez, Luis Blanco

TL;DR
Mouse PrimPol is more effective than human PrimPol in starting DNA synthesis, due to structural differences that enhance its primase activity.
Contribution
The study reveals structural and functional differences between mouse and human PrimPol that explain the superior primase activity in mice.
Findings
MmPrimPol has a more powerful primase activity than human PrimPol.
The shorter linker in MmPrimPol promotes a primase-ready configuration.
MmPrimPol forms primers more efficiently but has limited DNA polymerization on existing primers.
Abstract
The human PrimPol counteracts DNA replication stress by repriming DNA synthesis when fork progression is hindered by UV light or hydroxyurea treatment, or by encountering complex DNA structures, such as G-quadruplexes, R-loops, or interstrand crosslinks. The Mus musculus PrimPol (MmPrimPol) shares a high degree of amino acid similarity with its human ortholog; however, as shown here, MmPrimPol exhibits a more powerful primase activity compared to the human enzyme. Such a robust primase activity relies on an enhanced ability to bind the 5′ site nucleotide, and consequently to form initial dimers and further mature primers. Additionally, a shorter linker between the AEP core and the Zn finger domain (ZnFD) in the murine homolog likely promotes a constitutive closing of these domains into a primase-ready configuration. Consequently, a reinforced close configuration of the ZnFD would…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
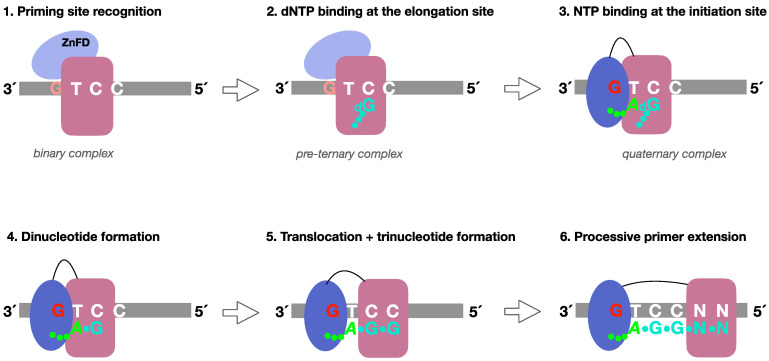 Figure 1
Figure 1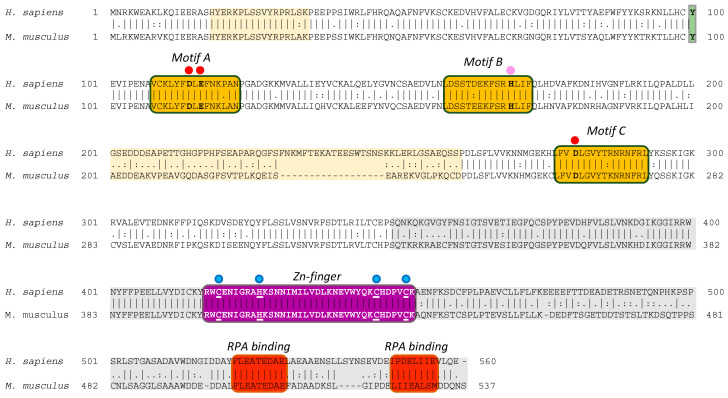 Figure 2
Figure 2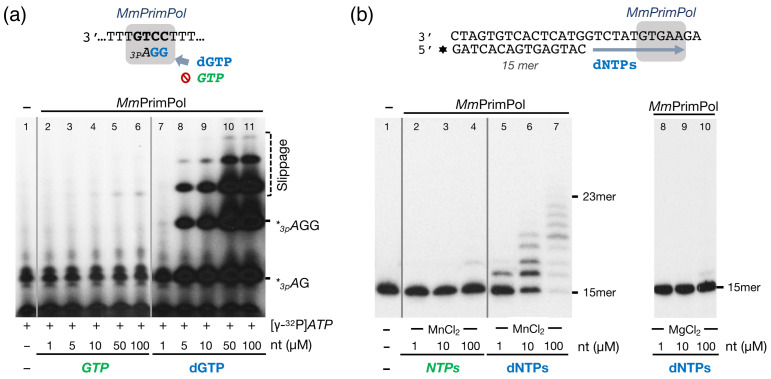 Figure 3
Figure 3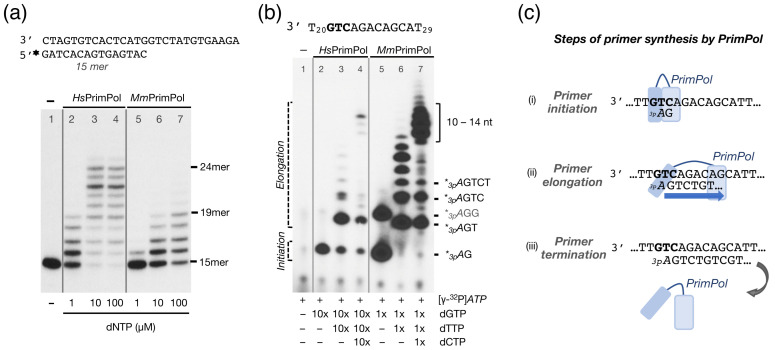 Figure 4
Figure 4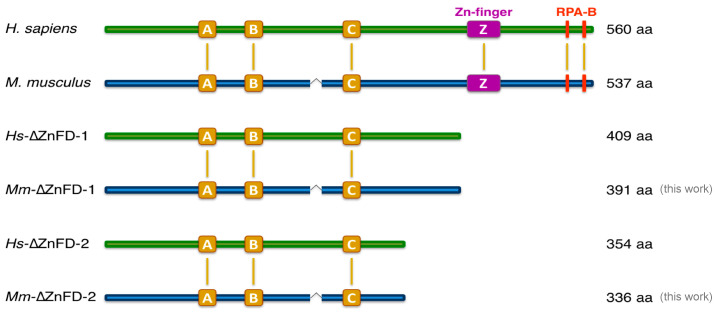 Figure 5
Figure 5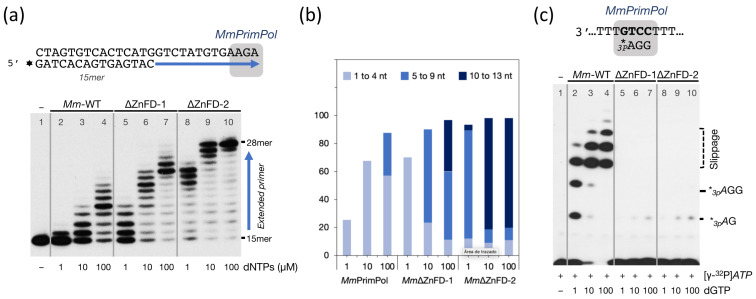 Figure 6
Figure 6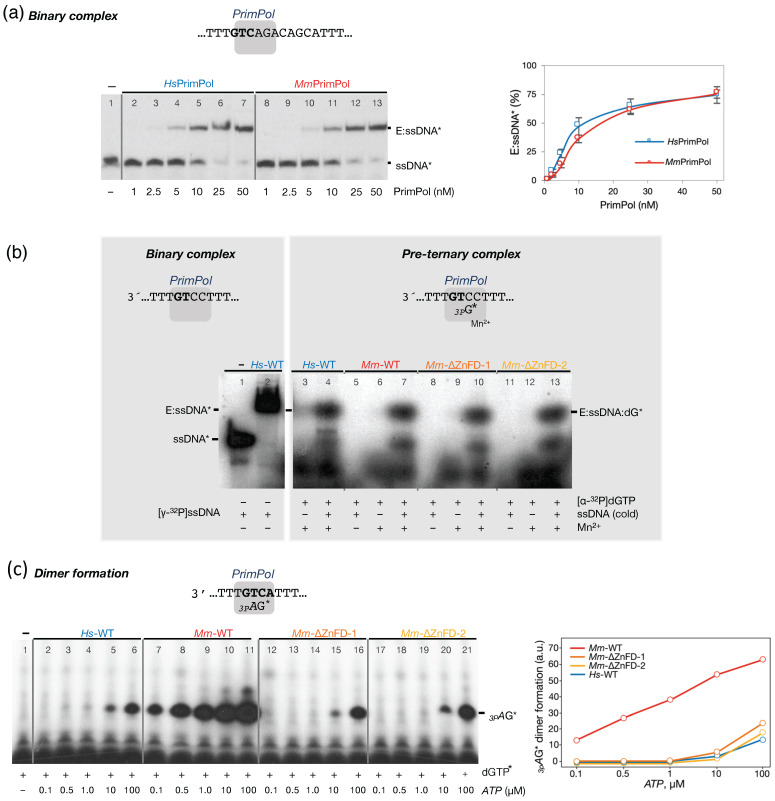 Figure 7
Figure 7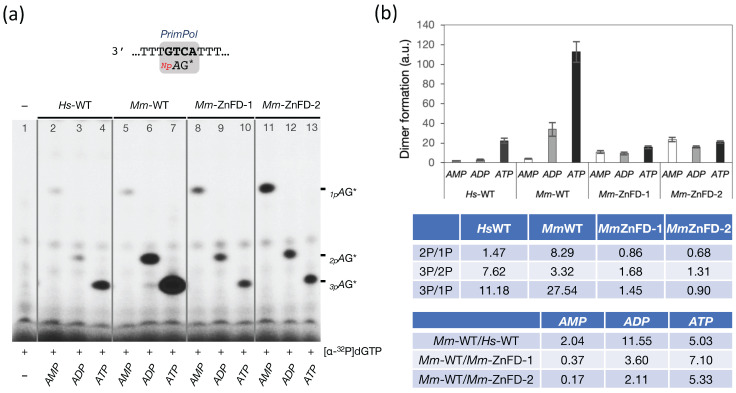 Figure 8
Figure 8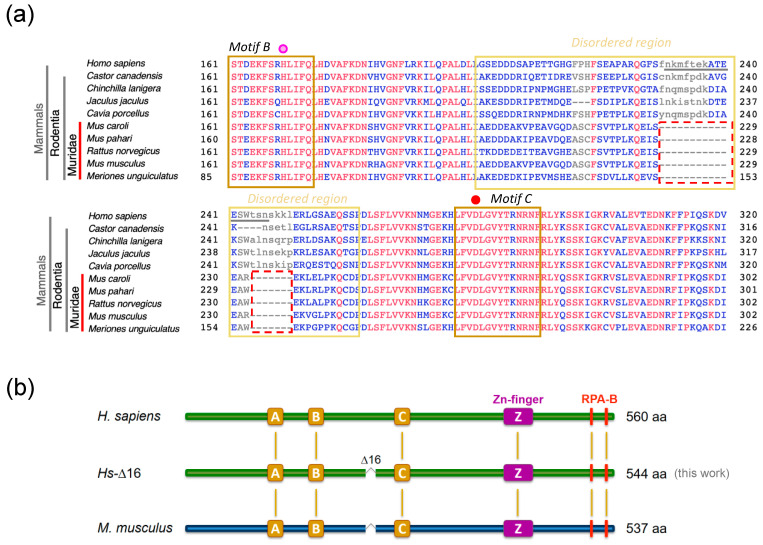 Figure 9
