Genome-Wide Identification and Comprehensive Analysis of Ubiquitin-Specific Protease Gene Family in Soybean (Glycine max)
Cuirong Tan, Dingyue Ban, Haiyang Li, Jinxing Wang, Baohui Liu, Chunyu Zhang

TL;DR
This study identifies 52 UBP genes in soybean and explores their role in seed development and traits like weight and nutrient content.
Contribution
The first comprehensive analysis of the UBP gene family in soybean, revealing their evolutionary patterns and functional diversity.
Findings
52 UBP genes were identified in soybean, grouped into 14 phylogenetic clusters with conserved domains.
GmUBP7 shows significant haplotype variation linked to seed weight, fatty acid, and protein content in natural soybean varieties.
Gene duplication and synteny suggest a close evolutionary relationship between soybean and Arabidopsis UBP genes.
Abstract
Deubiquitination plays a pivotal role in regulating plant responses to abiotic stress, growth, and development. Among the deubiquitinase (DUB) families, ubiquitin-specific proteases (UBPs) constitute the largest group. Despite this, limited research has been conducted on the functional characteristics of the UBP gene family in soybean (Glycine max). In this study, we identified 52 UBP gene family members in soybean, all of which harbored UCH (ubiquitin C-terminal hydrolase) domains with short yet evolutionarily conserved Cys-box and His-box. These genes were phylogenetically classified into 14 distinct groups; GmUBP genes within the same group shared analogous patterns of conserved domains