Brain and CSF Alzheimer’s Biomarkers Are Associated with SERPINE1 Gene Expression
Cynthia Picard, Henrik Zetterberg, Kaj Blennow, Sylvia Villeneuve, Judes Poirier

TL;DR
This study shows that SERPINE1 levels in cerebrospinal fluid and brain tissue are linked to Alzheimer’s disease biomarkers, suggesting it could be an early detection tool.
Contribution
The study links SERPINE1 levels to tau pathology and Braak staging, offering new insights into its potential as an early Alzheimer’s biomarker.
Findings
Elevated CSF SERPINE1 levels correlate with pTau, tTau, and synaptic markers in pre-clinical Alzheimer’s.
Cortical SERPINE1 mRNA levels in AD brains correlate strongly with Braak staging but weakly with amyloid plaques.
SERPINE1 shows stronger association with tau pathology than with amyloid formation.
Abstract
Background: SERPINE1, also known as plasminogen activator inhibitor (PAI), has been proposed as a potential blood biomarker for the early detection and diagnosis of Alzheimer’s disease (AD). Expanding on previous studies, this research contrasted SERPINE1 levels in CSF and brain tissue of AD patients and those at risk for AD with established AD biomarkers. Methods: Utilizing OLINK and immunoassay methods, CSF SERPINE1 protein levels were quantified across two separate cohorts: PREVENT-AD and ADNI. Microarray and RNAseq were used to measure tissue SERPINE1 mRNA levels in two separate cohorts: the Douglas-Bell Canada Brain Bank and the Mayo Clinic Brain Bank. Results: At the pre-clinical stage, elevated CSF levels of pTau, tTau and synaptic markers, alongside reduced hippocampal volume, correlate with CSF SERPINE1 levels. Elevated cortical SERPINE1 mRNA levels in autopsy-confirmed AD show…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
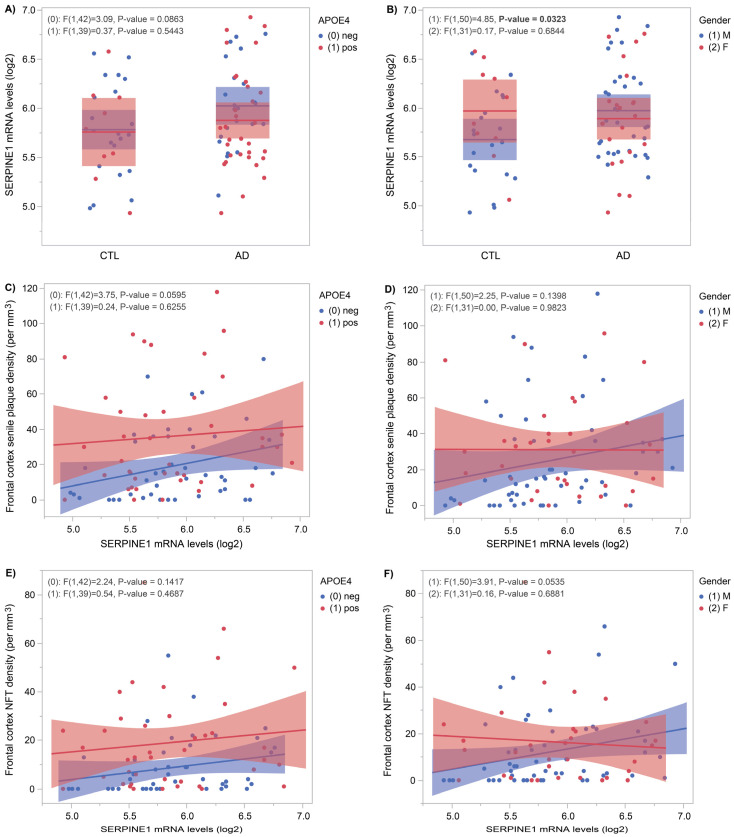 Figure 1
Figure 1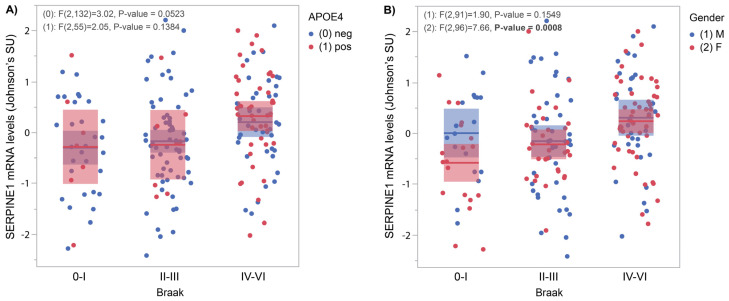 Figure 2
Figure 2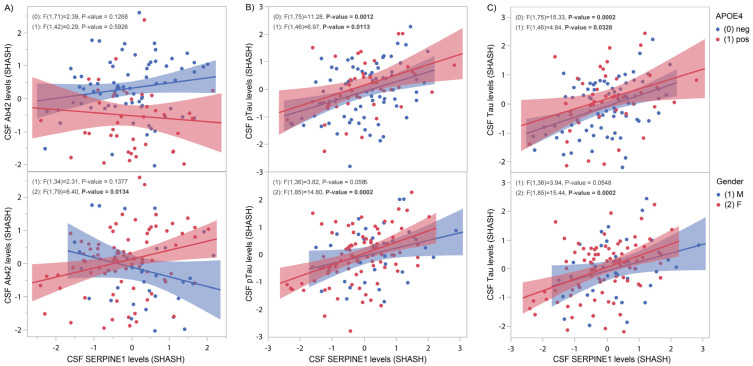 Figure 3
Figure 3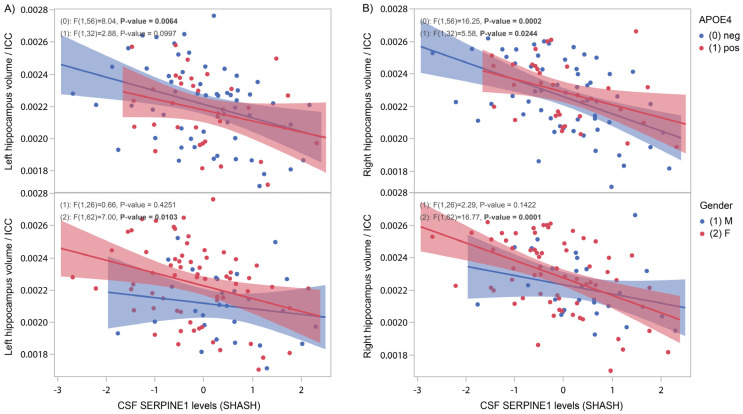 Figure 4
Figure 4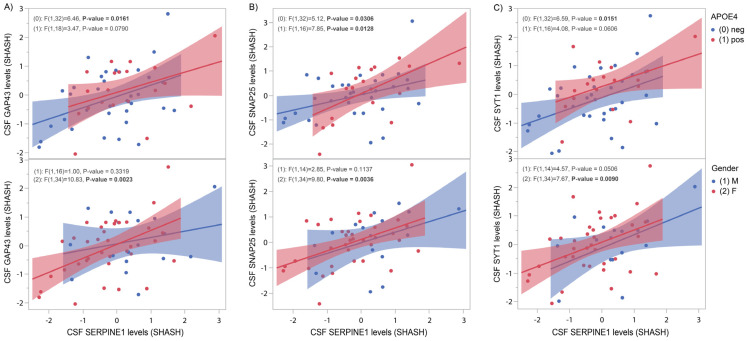 Figure 5