Figure 9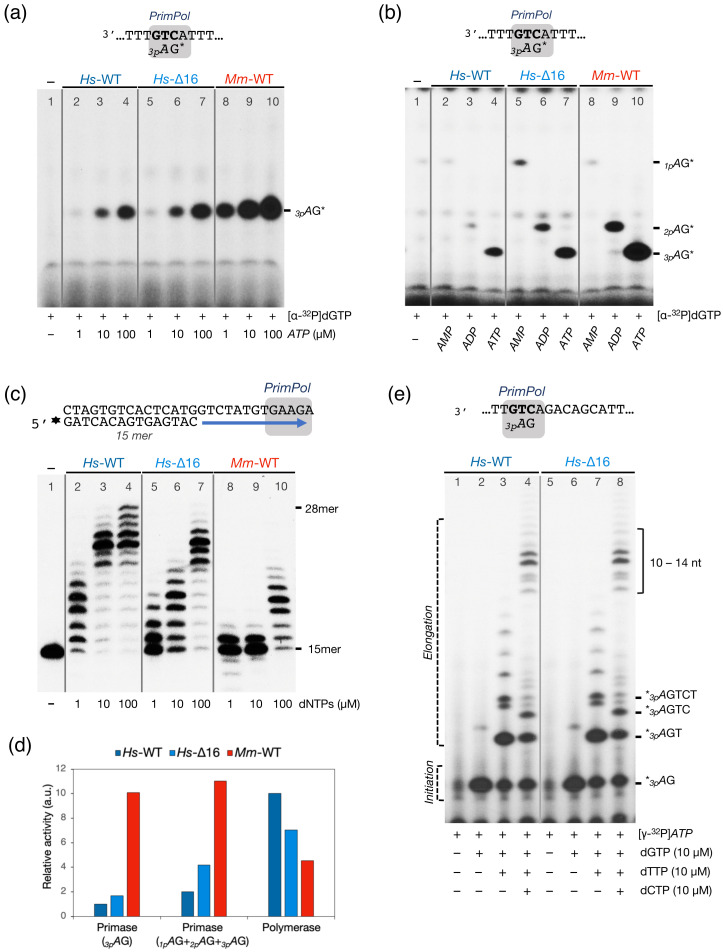 Figure 10
Figure 10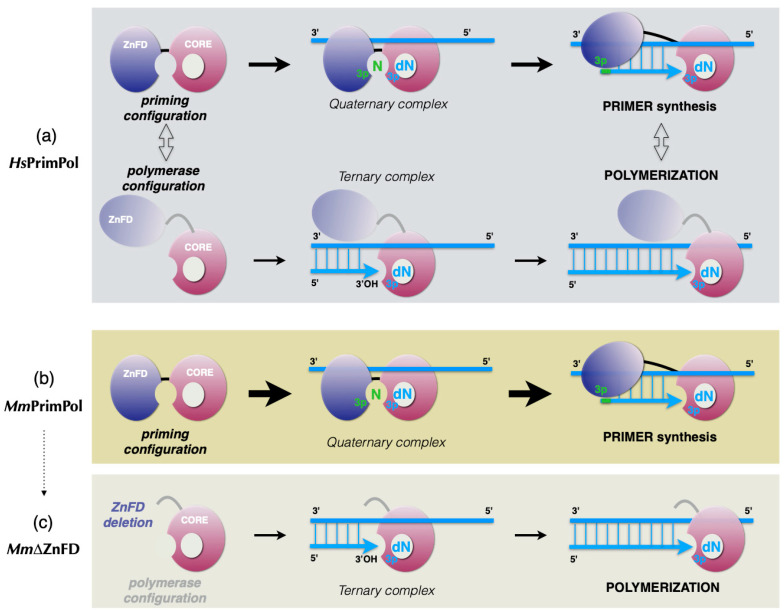 Figure 11
Figure 11- —MINECO
- —European Union Next Generation EU/PRTR
- —Brazilian National Council for Scientific and Technological Development
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsDNA Repair Mechanisms · CRISPR and Genetic Engineering · Molecular Biology Techniques and Applications
1. Introduction
Primases are enzymes responsible for initiating DNA replication, by making primers for the replicative DNA polymerases [1,2,3,4]. Humans possess two members of the archaeo-eukaryotic primases (AEP) family: the conventional dimeric primase (PriL–PriS), which is part of the Polα/primase complex and synthetizes RNA primers to initiate Okazaki fragments during lagging strand replication [5,6], and the monomeric PrimPol (HsPrimPol), which has the ability to synthetize the DNA primers [7,8] that are essential for rescuing stalled replication forks during leading strand synthesis [9]. Both human primases share a high similarity in the catalytic domain. HsPrimPol is characterized by three signature motifs: a highly conserved DxE motif (motif A, Asp^114^ and Glu^116^), which together with Asp^280^ residue from motif C, forms the divalent metal binding site [8,10,11], and motif B (formed by Lys^165,^ Ser^167^ and His^169^) along with Arg^291^ and Lys^297^ bind the incoming 3′ nucleotide mainly through its phosphates [11,12,13]. One specific difference between these two types of the AEP primases is that a single residue in PrimPols (Tyr^100^ in HsPrimPol) contributes to nucleotide selectivity by acting as a steric gate that favors dNTP incorporation [14]. Additionally, the C-terminal domain of HsPrimPol contains a highly conserved motif (Cys–X_5_–His–X_19_–Cys–X_4_–Cys), predicted to form a functional Zn-finger, which is essential for the HsPrimPol primase activity [9,11,15,16], followed by two motifs that interact with the human single-stranded binding protein RPA [11,17].