and motifs. Moreover, a synteny analysis reveals that the GmUBP family has undergone extensive gene duplication events and shares a close evolutionary relationship with Arabidopsis thaliana. We…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1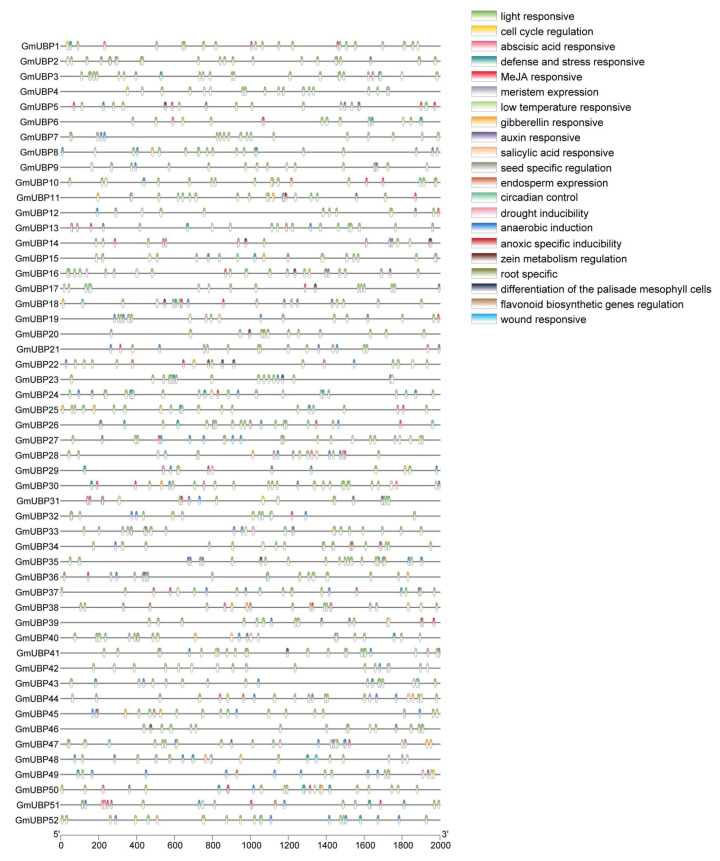 Figure 2
Figure 2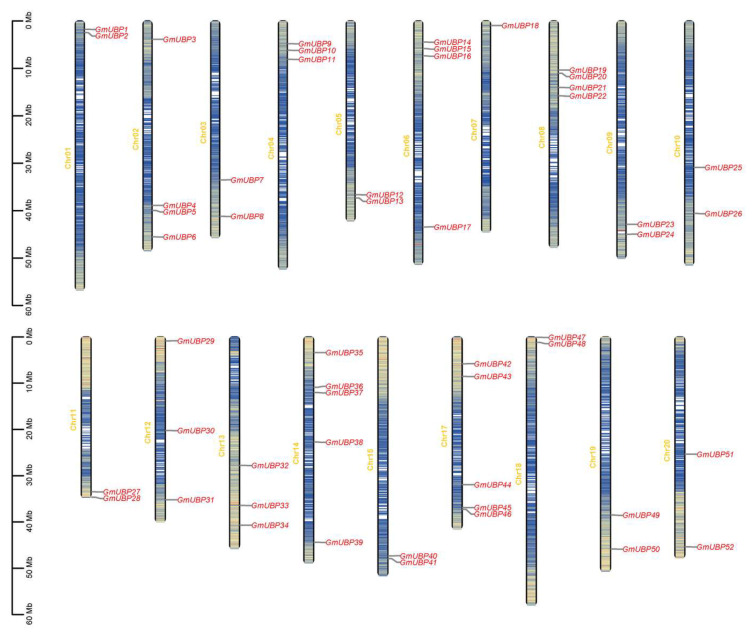 Figure 3
Figure 3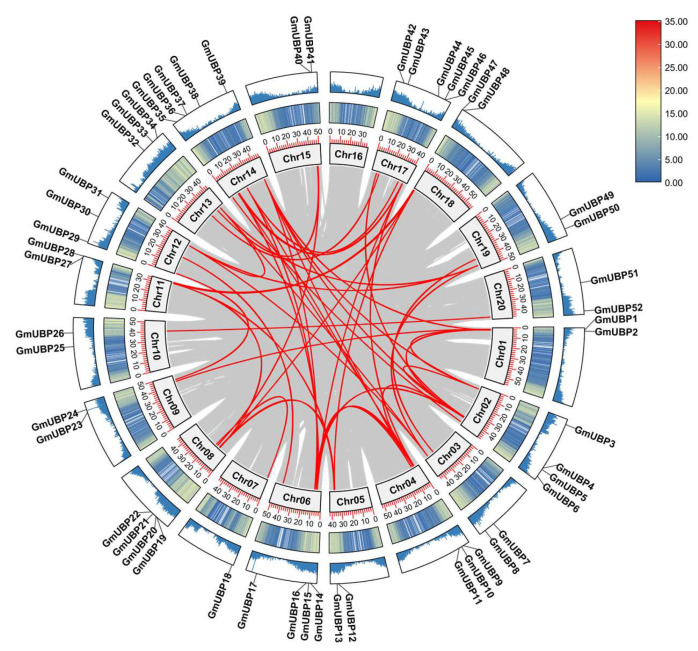 Figure 4
Figure 4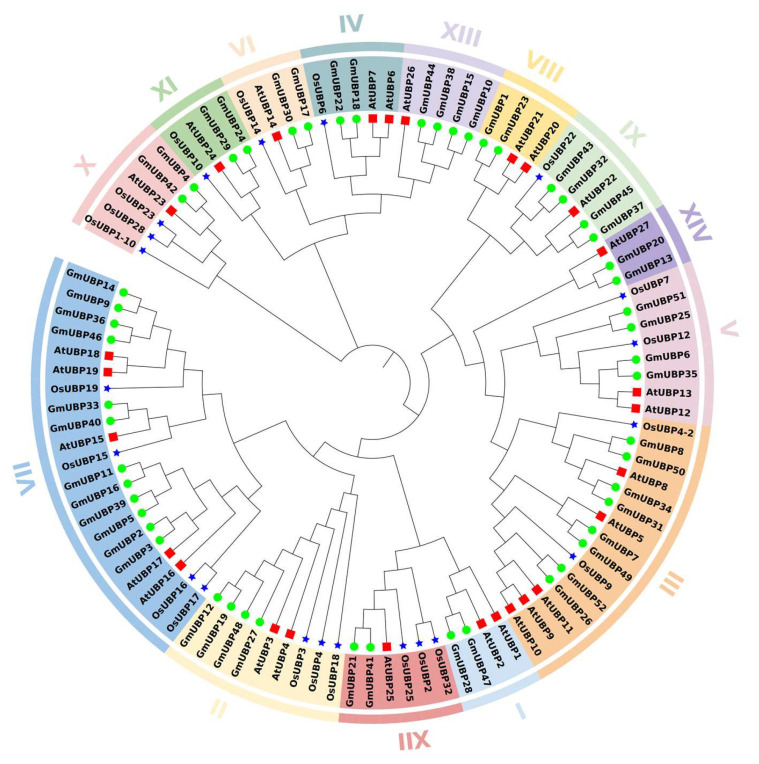 Figure 5
Figure 5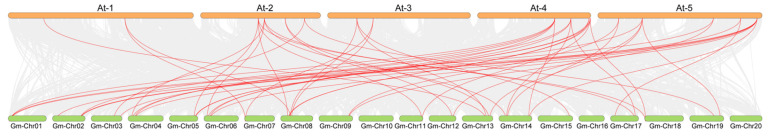 Figure 6
Figure 6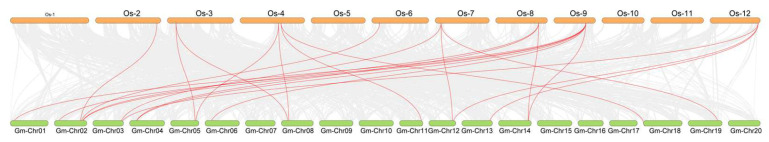 Figure 7
Figure 7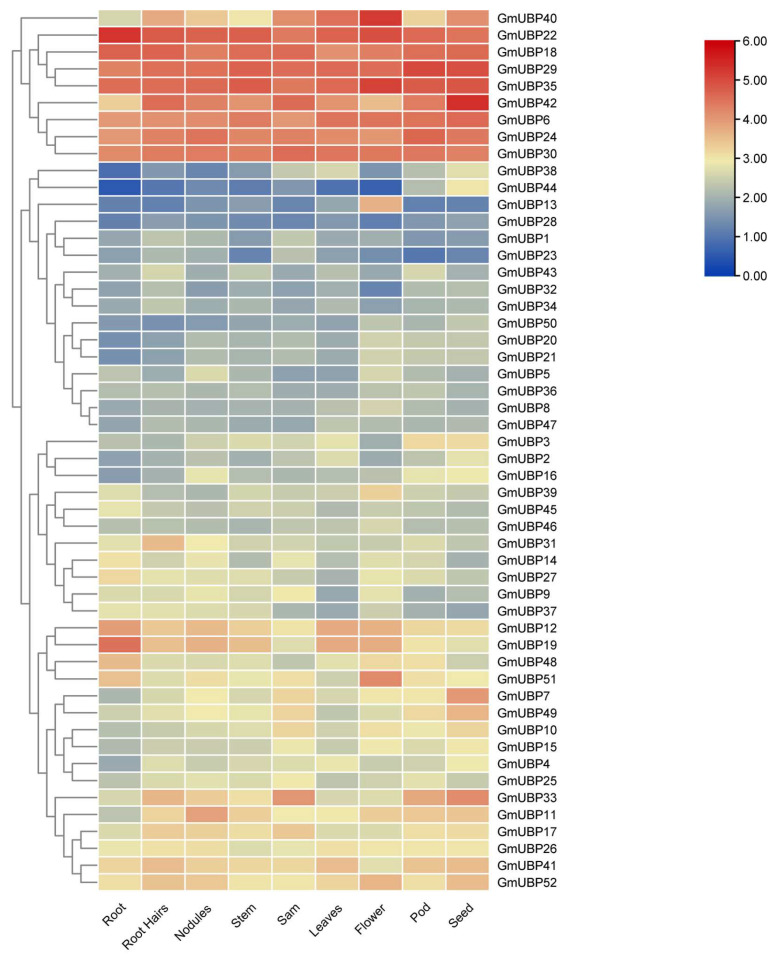 Figure 8
Figure 8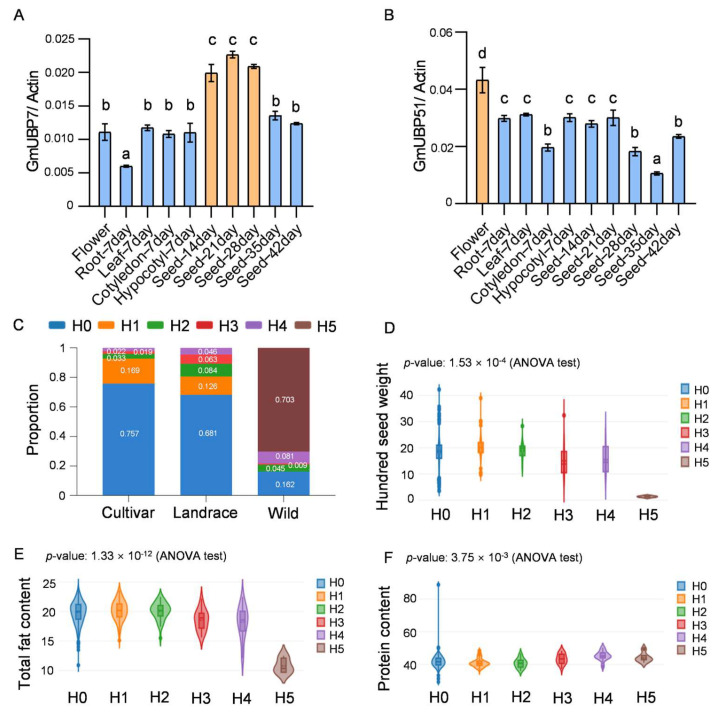 Figure 9
Figure 9- —National Natural Science Foundation of China
- —Guangdong Basic and Applied Basic Research Foundation
- —Science and Technology Innovation Project of the Suihua Branch, Heilongjiang Academy of Agricultural Sciences
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSoybean genetics and cultivation · Plant nutrient uptake and metabolism · Photosynthetic Processes and Mechanisms
1. Introduction
Soybean (Glycine max (L.) Merr.) is a globally significant crop that is utilized as an oil, feed, and forage source; it is a major provider of edible oil and plant protein for humans and livestock worldwide [1]. Yield and quality in soybean plants are closely tied to seed development, which is regulated by multiple signaling pathways, including the ubiquitin–proteasome pathway, the G-protein signaling pathway, the mitogen-activated