Figure 5- —Fonds de la Recherche en Santé du Québec (FRSQ), the Canadian Institute for Health Research
- —National Research Council of Canada (NSERC)
- —J.L. Levesque Foundation and Brain Canada
- —FRQS, the CIHR
- —Brain Canada
- —Swedish Research Council
- —European Union’s Horizon Europe research and innovation programme
- —Swedish State Support for Clinical Research
- —Alzheimer Drug Discovery Foundation (ADDF), USA
- —AD Strategic Fund and the Alzheimer’s Association
- —Bluefield Project
- —Cure Alzheimer’s Fund
- —Olav Thon Foundation
- —Erling-Persson Family Foundation
- —Familjen Rönströms Stiftelse
- —Stiftelsen för Gamla Tjänarinnor, Hjärnfonden, Sweden
- —European Union’s Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant
- —European Union Joint Programme—Neurodegenerative Disease Research
- —National Institute for Health and Care Research University College London Hospitals Biomedical Research Centre
- —UK Dementia Research Institute at UCL
- —Swedish Research Council
- —Swedish Alzheimer Foundation
- —Hjärnfonden, Sweden
- —Swedish state via the agreement between the Swedish government and the County Councils
- —ALF-agreement
- —European Union Joint Program for Neurodegenerative Disorders
- —Alzheimer’s Association 2021 Zenith Award
- —Alzheimer’s Association 2022–2025 Grant
- —La Fondation Recherche Alzheimer (FRA), Paris, France
- —Kirsten and Freddy Johansen Foundation, Copenhagen, Denmark
- —Familjen Rönströms Stiftelse, Stockholm, Sweden
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAlzheimer's disease research and treatments · S100 Proteins and Annexins · Dementia and Cognitive Impairment Research
1. Introduction
SERPINE1 (plasminogen activator inhibitor, PAI) encodes a member of the serine proteinase inhibitor superfamily. It is one of the most important inhibitors of fibrinolytic activity thus leading to pathological fibrin deposition and tissue damage [1]. This role is attributed to the fact that SERPINE1 is the principal inhibitor of tissue plasminogen activator (tPA, encoded by the gene PLAT) and urokinase-type plasminogen activator (uPA, encoded by the gene PLAU).