The C-terminal domain containing the Zinc-finger (ZnFD) in eukaryotic PrimPols is highly similar to that found in the AEP primases from the nucleo-cytoplasmic large DNA virus family [7], such as herpes virus primase (UL52), which also requires its C-terminal ZnFD for primase activity [18,19]. A double point mutation in two of the Zn^2+^ ligands (C419G, H426Y) and a deletion mutant lacking the C-terminal domain (ΔZnFD, missing amino acids 410–560) were instrumental to demonstrate that this domain is involved in DNA priming, and that PrimPol’s primase function is required in vivo to re-start stalled replication forks [9]. On the other hand, the ZnFD is not necessary for conventional DNA polymerase activity, or for translesion synthesis (TLS) during primer extension [9,15].
The initial steps of primer synthesis by HsPrimPol and the role of the ZnFD at each individual step were delineated as follows [16] (Figure 1, see legend for further details):
- (1)Formation of a PrimPol/ssDNA binary complex.
- (2)Further interaction with the incoming 3′-deoxynucleotide (pre-ternary complex).
- (3)The ZnFD is mobilized to facilitate binding and selection of the 5′-nucleoside triphosphate (preferentially a ribonucleotide), that will become the first nucleotide of the newly synthesized primer.
- (4)Once the 5′-nucleotide (acting as a primer) and the 3′-nucleotide are selected and a quaternary complex is formed, catalysis of the initial dimer occurs.
- (5)A maintained interaction of the ZnFD with the 5′-terminal triphosphate is essential for the subsequent translocation and insertion event to form a trimer.
- (6)Further elongation of the primer occurs processively, i.e., with no dissociation, thanks to the sustained interaction of the ZnFD with the initiating nucleotide. When the primer reaches an optimal length, PrimPol dissociates and the primer is extended by the replicative DNA polymerase.
The in vivo role of the human PrimPol has been mainly determined using human cell lines [20,21,22,23] but also taking advantage of chicken DT40 cells [24] and even a mouse model of PrimPol deficiency. Our laboratory established a C57BL/6 mice colony with a knock-out for the CCDC111 gene, encoding PrimPol. This CCDC111–/– colony was very relevant to study the physiological role of PrimPol, and to generate mouse embryonic fibroblasts (MEFs) for in vivo assays that revealed its nuclear and mitochondrial functions [8,9,14,25]. Strikingly, mouse cells lacking the endogenous murine PrimPol were poorly rescued by ectopic expression of the human PrimPol ortholog [14] suggesting that the two PrimPols may have important differences in their enzymatic activity. Therefore, and because only the activity of the human PrimPol has been described to date, we aimed to study the enzymatic activities of the mouse PrimPol. As shown here, MmPrimPol and HsPrimPol share similar characteristics, but unexpectedly large differences in specific activity. MmPrimPol exhibits a robust primase activity, also dependent on the ZnFD, and a poor DNA polymerase activity, which further supports PrimPol’s physiological relevance as a primase, and suggests that the constitutive configuration of MmPrimPol is ready to prime. Cross-species comparisons provide invaluable insights into enzyme evolution, mechanistic diversity, and physiological adaptation. This study exemplifies how such analyses can uncover both conserved and specialized functions, advancing our understanding of DNA replication across organisms.
2. Results
2.1. The Mouse and Human PrimPol Share High Amino Acid Sequence Similarity
MmPrimPol is codified by the CCDC111 gene, which is located on chromosome 8 (8 B1.1) and spans over 40 kb (NCBI Gene ID: 408022). It is transcribed into a 3835 bp mRNA (NCBI Reference sequence: NM_001001184.2) which expresses a 537- amino acid polypeptide (NCBI Ref. seq.: NP_001001184.1) with a predicted molecular weight of 61.34 kDa. Amino acid sequence alignments have revealed an 80% similarity between the human and mouse PrimPols (Figure 2). Like HsPrimPol, MmPrimPol contains the conserved catalytic motifs A, B, and C characteristic of the AEP primases, along with a highly conserved Zinc-finger and two C-terminal RPA binding motifs. As expected, the catalytic residues of Asp^114^ and Glu^116^ (DxE) in motif A of HsPrimPol, which serve as metal ligands, are invariantly conserved in MmPrimPol; in addition, the third catalytic residue, Asp^262^ in MmPrimPol (corresponding to Asp^280^ in HsPrimPol), located in motif C is also conserved (indicated with red dots in Figure 2). The His^169^ residue in motif B, critical for binding the incoming 3′-nucleotides [11,12,26], is also conserved in both PrimPols (Figure 2). The carboxy terminal region of both PrimPols contains a Zn-finger, formed by three cysteines and one histidine (Cys–His–Cys–Cys, indicated by blue dots in Figure 2) with the potential to coordinate a Zn^2+^ atom, which is 100% identical in both PrimPols. Downstream of the ZnFD, two RPA binding sites (indicated by red boxes in Figure 2) are also 100% identical between both HsPrimPol and MmPrimPol.
A multiple sequence alignment of PrimPols from diverse organisms [8] identified a variable region situated between the catalytic motifs B and C. This region is moderately conserved and varies in length, being approximately 50 residues longer in mammals compared to insects or plants. Further alignments of mammalian PrimPols performed in this study indicate that only the Muridae family has lost an 18 aa sequence within this variable region, corresponding to amino acids 229 to 246 in HsPrimPol (Figure 2 and Figure 9). Interestingly, this variable region (residues 201 to 260, indicated by light yellow in Figure 2) was disordered in the crystal structure of HsPrimPol [26].