protein kinase (MAPK) signaling pathway, plant hormone pathways, the HAIKU (IKU) pathway, and transcription regulatory factors [2]. Among these pathways, ubiquitination stands out as a critical process, influencing plant growth and development by regulating the activity and stability of target proteins [3].
Protein ubiquitination and deubiquitination are well-established fields of study [4,5]. The ubiquitination process involves three sequential steps and is mediated by a cascade of the following three enzymes: the E1 ubiquitin-activating enzyme, the E2 ubiquitin-conjugating enzyme, and the E3 ubiquitin ligase [3,6]. Deubiquitination, which is the opposite process of ubiquitination, involves deubiquitinating enzymes (DUBs) and removing ubiquitin (Ub) from ubiquitinated proteins in order to stabilize them [7]. Notably, in plants, ubiquitin-specific proteases (UBPs) form the largest and most diverse subfamily of DUBs. UBPs are involved in two primary biochemical activities: cleaving linked ubiquitin chains from proteins conjugated with ubiquitin and producing mature ubiquitin from its precursor [3]. All UBPs possess a ubiquitin C-terminal hydrolase (UCH) domain—this is unique to UBP proteins and is known as the UBP domain—along with two short but highly conserved motifs: the cysteine (Cys) box and the histidine (His) box. These motifs contain key catalytic residues—the cysteine in the Cys-box and histidine and aspartic acid/asparagine in the His-box—which are essential for the deubiquitination activity of UBPs [8,9].
The ubiquitin-specific peptidase (UBP) family has been characterized in various plant species, exhibiting marked interspecific differences in gene numbers. Specifically, genome-wide analyses have revealed 27 UBP genes in Arabidopsis thaliana [10], 25 in Oryza sativa [11], 48 in Phyllostachys edulis [12], 97 in Triticum aestivum [13], and 45 in Zea mays [14]. These enzymes play critical roles in diverse stress response pathways, including abscisic acid (ABA)-mediated drought tolerance, salt stress adaptation, and immune responses, as well as other key biological processes [15,16,17,18,19]. In Arabidopsis, the overexpression of UBP12/UBP13 enhances ORESARA1 (ORE1) protein levels, thereby promoting nitrogen-deficiency-induced leaf senescence [20]. Additionally, UBP12 and UBP13 regulate plant growth responses under nitrogen starvation and post-carbon starvation recovery by stabilizing the BRI1 EMS SUPPRESSOR 1 (BES1) transcription factor. Furthermore, these enzymes counteract root meristem growth factor 1 (RGF1)-induced ubiquitination by interacting directly with RGF1 receptors, thereby promoting root meristem development [21]. Interestingly, UBP12 and UBP13 also contribute to the regulation of flowering time and circadian rhythms [22]. Loss-of-function mutations in AtUBP1 and AtUBP2 result in hypersensitivity to the amino acid analog canavanine (CAN), accompanied by severe developmental defects, including dwarfism, short root growth, and chlorosis [10]. During reproductive growth, UBP15 has been shown to regulate seed development in both Arabidopsis and rice. It antagonizes DA1-mediated ubiquitin receptor signaling to modulate organ development and seed size while promoting maternal bead tissue proliferation to enhance seed size [23]. In rice, OsUBP15 interacts directly with OsDA1 to regulate grain dimensions [24]. Other UBP gene family members also contribute uniquely to plant reproduction, e.g., AtUBP3 and AtUBP4 are essential for male gametophyte development [25], AtUBP14 participates in early embryogenesis [26], and AtUBP26 plays a role in histone ubiquitination and seed development [27,28]. Moreover, OsUBP2 has been identified as a negative regulator of reactive oxygen species (ROS) accumulation and immune responses in rice [19]. Subsequent studies have demonstrated that OsUBP2 modulates rice cell death and immune responses by stabilizing the SPOTTED LEAF 35 (SPL35) protein [29]. Furthermore, this protein plays a role in rice immunity by deubiquitinating H2B; this is a process that regulates genes in the salicylic acid (SA) biosynthesis pathway [30]. In addition, rice seedlings with a homozygous T-DNA insertion mutant in OsUBP6 exhibit defective phenotypes during early growth, though these defects diminish as the plants mature [31]. In maize, the homologs of AtUBP16, namely, ZmUBP15, ZmUBP16, and ZmUBP19, are essential for the plant’s response to salt and cadmium stress [32].
In soybean, the UBP gene family remains poorly characterized compared to other plant species. To date, only one GmUBP gene has been comprehensively studied, encoding a ubiquitin-specific protease orthologous to AtUBP22. This gene, designated as GmSW17 (Seed Width 17), has been shown to determine seed width and weight in natural soybean populations. Population genomics analyses reveal that GmSW17 underwent artificial selection during soybean domestication but has not been fully fixed in modern cultivars [33]. To investigate the roles of soybean UBP genes in plant growth and development, we systematically identified 52 GmUBP genes across the soybean genome. Subsequent analyses focused on their conserved motifs, gene structures, chromosomal distributions, and expression patterns. To elucidate their evolutionary relationships with UBP genes in other plant species, phylogenetic tree construction and collinearity analysis were performed. Additionally, we conducted an in-depth investigation of GmUBP7, analyzing its expression levels across multiple stages of soybean seed development using RT-qPCR. Haplotype analysis further reveals significant differences in 100-seed weight, total fatty acid content, and protein content among distinct haplotypes. The findings of this study contribute to a better understanding of the functional roles of GmUBP genes in seed development and provide a foundation for identifying the roles of this essential gene family in soybean agronomics and improvement.
2. Results
2.1. Identification and Characterization of the UBP Genes in Soybean
To identify members of the UBP gene family in soybean, we initially queried the relevant databases using 27 Arabidopsis UBP protein sequences. This analysis yielded 63 putative GmUBP genes, which were further validated by examining their conserved UCH domain using the Pfam database and NCBI CD Search. After removing non-conforming sequences, 52 GmUBP genes were identified in the soybean genome and were systematically named GmUBP1 to GmUBP52 based on their chromosomal locations (Table S1). All 52 GmUBPs contained the UCH domain.
Detailed information about these genes, including gene length, physical and chemical properties, transmembrane domains, and subcellular localization, is presented in Table S1. The coding sequences of the GmUBP genes ranged from 1107 to 3423 bp, while their protein lengths spanned 369–1141 amino acids. The molecular weight (MW) of these proteins varied between 42.052 and 133.168 kDa, with isoelectric points (pIs) ranging from 4.96 to 9.2. Predictions for transmembrane domains reveal that 11 GmUBPs contained a single TM domain (e.g., GmUBP2). Subcellular localization analyses provided insights into the functional roles of these proteins. The results indicate that 24 GmUBPs were localized in the nucleus, while 13 were found in the plasma membrane. Additionally, seven GmUBPs were associated with the endomembrane system, and four were localized in chloroplasts. Notably, GmUBP5 (Glyma.02G213400) and GmUBP11 (Glyma.04G091700) were predicted to reside in the extracellular space, whereas GmUBP24 (Glyma.09G225200) and GmUBP29 (Glyma.12G011800) were localized in organelle membranes. Finally, all 52 GmUBPs exhibited hydrophilicity, as their predicted hydrophobicity scores were less than zero (Table S1).
2.2. Gene Structures, Conserved Motifs, and cis-Acting Element Analysis of the GmUBP Genes
In order to elucidate the relationship between gene function and evolution, we analyzed the structural organization and conserved motifs of GmUBP genes (Table S2–S4). The GmUBP25 gene (V subfamily) had the largest number of exons (32), while GmUBP28 and GmUBP47 (I subfamily) had the smallest number of exons (2). Other GmUBP genes contained exon numbers ranging from 3 to 31. Genes within the same subfamily exhibited similar intron–exon structures, suggesting functional conservation within the soybean UBP gene family. The variability in exon numbers among GmUBP genes may indicate functional diversification within the family (Figure 1A,C).