2.2. The Mouse PrimPol Is a True DNA Primase
HsPrimPol preferentially uses deoxynucleotides (dNTPs) over ribonucleotides as substrates at the elongation site for both primase and polymerase activities, unlike conventional RNA primases [8]. This preference for dNTPs is primarily attributed to HsPrimPol Tyr^100^, which acts as the sugar selector for incoming 3′-nucleotides; in fact, a mutation of Tyr^100^ to histidine inverts this selectivity, now allowing for the incorporation of ribonucleotides by HsPrimPol [14]. Given that Tyr^100^ is conserved in both the human and the mouse PrimPol, we presumed that MmPrimPol would also be biased to use dNTPs as 3′-incoming nucleotides. DNA primase activity was firstly assessed by using as a template the 29-mer single-stranded pyrimidine-rich oligonucleotide 3′ T_10_-GTCC-T_15_ 5′ (hereafter referred to as 3′ GTCC 5′). This 3′GTC sequence was previously identified as a preferred initiation site for HsPrimPol using in vitro assays [8], and as a favored template context for various viral, prokaryotic, and eukaryotic RNA primases [27], thus predicting its suitability for measuring the MmPrimPol primer synthesis. In addition to the 3′ GTCC 5′ template, a low concentration (16 nM) of [γ-^32^P]ATP was provided as the initiating 5′-nucleotide, to be positioned opposite the template’s T (GTCC), and increasing concentrations of GTP or dGTP as the only 3′-nucleotide, to be inserted opposite the two consecutive Cs in the template (GTC****C), thus forming di- and trinucleotide products, as outlined in the scheme in Figure 3a. Manganese ions (1 mM) were supplied as the preferred metal co-factor for PrimPol activity. As expected, MmPrimPol was unable to initiate a de novo synthesis of RNA primers when ATP and GTP were supplied as substrates, even with GTP concentrations up to 100 µM (Figure 3a, lanes 2–6); however, MmPrimPol exhibited robust DNA primase activity when dGTP was provided as the 3′-nt, synthesizing _3p_AG dimers when dGTP was supplied at a concentration as low as 5 µM (Figure 3a, lanes 8–11). Increasing dGTP concentrations led to the production of canonical trimers (_3p_AGG), and to longer non-canonical products (such as _3p_AGGG or _3p_AGGGG), obtained by backwards slippage of the last incorporated G to be realigned with the first C in the template, yielding to the iterative insertion of additional Gs opposite the second consecutive C of the template, as previously described for the human PrimPol [8].
The sugar selectivity of MmPrimPol was further explored during a conventional polymerization assay. For this, a pre-synthesized DNA primer was hybridized to a DNA template with a 13-nt overhang, and polymerization was tested in a reaction mixture containing the four NTPs or dNTPs. As shown in Figure 3b (lanes 2–4), no primer extension (polymerization) by MmPrimPol was observed with NTPs as 3′-substrates, consistent with the primase assay. Conversely, MmPrimPol extended the pre-synthesized DNA primer in the presence of dNTPs and MnCl_2_, with products increasing in yield and length in response to the concentration of dNTPs provided (Figure 3b, lanes 5–7). This confirms MmPrimPol’s preference for dNTPs at the 3′-elongation site, but the limited elongation pattern reflects a modest DNA polymerase activity. As other groups have reported that HsPrimPol can utilize magnesium as catalytic metal [11,28,29,30], we tested if MmPrimPol could be using magnesium to activate DNA polymerization. MmPrimPol showed a null polymerization capacity (Figure 3b, lanes 8–10), indicating that magnesium is not a suitable metal co-factor for MmPrimPol (at least, under our experimental conditions).
Collectively, these preliminary experiments suggested an altered balance of Primase/Polymerase activities in MmPrimPol compared to HsPrimPol [8].
2.3. The Mouse PrimPol Has a Strong Preference to Initiate and Complete Primer Synthesis, but Not to Extend Pre-Existing Primer Chains
After demonstrating that MmPrimPol has a strong capacity to initiate DNA primers, but a very limited capacity as a DNA polymerase, the balance of both activities was comparatively tested between HsPrimPol and MmPrimPol. As shown in Figure 4a, HsPrimPol was significantly more efficient (more than 10-fold) than MmPrimPol in a conventional DNA polymerization assay (qualitatively estimated to be more than 10-fold; note the same elongation pattern observed with the two PrimPols at different (10-fold) dNTP concentration: lane 2 vs. lane 6, and lane 3 vs. lane 7). For a complete evaluation of the DNA primase activity, it is important to consider that, during de novo primer synthesis, a primase has to encompass both initiation (dinucleotide formation) and its further elongation (extending the dimer to a 7–10 nucleotide primer). Given MmPrimPol’s low DNA polymerase activity, we investigated whether this would impair the elongation step of primer synthesis, leading to the generation of only short, aborted primers. To address this, we designed a longer ssDNA template containing PrimPol’s favorite priming sequence, 3′ GTC 5′, followed by a heterogeneous 8 nt sequence (3′ T_20_GTCAGACAGCAT_29_ 5′ called 3′ GTC-step 5′). This template allowed us to explore the initiation and elongation process of primer synthesis by MmPrimPol in a single assay. To delineate the sequential steps (initiation and elongation) when priming on the 3′ GTC-step 5′ template, we provided a low dose (16 nM) of the initiating 5′-nucleotide [γ-^32^P]ATP to radioactively label all nascent products, and complementary 3′-dNTPs were consecutively added to the reaction, to follow the sequential products of primer synthesis (_3p_AG, _3p_AGT, _3p_AGTC, and so on…). Of note, due to its robust DNA primase activity, MmPrimPol was assayed with a 10-fold lower dNTP concentration than HsPrimPol.
Upon addition of the first two nucleotides, ATP and dGTP, both HsPrimPol and MmPrimPol successfully initiated primer synthesis, generating _3p_AG dimers (Figure 4b, lanes 2 and 5). This indicates that both enzymes recognized their favorite priming site (GTC**)** on the new oligo sequence. Despite the 10-fold lower dGTP concentration used, the MmPrimPol primase activity resulted in a much higher formation of the _3p_AG dimer, and even the _3p_AGG trimer, likely through repetitive insertion of a second dGMP unit by slippage (Figure 4b, lane 5). With the addition of the next nucleotide (dTTP), HsPrimPol primarily produced the expected trimer (_3p_AGT), alongside some residual dimers and longer unscheduled products; whereas, MmPrimPol efficiently converted all dimers into trimers or longer primers (Figure 4b, lanes 3 and 6), the latter arising from misalignment events [12]. Finally, upon addition of a fourth nucleotide (dCTP), both PrimPols elongated their newly synthesized dimers and trimers, generating canonical extension products up to 10 to 14-nt long (Figure 4b, lanes 4 and 7). Thus, in stark contrast to our previous DNA polymerase assay (Figure 4a), MmPrimPol was perfectly capable of elongating its newly initiated primers, performing even better than HsPrimPol, despite the 10-fold lower total dNTP concentration. These results demonstrate that MmPrimPol effectively carries out “polymerization” when elongating its self-generated primers, but is very deficient to extend a pre-existing primer (unlike conventional DNA polymerases do). These results reinforce the notion that the important function of PrimPols is their DNA primase activity, and points to the Zn-finger of MmPrimPol as the main responsible for its robust DNA primase activity.