To further investigate the evolutionary changes in the GmUBP family, we analyzed the conserved motifs in the 52 GmUBP proteins using MEME online software, identifying 10 distinct motifs (designated motifs 1–10). Motifs 1, 4, and 5 were consistently present in all GmUBP proteins, and their order (1, 4, and 5) was consistent except for GmUBP24 and GmUBP29. Additionally, motif 10 was primarily found in subfamily III, while motif 7 was exclusive to subfamily VII (Figure 1B, Table S5).
For promoter analysis, we directly extracted promoter sequences (2000 bp upstream of the start codon—ATG) for the 52 GmUBP genes from the soybean genome sequence. Using the PlantCare online tool, we predicted cis-acting elements, which were subsequently visualized using TBtools software. These elements included hormone-responsive motifs (e.g., ABA, IAA, GA, MeJA, and SA), environmental stress response elements (e.g., low temperature, light responses, and circadian rhythms), and anaerobic induction elements. Of these, light-responsive elements and anaerobic induction elements were the most prevalent (Figure 2).
2.3. Visualization of Chromosomal Location and Duplication of GmUBPs
To examine the chromosomal distribution of GmUBP genes, their chromosomal locations were determined. The results reveal that the 52 GmUBP genes were mapped to 20 chromosomes, with an uneven distribution across each chromosome (Figure 3). Specifically, chromosomes 14 and 17 each contained five GmUBP genes, while chromosomes 2, 6, and 8 each had four GmUBP genes. Chromosomes 4, 12, and 13 each harbored three GmUBP genes. Chromosomes 1, 3, 5, 9, 10, 11, 15, 18, 19, and 20 each carried two GmUBP genes. Notably, only one GmUBP gene was found on chromosome 7. Additionally, there was no apparent correlation between the number of GmUBP genes and the soybean chromosome length.
Furthermore, the gene duplication events of the GmUBP family within the soybean genome were analyzed (Figure 4). Gene duplication can occur through mechanisms such as tandem duplication and segmental duplication [34]. In total, 47 gene pairs undergoing segmental duplication were identified across 19 chromosomes (except for chr16), with no evidence of tandem duplications. This suggests that segmental duplications may play a dominant role in the expansion of GmUBP genes.
2.4. Phylogenetic Analysis and Collinearity Analysis of the UBP Genes Among Arabidopsis, Soybean, and Rice
To investigate the phylogenetic organization of the UBP family, we conducted a comprehensive phylogenetic analysis using the protein sequences of 52 soybean UBPs, 32 rice UBPs, and 27 Arabidopsis UBPs (Tables S6 and S7). The phylogenetic tree was constructed using the neighbor-joining (NJ) method, followed by Maximum Likelihood (ML) analysis. Based on their phylogenetic relationships, the UBPs were classified into 14 distinct groups, designated as I to XIV (Figure 5). Notably, Group VII exhibited the largest membership, comprising 12 GmUBP proteins, while Group III contained 8 GmUBP proteins, representing the second-largest group. Interestingly, many UBP genes in soybean exist as closely related pairs, such as GmUBP4 and GmUBP42, further supporting the presence of paralogous genes within the soybean UBP family.
However, no orthologous relationships were observed among the UBP genes of soybean, rice, and Arabidopsis, suggesting that these UBP genes have undergone functional divergence during evolution. Despite this divergence, UBP proteins from soybean and Arabidopsis were distributed across 15 phylogenetic groups, indicating a conserved functional diversity within these two species. Conversely, Groups I, VIII, XIII, and XIV lacked rice UBP representatives, highlighting evolutionary differences in the functional specialization of UBP genes between soybean and rice.
To elucidate the origin and evolutionary trajectory of the GmUBP gene family, we performed a comprehensive collinearity analysis by generating a collinearity map of Glycine max (soybean) against Arabidopsis thaliana and Oryza sativa (Figure 6). This analysis reveals 51 significant collinearity relationships between soybean and Arabidopsis genes, while only 23 such relationships were identified with rice orthologs. These findings suggest that the GmUBP gene family exhibits a closer evolutionary relationship with Arabidopsis, a conclusion that is strongly supported by phylogenetic tree analysis.
2.5. Expression Patterns of UBP Family in Different Tissues
To further investigate the biological functions of the UBP gene family in soybean, we analyzed the expression profiles of GmUBPs across various tissues, including the root, root hairs, nodule, stem, shoot apical meristem (SAM), leaves, flower, pod, and seed (Figure 7). The expression patterns of the 52 GmUBP genes were categorized into four distinct groups. The first group comprises nine genes, such as GmUBP40, which exhibits the highest expression levels across multiple tissues. The second group, consisting of 16 genes (e.g., GmUBP12 and GmUBP52), displays overall high expression levels, despite a relatively low expression in some tissues (Figure 7). Notably, GmUBP7, whose high expression in seeds is analogous to that of AtUBP14 (AT3G20630), is suggested to have an important role in seed development or maturation. The third group includes 16 genes (e.g., GmUBP38 and GmUBP47) characterized by low expression across most tissues, except for GmUBP13, which shows the highest expression in flowers. Among these, GmUBP31 is highly expressed in root hairs, while GmUBP39 is prominently expressed in flowers, indicating potential involvement in soybean flowering. Furthermore, GmUBP transcripts in pods and seeds generally demonstrated higher expression levels compared to other tissues, implying that GmUBP-mediated deubiquitination is predominantly utilized to regulate reproductive growth rather than vegetative development.