2.4. The Zinc-Finger Domain of the Mouse PrimPol Brakes Its DNA Polymerase Activity but Is Essential for Its Strong DNA Primase Activity
The AEP core of HsPrimPol has been crystallized, but is lacking the C-terminal part which includes the Zinc-finger domain (ZnFD) and the RPA binding motifs [26]. Consequently, our understanding of the ZnFD primarily comes from in vitro biochemistry assays and in vivo studies using deletion or point mutants. Previously, we demonstrated that the ZnFD of HsPrimPol is essential for primase activity by eliminating this entire domain, along with the downstream C-terminal region (Hs∆ZnFD1^(1–409)^) of HsPrimPol [9,16] (Figure 5). Conversely, the Hs∆ZnFD-1 displayed a significantly enhanced DNA polymerase activity, indicating that the ZnFD inhibits extension of a pre-existing primer. Given the altered balance between the primase and polymerase activities in MmPrimPol, we questioned whether the elimination of the Zn-finger would also be crucial for the primase activity, and could similarly enhance its DNA polymerase activity. To investigate this, we generated two MmPrimPol mutants (Figure 5) by deleting the ZnFD The first mutant, Mm∆ZnFD-1^(1–391)^, resembling our previously studied Hs∆ZnFD-1^(1–409)^, and obtained by introducing a stop codon immediately after the conserved sequence FFPEELL, localized 7 amino acids upstream the ZnFD in both the human and the mouse PrimPols (Figure 5). The second deletion mutant of MmPrimPol, Mm∆ZnFD-2^(1–336)^, which preserves the semi-conserved sequence PSQTKRK (amino acids 330–336 of MmPrimPol; the amino acids that differ from HsPrimPol are underlined), and corresponds to the crystalized form of HsPrimPol Hs∆ZnFD-2^(1–354)^ [15,26] (Figure 5).
The impact of ZnFD elimination was firstly examined by re-evaluating the limited DNA polymerase activity of MmPrimPol when extending a pre-synthesized primer (Figure 6a). Notably, both ZnFD-deletion mutants of MmPrimPol significantly stimulated its polymerization activity, generating longer products compared to the WT MmPrimPol (Figure 6a,b), and mimicking the increase in the polymerase activity reported for HsPrimPol as a consequence of deleting its ZnFD [9]. The deletion mutants generated longer products compared to the WT MmPrimPol (Figure 6a,b); moreover, a direct correlation was observed between the extent of the ZnFD deletion and the gain in DNA polymerase activity. Collectively, these findings indicated that the poor DNA polymerase activity of MmPrimPol is not attributable to a catalytic deficiency, but rather to an inhibitory effect exerted by its ZnFD. Such an inhibition is likely the consequence of a more closed configuration of both the ZnFD and the AEP, that impedes binding to a preexisting primer, but is more prone to initiate a de novo primer.
In agreement with this hypothesis, both ZnFD-depleted mutants Mm∆ZnFD-1 and Mm∆ZnFD-2 failed to initiate de novo primer synthesis, in stark contrast with the strong primase activity showed by the wild-type MmPrimPol (Figure 6c). This indicates that the ZnFD of MmPrimPol is essential for its strong DNA primase activity, in good agreement with the data reported for HsPrimPol [9,15,16].
2.5. The Mouse and Human PrimPols Form Similar Binary and Pre-Ternary Complexes as Primase Intermediates
To elucidate the basis for the robust primase activity observed in MmPrimPol, a detailed investigation of the individual steps leading to primer synthesis is essential. As previously proposed by Sheaff and Kuchta (1993) [31], primer synthesis proceeds through the following series of defined stages: formation of an enzyme/ssDNA binary complex; followed by the binding of the first nucleotide at the 3′-elongation site of the primase (pre-ternary complex); followed then by the binding of a second nucleotide at the 5′-initiation site (quaternary complex), leading to subsequent catalysis, which renders dimer formation. These fundamental steps have been well-defined for HsPrimPol [16].
Consequently, the individual steps required for primer initiation by MmPrimPol were studied, in comparison with its human ortholog. Firstly, we assessed the capacity of MmPrimPol to form a binary complex with an ssDNA template (3′ T_20_GTCAGACAGCAT_29_ 5′) using EMSA assays. Both the human and the mouse PrimPol formed a single E/ssDNA binary complex, at similar protein/ssDNA ratios (Figure 7a), therefore suggesting a similar affinity for ssDNA. Next, we investigated the binding of [α-^32^P]dGTP (hereafter referred as dGTP*) at the 3′site of both PrimPols in the presence of ssDNA (pre-ternary complex). Upon providing both the ssDNA template 3′ GTCC 5′ and dGTP*, the labeled deoxynucleotide is expected to occupy the elongation site, opposite the dC at the 3′GTC5′ priming site. Paradoxically, the first nucleotide bound by PrimPol (and other primases) ultimately becomes the second nucleotide in the newly synthesized dimer. As shown by the EMSA assays, a PrimPol/ssDNA/dGTP* pre-ternary complex (Figure 7b) was formed with either the human or the mouse PrimPol with similar efficiency, revealing that the binding of the 3′-dNTP at the elongation site does not account for the robust priming activity of MmPrimPol. The pre-ternary complex (MmPrimPol/ssDNA/dGTP*) was formed only in the presence of Mn^2+^ ions (Figure 7b), underscoring the crucial role of the activating metal for 3′dNTP binding. An analysis of the two Mm∆ZnFD deletion mutants demonstrated their ability to form the pre-ternary complex (Figure 7b). These results allow us to conclude that the murine ZnFD is not required for the binding of the incoming 3′-dNTP, also in agreement with previous studies with HsPrimPol [16].
2.6. The Zn-Finger Domain of the Mouse PrimPol Confers High Affinity for the Initiating 5′-Ribonucleotide
Having discarded a differential interaction of MmPrimPol and HsPrimPol with ssDNA and the 3′-site dNTP, the robust primase activity displayed by MmPrimPol is still not understood. This led us to investigate whether this robustness relays on the use of the 5′-site nucleotide during primer synthesis initiation. To explore this alternative, we performed DNA primase assays with the template oligonucleotide 3′ T_10_GTCAT_15_ 5′ to limit the synthesis to a _3p_AG dimer. Reactions were set up with a low concentration (16 nM) of the 3′ site nucleotide [α-^32^P]dGTP and increasing doses of the initiating 5′ site nucleotide, ATP. Our findings revealed that the potent primase activity of MmPrimPol is likely driven by an exceptionally high affinity for the 5′-nt. Remarkably, MmPrimPol synthesized much higher amounts of dimer than HsPrimPol, requiring approximately 500–1000-fold lower ATP concentrations to produce the same amounts of dimers (Figure 7c, compare lanes 6 and 7). This new experiment confirmed the critical role of the ZnFD of MmPrimPol in dimer formation, as a 500- to 1000-fold higher concentration of ATP was required to produce similar dimers when the ZnFD of MmPrimPol was deleted (compare lanes 7, 16, and 21), suggesting that the ZnFD is directly contributing to the strong interaction with the initiating ATP. Strikingly, both the Mm∆ZnFD deletion mutants displayed a drastically reduced primase activity, but comparable to the wild-type HsPrimPol (Figure 7c, compare lanes 5, 18, and 21), thereby emphasizing the enormous difference in primase activity of these two wild-type (mouse and human) PrimPols.
2.7. The Zn-Finger Domain of the Mouse PrimPol Likely Interacts with the β- and γ-Phosphate Moiety of the 5′-Initiating Ribonucleotide
As reported in Martínez-Jiménez et al., 2018 [16], the ZnFD of HsPrimPol contributes to primer synthesis through the interaction with the γ-phosphate moiety of the 5′-nucleotide, which is critical during dimer formation as well as during primer maturation. To determine whether the murine ZnFD confers enhanced primase activity via a stronger interaction with the γ-phosphate moiety, we assessed dimer formation using mono-, di-, and triphosphate versions of the initiating 5′-nucleotide (ATP, ADP, or AMP). These assays were performed in the presence of a 3′GTCA template and [α-^32^P]dGTP as the 3′-site nucleotide. As depicted in Figure 8a,b, the formation of the _3p_AG dimers by both the human and the mouse PrimPols was maximal with ATP (lanes 4 and 7), consistent with the importance of the γ-phosphate moiety described by Martínez-Jiménez et al. (2018) [16]. Interestingly, while HsPrimPol was just slightly affected by the presence of β-phosphate in the initiating 5′-nucleotide (compare lanes 2 and 3), dimer formation by MmPrimPol was enhanced more than 8-fold when using ADP instead of AMP (compare lanes 5 and 6), being more efficient for making the _2p_AG* dimers (lane 6) than HsPrimPol for making the _3p_AG dimers (lane 4). This result highlights that MmPrimPol exhibits a very strong selectivity for the β- and γ-phosphate moieties of the incoming 5′-nt, as the dimer formation was enhanced 27.5-fold when using ATP instead of AMP. By contrast, HsPrimPol showed only an 11.2-fold advantage with the presence of these two additional phosphates (Figure 8b).
Also consistent with Martínez-Jiménez et al. (2018) [16], our data demonstrate that the interaction of MmPrimPol with the β- and γ-phosphates of the initiating nucleotide is dependent on the ZnFD. As shown in Figure 8a (lanes 8 to 13), both the Mm∆ZnFD deletion mutants gain no advantage of the presence of a β- or γ-phosphate moiety during dimer synthesis. Beyond losing the benefit of using an ATP for primer initiation, the absence of the Zn-finger domain in MmPrimPol actually facilitated the use of AMP during primer synthesis. As shown in Figure 8a,b, both the MmZnFD mutants exhibited a 2.5- to 6-fold increase capacity to use AMP as the 5′-nt compared to the WT MmPrimPol (compare lane 5 to lanes 8 and 11). This could indicate that the three phosphates of ATP could be relevant ligands for MmPrimPol, but the lack of the ZnFD improves the stabilization of the α-phosphate moiety of AMP.
Collectively, our enzymatic characterization of MmPrimPol suggests that this enzyme is optimized for primer synthesis to a greater extent than its human ortholog. This optimization is attributable to the Mm∆ZnFD, likely providing a stronger interaction with the initiating 5′-ribonucleoside triphosphate during primer synthesis.
2.8. Deletion of HsPrimPol Flexible Region (∆231–246) Alters Its DNA Polymerase-DNA Primase Balance
The significant difference in DNA primase activity between the mouse and human PrimPol, as well as their different DNA primase/DNA polymerase balance, suggests that there is an intrinsic structural reason behind this. The multiple amino acid sequence alignment of PrimPols from diverse organisms identified a variable region situated between the catalytic motifs B and C, which exhibits moderate conservation and varies in length, being approximately 50 amino acids longer in mammals compared to insects or plants [8]. Further alignments conducted in this study (Figure 9a), more specifically of PrimPols from mammals within the Euarchontoglires clade, revealed that only the Muridae family has lost an 11 + 7 amino acid sequence within this variable region, that corresponds to amino acids 230 to 250 in HsPrimPol (Figure 9a, indicated by red boxes). Interestingly, this variable region is part of a larger disordered region (residues 201 to 260) that could not be crystalized in HsPrimPol [26].
This variable region, and especially these 18 amino acids that were lost in Muridae, could be related to the differences in the DNA primase/DNA polymerase balance displayed by the murine and human PrimPol. For this purpose, a deletion mutant of HsPrimPol lacking 16 amino acids from the region of interest (Hs∆16^(∆231–246)^) was generated, thereby rendering its length more similar to that of MmPrimPol (Figure 9b). We first evaluated the initiation of primer synthesis (dimer formation) of the Hs∆16 mutant by providing the GTCA oligonucleotide as a template, [α-^32^P]dGTP as the 3′-site nucleotide, and increasing concentrations of ATP as the preferred 5′-site initiating nucleotide. Interestingly, formation of the initiating dimers by Hs∆16 was slightly higher in comparison to HsPrimPol, but much lower than MmPrimPol (Figure 10a). Additionally, we explored the preference of Hs∆16 for the number of phosphates present in the initiating 5′-nucleotide. Hs∆16 was slightly better than HsPrimPol with any of the alternative nucleotides provided (AMP, ADP, or ATP), but the preference for nucleoside triphosphate (ATP) remained unaltered in the Hs∆16 mutant, and comparable in extent to HsPrimPol (Figure 10b, compare lanes 2–4 vs. 5–7). These results indicated that the elimination of the 16 amino acids sequence in HsPrimPol slightly improved its DNA primase activity, but did not reach the efficiency of MmPrimPol (Figure 10a,b, lanes 8–10, and Figure 10d).
Next, we evaluated the DNA polymerase activity of the Hs∆16 mutant by providing a pre-synthesized primer hybridized to a template with increasing concentrations of the four dNTPs. Interestingly, the primer extension activity of Hs∆16 was reduced when compared to HsPrimPol (Figure 10c, compare lanes 2–4 vs. 5–7); nevertheless, it was significantly more active than MmPrimPol (Figure 10c, lanes 8–10, and Figure 10d). We further asked if the reduced polymerization of the Hs∆16 mutant would affect the elongation step during primer synthesis. By using the 3′ GTC-step 5′ as a template, [γ-^32^P]ATP as the 5′-initiating nucleotide, and dGTP, dTTP, and dCTP as the 3′-site elongating nucleotides, the mutant Hs∆16 was able to initiate and extend de novo primers as well as HsPrimPol (Figure 10e). In conclusion, deletion of the 16 amino acids (residues 231 to 246) within the disordered region (residues 201 to 260) of HsPrimPol causes a moderate alteration in the DNA primase/DNA polymerase balance, that points to a slight “murinization” of this deletion variant.
3. Discussion
This work presents the enzymatic characterization of MmPrimPol, which revealed a much higher DNA primase activity compared to the previously described HsPrimPol [8]. We show here that the robust capacity of MmPrimPol to initiate DNA primer synthesis relies, for the most part, on a strong interaction with the β- and γ-phosphate moiety of the 5′-initiating ribonucleotide, which requires the ZnFD. Based on this study with MmPrimPol, we can predict that the strength of interaction with the 5′-nucleotide is the main factor that will define differences in the priming efficiency of PrimPols, a prediction also extrapolatable to more conventional primases. On the other hand, MmPrimPol and HsPrimPol displayed similar efficiencies in other enzymatic parameters, such as ssDNA binding, template sequence recognition, and binding of 3′-elongating nucleotides in the context of priming. Paradoxically, the MmPrimPol performance as a conventional DNA polymerase (extension of a pre-existing DNA primer) was very poor, when compared to HsPrimPol. That inverted DNA primase/DNA polymerase balance in MmPrimPol relative to HsPrimPol is explained by a special difficulty of MmPrimPol in accommodating a template/primer at the “initiation site”, likely due to physical interference with a constitutively closed ZnFD, which promotes a primase-ready configuration. Hence, MmPrimPol is fully devoted to DNA priming.
As previously shown, while the integrity of the Zn-finger is dispensable for the DNA polymerase activity of HsPrimPol, the Zn finger was essential for priming [9,15,16]; importantly, not only deletion of the entire ZnFD affected primer synthesis by PrimPol, but point mutations targeting Zn-ligands completely disrupted the primase activity of HsPrimPol, and both type of mutants failed to rescue stalled forks in human cells [9]. The relevance of the ZnFD for priming is reinforced here, by demonstrating that its elimination in MmPrimPol abolishes its robust DNA primase activity, but allows a better performance in conventional DNA polymerase assays. Therefore, these results also confirm that the Zn-finger domain modulates/inhibits the PrimPol DNA polymerase activity.