In addition, based on the results of online transcriptome data, we selected two GmUBP genes to further determine their expression patterns. GmUBP7 and GmUBP51 were expressed in ten tissues (flower and root—7 days, leaf—7 days, cotyledon—7 days, hypocotyl—7 days, seed—14 days, seed—21 days, seed—28 days, seed—35 days, and seed—42 days) with relative expression levels. The results show that the qRT-PCR expression patterns of the two genes were consistent with the general trends of transcriptome expression patterns. GmUBP7 is highly expressed in seeds, especially in the early and middle stages of seed development, while GmUPB51 is highly expressed in flowers (Figure 8A,B). However, while GmUBP51 expression was slightly higher in flowers than in other tissues, it was not as different as the transcriptome data showed, which may have been due to sample differences in sequencing and qRT-PCR.
2.6. Functional Analysis of the GmUBP Family Genes
To investigate the potential selection and biological functions of the soybean GmUBP gene family, we performed a haplotype analysis using the Soybean Multi-Omics Database. In a comprehensive analysis of 4414 re-sequenced soybean accessions, all 52 GmUBP genes exhibited distinct haplotypes. Of these, we found members in each subfamily that demonstrated evidence of selection across wild soybean, landraces, and cultivars. Therefore, we present the haplotypes of 14 members as being representative of the 14 groups. (Figure S1). This observation suggests that most of the GmUBP genes have undergone selection during soybean domestication, which may be attributed to either passive selection due to genetic linkage with other genes or active selection driven by advantageous phenotypes.
Furthermore, we speculated on the function of genes based on their specific expression in tissues. As flowering- and seed-related traits are important agrological traits of soybean, we focused on the genes GmUBP7 and GmUBP51, which are highly expressed in flowers and seeds in the transcriptome, and further verified them (Figure 7). In both genes, the qPCR results of GmUBP7 were consistent with the transcriptome, and both showed that the gene was highly expressed during the early and middle stages of seed development. However, the expression level of GmUBP51 in flowers was different from that of the transcriptome; therefore, we focused only on GmUBP7 for further haplotype analysis (Figure 8A,B). The haplotype analysis of this gene reveals notable shifts in the proportions of the H0 and H5 haplotypes across wild soybean, landraces, and cultivars. Specifically, the frequency of H0 increased during domestication, while the frequency of H5 decreased (Figure 8C, Table S8). The phenotypic correlation analysis of the five haplotypes with database phenotypic data demonstrates significant differences in the hundred-seed weight, total fat content, and protein content between H0–H4 and H5 (Figure 8D–F). In addition, H0 also appears to have been subjected to artificial selection, but its seed phenotype was not significantly different from that of H1–H4, probably because H0 is associated with other traits, such as seed coat thickness and germination rate. These findings suggest that GmUBP7 may have been subjected to selection during domestication. The high expression levels of GmUBP7 in seeds, as well as its association with seed traits, highlight its potential importance in soybean improvement programs.
3. Discussion
As an essential form of protein modification, ubiquitination significantly impacts various cellular processes, such as hormone signaling, stress responses, and organ development [13,23,24,26]. Deubiquitination, serving as a reversible counterpart to ubiquitination, involves the removal of ubiquitin from modified substrates by deubiquitinating enzymes (DUBs), thereby regulating the levels of ubiquitination on target proteins. USPs/UBPs, a major category of DUBs, form a diverse group of enzymes comprising numerous members. Compelling evidence demonstrates that distinct UBPs play varied and vital roles in plant development and stress responses [35,36,37,38,39]. The UBP gene family has been studied in several species, including Arabidopsis thaliana [10], Oryza sativa [11], Phyllostachys edulis [12], Triticum aestivum [13], and Zea mays [14]. However, research into the function of GmUBPs in soybean development remains underexplored. In this study, we conducted a thorough genome-wide analysis of soybean UBP genes utilizing publicly accessible genome data. Additionally, we examined the potential roles of UBP genes in soybean by analyzing genetic variation and phenotypic data.
In the present investigation, structural analysis reveals that all characterized GmUBP proteins harbored the UCH domain, which is characterized by the presence of conserved Cys and His-box motifs (Figure 1). Furthermore, a comparison of domain composition among different genetic clades indicates that members within more closely related branches tended to share identical structural motifs, whereas proteins from distinct clades exhibited variations in their domain composition (Figure 1). These findings suggest inherent differences in substrate affinity and functional roles among the GmUBP family members. Soybean, as a recognized paleopolyploid genome [40], has undergone a series of whole-genome duplication events, including the Gamma event (or WGT), which is estimated to have occurred prior to the divergence of monocots and dicots approximately 300 million years ago [41]. Additional polyploidization events, such as legume-wide genome duplication (WGD) around 59 million years ago and Glycine-specific WGD approximately 13 million years ago [42,43], have collectively contributed to the highly duplicated nature of the soybean genome. Currently, over 70% of soybean genes exist in multiple copies [44,45]. In the current study, a total of 47 GmUBP gene pairs resulting from segmental duplication were identified, with no evidence of tandem duplications (Figure 4). This observation implies that segmental duplication may predominantly drive the expansion of the GmUBP gene family.