However, our findings contradict a previous report claiming that the Zn-finger is not essential for priming by MmPrimPol [32]. It is worth mentioning that these authors used a fluorescein-labeled nucleotide (FAM-ATP) for the initiating 5′ site. We hypothesize that the bulky nature of FAM-ATP could favor its binding at the defective 5′site in the MmΔZnFD. If that artificial stabilization enables some catalysis, it explains the misleading conclusion of a dispensable ZnFD in MmPrimPol. Surprisingly, Li et al. (2022) [32] did not include a comparison with the wild-type MmPrimPol, that would be very useful to know if (1) the primase activity observed with FAM-ATP in the absence of the ZnFD was residual in comparison with the robust primase activity shown here for the wild-type MmPrimPol, or (2) the wild-type MmPrimPol has problems accommodating the fluorescent FAM-ATP at the 5′ site due to steric hindrance with the ZnFD, impeding its closed conformation with the AEP (primase configuration). Considering these points, and supported by our previous and current experimental data, we strongly conclude that the ZnDF of MmPrimPol (and that of other PrimPols) is essential for its primase activity.
Another factor that may contribute to the robust primase activity of MmPrimPol could reside in the variable and disordered region located between catalytic motifs B and C (amino acids 201–242). In HsPrimPol, the disordered region corresponds to amino acids 201 to 260, and includes 18 extra amino acid residues that are absent in MmPrimPol. It is likely that this disordered region [26] may function as a ‘flexible’ linker that connects the catalytic core with the ZnFD, allowing different configurations to modulate primase and polymerase function. As shown here, elimination of the 16 amino acids of this hypothetical linker in HsPrimPol (mimicking MmPrimPol), moderately affected the DNA primase/DNA polymerase balance, slightly improving the primase but inhibiting the conventional DNA polymerase activity of HsPrimPol.
3.1. Initial Closing and Progressive Mobilization of the ZnFD During DNA Primer Synthesis: A Model
So far, our knowledge about PrimPol’s ZnFD function relies basically on in vitro and in vivo analysis of the ZnFD deficient mutants [9,15,16], but structural information about this domain remains unknown, as the current crystal structure of HsPrimPol lacks the ZnFD [26]. Previous structural analysis of the human primosome revealed that the C-terminal domain of the non-catalytic p58 subunit participates in the binding and stabilization of the template and the initiating NTP [33,34]. Here we showed that the C-terminal ZnFD of the monomeric MmPrimPol is also involved in the interaction with the initiating NTP, thus mimicking the function of p58; on the other hand, the catalytic core of PrimPol (as the catalytic subunit p48 of the dimeric primase) would be in charge of inserting 3′-nucleotides (dNTPs in the case of PrimPol) during both the initiation and elongation stages of primer synthesis, as it was also predicted for HsPrimPol [16,35].
The enzymatic characterization of HsPrimPol as both a DNA primase and a DNA polymerase supports a model in which PrimPol can assume two different configurations, based on the mobilization of the ZnFD, which favor either de novo primer synthesis or conventional polymerization (Figure 11a); the priming configuration is a tight (or closed) configuration that brings close together the catalytic core, responsible for binding the ssDNA template and 3′dNTP, and the ZnFD, which is crucial for binding the 5′NTP. Thus the two domains collaborate to complete the initiating quaternary complex, followed by dimer formation and its further extension to build a mature primer; alternatively, the polymerase configuration is a loose (or open) form of PrimPol, where the catalytic core operates independently of the ZnFD, that has to be mobilized to allow the accommodation of a primer–template substrate close to the catalytic site and the incoming 3′dNTP (ternary complex) to sustain normal (DNA polymerase-like) polymerization of dNTPs onto the primer strand.
The enzymatic characterization of MmPrimPol reported here supports that the murine PrimPol has a more prevalent priming configuration, that explains its robust DNA primase activity, in detriment to polymerization (Figure 11b). Thus, MmPrimPol seems to be pre-configured to start primer synthesis by assuming a constitutive and tightly closed priming configuration likely due to a more defined and closed positioning of its ZnFD. Importantly, the elongation phase of primer synthesis, that requires progressive mobilization of the ZnFD is not compromised in MmPrimPol. In agreement with these conclusions, an effective DNA polymerase activity was unmasked in MmPrimPol when the ZnFD was eliminated, leaving the protein in an artificially opened configuration that licenses polymerization (Figure 11c).
Future crystal structures of the entire PrimPol in priming configuration will be of great value to describe how the ZnFD contributes to stabilize the initiating 5′-nucleotide, which specific residues are involved in the interaction with its sugar and phosphate residues, and how these interactions are modified or remain during processive elongation to generate a mature primer. The robust DNA primase activity of MmPrimPol, and in particular its constitutive priming configuration, with a less mobile ZnFD, make MmPrimPol an ideal candidate for crystallization in primase mode.
3.2. Mouse PrimPol: Need for a Strong Primase?
The significant differences in the specific activity of the human and mouse PrimPols raise the question of why mice have evolved such a potent DNA primase. A brief look into these two species gives a clue to explain the need for a robust priming by MmPrimPol. Some physiological characteristics of humans and mice are very contrasting, such as body weight and size, thermoregulation, lifespan, reproductive rate, aging processes, and rapid cellular turnover, just to list a few. Many of these features have close connections to metabolism, which directly correlates with mitochondrial function [36]. Thus, preserving the integrity of the mitochondria is very important to keep the homeostasis in the physiology of the entire organism. In fact, the metabolic rates between these two species vary enormously, with the level of oxidative stress and damage being higher in mice.
It is known that the lack of PrimPol in mice results in a slight shortening of lifespan, increased incidence of late-onset tumor development, higher sensibility to UV-light injury, and impaired mitochondrial function (unpublished data). In this context, PrimPol has been shown to contribute to the maintenance of both nuclear and mitochondrial genomes through its repriming activity, that allows the completion of DNA replication under many different stress conditions [9,24,25,28]. Thus, it is conceivable that selection of a murine PrimPol with a robust primase activity could be an adaptive response to its high metabolic rate, elevated ROS production, and rapid cellular turnover. So, we speculate that the potent primase activity of MmPrimPol would meet the need to restart stalled forks in a more demanding cellular environment.
4. Materials and Methods
4.1. Reagents
Inorganic salts, acids, bases, and organic compounds were purchased from Merck (Darmstadt, Germany), Sigma (St. Louis, MO, USA), and AppliChem (Darmstadt, Germany). The chromatography resins for protein purification used in this work were Ni-NTA Superflow purchased from Qiagen (Hilden, Germany) and Heparin Sepharose Fast Flow purchased from GE Healthcare (Fairfield, CT, USA).
Unlabeled ultrapure AMP, ADP, NTPs, and dNTPs were supplied by GE Healthcare (Fairfield, CT, USA). Labeled nucleotides, [γ-^32^P]ATP and [α-^32^P]dGTP, were purchased from Revvity (Waltham, MA, USA). T4 polynucleotide kinase, T4 DNA Ligase, and DNA restriction endonucleases were obtained from New England Biolabs (Ipswich, MA, USA) and Roche (Basel, Switzerland). Vent polymerase was supplied by New England Biolabs (Ipswich, MA, USA).
4.2. Oligonucleotides
DNA oligonucleotides were synthesized by Sigma Aldrich (St. Louis, MO, USA) and purified by 8 M urea-20% polyacrylamide gel electrophoresis.