In this study, all GmUBP promoters contain light-responsive elements, which serve as binding sites for transcription factors mediating gene expression in response to light signals (Figure 2). Therefore, light signals may regulate the stability of downstream gene products by influencing the expression of GmUBP genes; this is a process that is associated with photoreception [46]. Similarly, approximately two-thirds of the GmUBP promoters contain anaerobic-responsive elements (Figure 2). These elements represent specific cis-acting sequences found in the promoter regions of the genes activated under low-oxygen conditions, such as flooding or soil waterlogging. Such elements facilitate the binding of transcription factors, thereby enhancing the plant’s ability to respond to hypoxic or anoxic stress through the activation of genes involved in anaerobic responses [47]. Moreover, the promoters of GmUBP genes are characterized by abundant stress-responsive cis-elements, including those associated with low-temperature and drought conditions (Figure 2). This suggests that GmUBP genes likely play significant roles in abiotic stress responses. Specifically, when plants detect abiotic stress signals, certain transcription factors may bind to these cis-elements, activating the transcription of GmUBP genes. The resulting GmUBP proteins then cleave ubiquitin chains from ubiquitinated proteins, thereby regulating multiple physiological pathways to enable the plant to cope with abiotic stresses. Additionally, plant hormones are key signaling molecules that regulate various physiological processes during plant growth and development [47]. Our analysis reveals that the promoter regions of GmUBP genes contain multiple hormone-responsive elements, including those responsive to jasmonic acid, abscisic acid, gibberellin, and auxin (Figure 2). This finding suggests that soybean UBP genes may actively participate in hormone signaling pathways, potentially integrating hormonal regulation with other stress response mechanisms.
Soybean, a vital legume crop, serves as an excellent source of both protein and oil. Enhancing soybean productivity hinges on elucidating and comprehending the mechanisms underlying the regulation of seed-related traits. Research has demonstrated that various UBP (ubiquitin-specific protease) family members play significant and diverse roles in controlling different traits. However, aside from Arabidopsis UBP14 [26], rice OsUBP15 [24], and GmSW17 (the ortholog of AtUBP22) [33], the roles of other UBPs in seed development remain largely unexplored. In this study, we identified GmUBP7—a homolog of Arabidopsis UBP5—which is highly expressed in soybean seeds and potentially regulates seed size and lipid–protein content (Figure 8). Glycine max, the cultivated soybean, was domesticated in China approximately 5000–9000 years ago from its wild progenitor, Glycine soja. This domestication process is typically marked by an increasing trend in 100-seed weight, accompanied by a decline in protein content and a concomitant rise in oil content [48,49,50]. Our analysis reveals that the frequency of GmUBP7-H0 increased throughout the domestication process, while the frequency of H5 decreased. Importantly, the seed phenotypes associated with H0 and H5 aligned with the general trajectory of domestication. These findings suggest that GmUBP7 may function as a key domestication gene, with GmUBP7-H0 playing a pivotal role in the domestication process (Figure 8). Further analysis reveals that H5 exhibited base deletion (CCCAAACAAAGTGATTTA) in the first intron region of the GmUBP7 gene compared to H0, which could alter the gene’s function (Table S8). It has been suggested that variations in introns may impact the binding capacity of upstream transcription factors, thereby regulating gene expression [51,52]. To investigate whether H5 deletion represented a transcription factor binding site, we utilized the online tool PlantPAN4.0. The analyses indicate that the deleted sequence contains recognition sites for NFY family transcription factors, specifically NF-YA, NF-YB, and NF-YC. It is known that NFY family members are involved in regulating seed size and oil content in soybean and maize [53,54,55,56]. Therefore, GmUBP7 may collaborate with NFY family members to regulate seed development, and the phenotypic differences observed between H0-4 and H5 might result from varying NFY transcription factor binding affinities to the GmUBP7 gene.
In conclusion, the bioinformatic analysis of soybean UBP genes presented in this study offers a comprehensive overview of their chromosomal localization, motif and domain structures, expression patterns, and haplotype analysis. These findings provide valuable insights and serve as key references for the further characterization and functional studies of specific GmUBP genes in soybean.
4. Materials and Methods
This section is divided into subheadings. It should provide a concise and precise description of the experimental results and their interpretation, as well as the experimental conclusions that can be drawn.
4.1. Identification and Chromosomal Locations of Soybean UBP Genes
We retrieved the protein and genome sequence files for Glycine max (Wm82.a2.1) from the plant genome database Phytozome (https://phytozome-next.jgi.doe.gov/, accessed on 3 September 2024), and downloaded the protein and genome sequence files for Arabidopsis thaliana and Oryza sativa from the EnsemblPlants database (https://plants.ensembl.org/, accessed on 3 September 2024). [57].
To identify UBP genes in these species, we used two approaches. First, we extracted the conserved domain UCH (PF00443) from Arabidopsis UBP proteins on PFAM (http://pfam.xfam.org, accessed on 3 September 2024) and utilized TBtools’ Advanced Hmmer Search plugin to search for candidate UBP proteins in soybean, with default parameters. Second, we used Arabidopsis UBP proteins as queries against soybean protein sequences in MEGA11.0 with default settings. Proteins lacking the UCH domain or its characteristic Cys-box and His-box residues were excluded.
The genomic distribution of UBP genes was determined using the respective genome and annotation files. Subcellular localization was predicted via the BUSCA server (http://busca.biocomp.unibo.it/, accessed on 9 September 2024) [58]. Physicochemical properties, including amino acid count, molecular weight, and isoelectric point, were analyzed using Tbtools. To further understand their structural features, we predicted transmembrane domains using TMHMM (https://services.healthtech.dtu.dk/service.php?TMHMM-2.0, accessed on 8 September 2024).
4.2. Chromosome Mapping and Phylogenetic Analysis
To determine the genomic positions of all identified UBP genes in soybean, their chromosomal locations were visualized using TBtools (v2.310). Full-length amino acid sequences from Glycine max, Arabidopsis thaliana, and Oryza sativa were aligned using MEGA11.0 with the MUSCLE function. The phylogenetic tree was constructed by subjecting the multiple sequence alignment file to MEGA11, employing the neighbor-joining (NJ) method with 1000 bootstrap replicates, pairwise deletion, and a Poisson model. Finally, the tree was refined using iTOL (https://itol.embl.de/upload.cgi, accessed on 9 January 2025).