4.3. Cloning the Mouse PrimPol and Mutant Generation
MmPrimPol is codified by the CCDC111 gene, with the same name as its human ortholog, and it is located in chromosome 8 (8 B1.1). A full-length cDNA of the CCDC111 mouse gene, cloned in the pET28/murineCCDC111 vector was kindly given by Dr. Raimundo Freire (Hospital Universitario La Laguna, Canary Islands, Spain). The CCDC111 gene was amplified by PCR using oligos MmWT-forward (5′-ACGATCATATGTTGAGGAAATGGGAGGCAAG-3′) and MmWT-reverse (5′-ACGATGGATCCTCAGCTATTCTGTAAAGCTTCTATA-3′), that introduce the NdeI-BamHI sites, respectively, then PCR products were digested and introduced in pET16b expression vector (Novagen, Darmstadt, Germany) cut with the same restriction enzymes, as already described for HsPrimPol [8]. The resulting plasmid was named pET16/murineCCDC111 and kept in an E. coli DH5α strain. Two deletion mutants of MmPrimPol lacking the C-terminal domain including the ZFD were generated, named Mm-∆ZnFD-1^(1–391)^ and Mm-∆ZnFD-2^(1–336)^. The deletion mutants were constructed by PCR amplification of the first 1173 nucleotides (Mm-∆ZnFD-1) or 1008-nt (Mm-∆ZnFD-2) of the mouse CCDC111 cDNA using primers *Mm-*WT-forward combined with Mm-∆ZnFD-1-reverse (5′-ACGATGGATCCTCACAGTAATTCTTCCGGGAAAAAG-3′) or Mm-∆ZnFD-2-reverse (5′-ACGATGGATCCTCACTTTCGTTTAGTCTGAGATGGG-3′). The amplified products were inserted in E. coli expression plasmid pET16 (Novagen) in NdeI-BamHI sites and further sequenced to confirm the deletion version of MmPrimPol.
4.4. Purification of the Human and Mouse PrimPol Variants
Protein expression and purification was performed identically for the human and mouse PrimPol, both the wild-type and the mutant variants. The protocol for purification was as described previously [8].
4.5. Polymerase Assay on Specific Primer: Template Molecules
DNA polymerase assays were carried out with labeling at the 5′ the primer using PNK and [γ-^32^P]ATP (250 µCi, 3000 Ci/mmol), then hybridized to a complementary template in a buffer containing 0.3 M NaCl, 50 mM Tris-HCl pH 7.5. DNA sequences are indicated in the figures. Reaction mixtures (in 20 µL) contained Buffer R [50 mM Tris–HCl pH 7.5, 40 mM NaCl, 2.5% (w/v) glycerol, 1 mM DTT, 0.1 mg/mL BSA, 1 mM MnCl_2_, or 5 mM MgCl_2_], 2.5 nM [γ-^32^P]-labeled primer–template, 200 nM purified PrimPol variant, and dNTPs or NTPs as indicated in each figure. Polymerase assays were incubated 20 min at 30 °C, and stopped by adding 8 μL of formamide loading buffer (95% formamide, 20 mM EDTA, 0.1% xylene-cyanol, and 0.1% bromophenol blue), then loaded onto an 8 M urea-containing 20% polyacrylamide sequencing gel. Primer extension was detected by autoradiography and the images analyzed with ImageJ software 1.49v.
4.6. Primase Assay on Specific Oligonucleotide Templates
Primase assays were carried out using the indicated ssDNA oligonucleotide, containing the priming site 3′GTC 5′ flanked by thymidine tracts or variations around this sequence. The reaction mixture (20 µL) contained Buffer R, 1 µM ssDNA template, 400 nM PrimPol, 16 nM [γ-^32^P]ATP or [α-^32^P]dGTP (250 µCi, 3000 Ci/mmol) and the nucleotides indicated in each figure at the described concentrations. After incubation for 30 min at 30 °C, reactions were stopped by adding 8 μL of formamide loading buffer (95% formamide, 20 mM EDTA, 0.1% xylene-cyanol, and 0.1% bromophenol blue) and the products resolved in an 8 M urea-containing 20% polyacrylamide sequencing gel (50 cm long) that was run at 50 W for 2 h. Following electrophoresis, de novo synthesized primers were detected by autoradiography and the images were analyzed with ImageJ software 1.49v.
4.7. EMSA for PrimPol/ssDNA Binary Complex
The PrimPol/ssDNA binding assay was performed with [γ-^32^P]-labeled ssDNA oligonucleotide (3′T_20_-GTCAGACAGCA-T_29_ 5′). The reaction mixtures (20 µL) contained Buffer R, 2.5% (w/v) PEG-4000 and purified PrimPol in the concentrations indicated in the figure. Samples were pre-incubated for 10 min at 4 °C and then incubated for 10 min at 30 °C with 2.5 nM 5′-labeled ssDNA. Reactions were stopped by adding 2 µL loading Buffer S (50% glycerol, 0.1% (w/v) xylene cyanol, and 0.1% (w/v) bromophenol blue), and analyzed in a native 6% polyacrylamide gel, resolved in Tris-glycine buffer (pH 8.3) at 180 V for 120 min at 4 °C. After electrophoresis, gels were vacuum-dried and mobility shift of free ssDNA versus enzyme/ssDNA complex was analyzed by autoradiography.
4.8. EMSA for PrimPol/ssDNA/dNTP Pre-Ternary Complex
Pre-ternary (PrimPol/ssDNA/dG) complex formation using PrimPol variants and [α-^32^P]dGTP (16 nM) were evaluated in Buffer D (50 mM Tris–HCl pH 7.5, 40 mM NaCl, 2.5% (w/v) glycerol, 1 mM DTT, 0.1 mg/mL BSA), supplemented with 0.5 µM ssDNA 3′ T_20_-GTCC-T_36_ 5′, and 1 mM MnCl_2_. Reactions (20 µL final volume) were incubated for 10 min at 30 °C. Then, 2 µL of loading Buffer S was added and the reactions were analyzed as described above.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Frick D.N. Richardson C.C. DNA Primases Annu. Rev. Biochem.200170398010.1146/annurev.biochem.70.1.3911395402 · doi ↗ · pubmed ↗
- 2Laosirieix S. Pellegrini L. Bell S. The Promiscuous Primase Trends Genet.20052156857210.1016/j.tig.2005.07.01016095750 · doi ↗ · pubmed ↗
- 3Kuchta R.D. Stengel G. Mechanism and Evolution of DNA Primases Biochim. Biophys. Acta BBA Proteins Proteom.201018041180118910.1016/j.bbapap.2009.06.01119540940 PMC 2846230 · doi ↗ · pubmed ↗
- 4Bergsch J. Allain F.H.-T. Lipps G. Recent Advances in Understanding Bacterial and Archaeoeukaryotic Primases Curr. Opin. Struct. Biol.20195915916710.1016/j.sbi.2019.08.00431585372 · doi ↗ · pubmed ↗
- 5Stodola J.L. Burgers P.M. Mechanism of Lagging-Strand DNA Replication in Eukaryotes DNA Replication Advances in Experimental Medicine and, Biology Masai H. Foiani M. Springer Singapore Singapore 2017 Volume 104211713310.1007/978-981-10-6955-0_629357056 · doi ↗ · pubmed ↗
- 6Baranovskiy A. Tahirov T. Elaborated Action of the Human Primosome Genes 201786210.3390/genes 802006228208743 PMC 5333051 · doi ↗ · pubmed ↗
- 7Iyer L.M. Origin and Evolution of the Archaeo-Eukaryotic Primase Superfamily and Related Palm-Domain Proteins: Structural Insights and New Members Nucleic Acids Res.2005333875389610.1093/nar/gki 70216027112 PMC 1176014 · doi ↗ · pubmed ↗
- 8García-Gómez S. Reyes A. Martínez-Jiménez M.I. Chocrón E.S. Mourón S. Terrados G. Powell C. Salido E. Méndez J. Holt I.J. Prim Pol, an Archaic Primase/Polymerase Operating in Human Cells Mol. Cell 20135254155310.1016/j.molcel.2013.09.02524207056 PMC 3899013 · doi ↗ · pubmed ↗