4.3. Gene Structure and Conserved Motif Analysis
To examine the gene structure of UBP genes in soybean, TBtools was employed for analysis. The conserved structural motifs of UBP genes were identified using MEME (https://meme-suite.org/meme/tools/meme, accessed on 24 September 2024) [59]. The results from both gene structure and motif analyses were consolidated using TBtools for visualization.
4.4. Analysis of cis-Regulatory Elements of UBP Genes
To examine the upstream regulatory regions of UBP genes in soybean, the genomic sequences spanning 2000 bp upstream of the translation initiation site for each UBP gene were extracted from the genome files. The cis-acting elements within the UBP gene promoters were identified using PlantCare Serve (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 22 September 2024) [60]. Finally, the results of promoter analysis were visualized using TBtools’ Basic Biosequence View function.
4.5. Collinearity Analysis
The Dual-Synteny Plot program in TBtools was used to analyze the homology of UBP genes across maize and other species, including Arabidopsis, soybean, and rice. To further examine collinearity, we implemented the One-Step MCScanX tool from TBtools [61,62]. The resulting data were visualized using TBtools to provide a comprehensive overview of the homology relationships among these species.
4.6. Expression Analysis
SoyBase (http://www.soybase.org, accessed on 29 September 2024) served as a platform for extracting GmUBP expression profile data across diverse tissues and developmental stages. Heatmap visualizations of the expression profiles were generated using TBtools. For qRT-PCR, we first planted soybean (W82) in a short-day (a light–dark ratio of 10 h/14 h) greenhouse and sampled tissues at different stages of plant development. Total RNA was extracted from different samples using an Eastep^®^ Super Total RNA Extraction Kit (Promega, Shanghai, China), and cDNA was synthesized from the RNA by a reverse transcription reagent kit (HiScript II 1st Strand cDNA Synthesis Kit (Vazyme, Nanjing, China). Real-time quantitative PCR was performed using 2 × ChamQ Universal SYBR qPCR Master Mix (Vazyme, Nanjing, China). GmActin was used as an internal control gene. The gene-specific primers are listed in Table S9.
4.7. Haplotype Analysis
The Soybean Multi-omics Database (https://yanglab.hzau.edu.cn/SoyMD/#/, accessed on 13 February 2025) was employed to investigate the haplotype structure of GmUBP genes in soybean. During the analysis, the single-locus model was selected to evaluate the gene haplotypes. By inputting the corresponding gene identifier, researchers can retrieve allele frequency data specific to sub-populations. Furthermore, the platform enables the integration of existing phenotypic records with haplotype information, facilitating statistical analysis to determine significant differences between haplotypes.
5. Conclusions
In this study, we conducted a comprehensive investigation into the GmUBP gene family. Initially, phylogenetic analysis was employed to classify the 52 identified GmUBP genes into 14 distinct groups based on their evolutionary relationships with UBP genes from Arabidopsis, rice, and soybean. Subsequent analyses, including exon–intron structure characterization, the identification of conserved domains, and motif analysis, provide robust confirmation of the high conservation of GmUBP genes within the same group. Synteny analysis was then employed to elucidate the evolutionary trajectory of the GmUBP gene family. Furthermore, promoter cis-regulatory element analysis, combined with tissue-specific expression profiling in Glycine max, reveals the potential involvement of GmUBP genes in key aspects of plant growth and development. Notably, we identified that GmUBP7 may have undergone selection during domestication and appears to play a role in the coordinated regulation of seed size and quality. Our findings offer a valuable foundation for future research on the GmUBP gene family, particularly with respect to soybean seed development, and provide a scientific framework for exploring the biological functions and mechanisms underlying GmUBP gene activity.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Graham P.H. Vance C.P. Legumes: Importance and constraints to greater use Plant Physiol.200313187287710.1104/pp.01700412644639 PMC 1540286 · doi ↗ · pubmed ↗
- 2Li N. Li Y. Signaling pathways of seed size control in plants Curr. Opin. Plant Biol.201633233210.1016/j.pbi.2016.05.00827294659 · doi ↗ · pubmed ↗
- 3Zhou H. Zhao J. Cai J. Patil S.B. UBIQUITIN-SPECIFIC PROTEASES function in plant development and stress responses Plant Mol. Biol.20179456557610.1007/s 11103-017-0633-528695315 · doi ↗ · pubmed ↗
- 4Frappier L. Verrijzer C.P. Gene expression control by protein deubiquitinases Curr. Opin. Genet. Dev.20112120721310.1016/j.gde.2011.02.00521411309 · doi ↗ · pubmed ↗
- 5Zhang Y. Transcriptional regulation by histone ubiquitination and deubiquitination Genes Dev.2003172733274010.1101/gad.115640314630937 · doi ↗ · pubmed ↗
- 6Atanassov B.S. Koutelou E. Dent S.Y. The role of deubiquitinating enzymes in chromatin regulation FEBS Lett.20115852016202310.1016/j.febslet.2010.10.04220974139 PMC 3036772 · doi ↗ · pubmed ↗
- 7Neutzner M. Neutzner A. Enzymes of ubiquitination and deubiquitination Essays Biochem.20125237502270856210.1042/bse 0520037 · doi ↗ · pubmed ↗
- 8Amerik A.Y. Hochstrasser M. Mechanism and function of deubiquitinating enzymes Biochim. Biophys. Acta 2004169518920710.1016/j.bbamcr.2004.10.00315571815 · doi ↗ · pubmed ↗