Plasminogen is expressed primarily by the liver and is therefore present in the plasma, but mRNA levels have been detected in various tissues including brain, adrenal, kidney, testis, heart, lung, uterus, spleen thymus and gut [2]. The plasminogen activator system is pivotal in various physiological and pathological processes, encompassing coagulation, fibrinolysis, inflammation, wound healing and malignancy [1].
SERPINE1 is also known to inhibit cell migration by competing for vitronectin binding to integrins [3]. Recent findings reveal vitronectin plays multiple roles within the nervous system, where it contributes to activities such as neural differentiation and neurogenesis, while also having a role in regulating axon size and providing support and guidance for neurite extension [4]. By interacting with integrin receptors in vascular endothelial cells, vitronectin can reduce the permeability of the blood–brain barrier [4].
Amyloid beta (Aβ), one of the hallmarks of Alzheimer’s disease (AD), is known to increase tPA and uPA [5]. In turn, tPA and uPA cleave plasminogen (encoded by the gene PLG), resulting in the activation of the serine protease plasmin [5]. Plasmin was shown to degrade Aβ fibrils, therefore attenuating Aβ neurotoxicity [5]. In AD, brain tissues contain reduced levels of plasmin when compared to controls [6].
SERPINE1 was shown to accumulate in blood samples from mild cognitively impaired (MCI) and AD patients [7]. Given the established correlation between plasma SERPINE1 and cognitive impairment, SERPINE1 is proposed as a potential biomarker for early AD detection and diagnosis [7]. Consistent with this finding, SERPINE1 hippocampal RNA prevalence is significantly increased in AD patients relative to controls [8].
In the present study, we examined SERPINE1 concentrations in control and AD brain tissue and contrasted SERPINE1 mRNA levels as a function of neurofibrillary tangles density, senile plaque density and Braak stages. To validate SERPINE1 as a potential presymptomatic AD biomarker, we analyzed data from the PREVENT-AD cohort, which consists of asymptomatic individuals with a family history of late-onset AD. CSF SERPINE1 protein levels were measured and contrasted with classical AD pathological biomarkers such as tTau, pTau181 tau and Aβ42. We replicated our biochemical findings in living ADNI subjects spanning various disease stages.
2. Materials and Methods
2.1. Study Participants from the Douglas-Bell Canada Brain Bank Cohort
Fifty-five autopsy-confirmed AD brains and thirty autopsy-confirmed control brains were obtained from the Douglas-Bell Canada Brain Bank (see Table 1 for demographics). Based on reviews of medical records, neuropsychological assessments and care caregiver interviews, there was no indication of memory impairment, neurological or neuropsychiatric diseases in the older adults control group. Controls only exhibited neuropathology that is consistent with normal ageing, characterized by plaque and tangle densities of less than 10/mm^3^ and 20/mm^3^, respectively, in at least one section of the hippocampus and neocortex. AD cases had to fulfill the histopathological NINCDS-ADRDA criteria for definite AD [9]. Written informed consent was obtained from each participant and their study partner. All procedures were approved by the McGill University Faculty of Medicine Institutional Review Board and by the Ethics Board of the Douglas Mental Health University Institute. It complies with ethical principles of the Declaration of Helsinki and conformed to the Code of Ethics of the World Medical Association as per protocol approval number #IUSMD-02-34, 17 September 2024.
Total RNA was extracted from 100 mg of cortical brain tissues and quantified using a NanoDrop spectrophotometer ND-100 (NanoDrop Technologies, Inc, Wilmington, NC, USA). RNA integrity was assessed using a 2100 Bioanalyzer (Agilent Technologies, Saint-Laurent, QC, Canada) and samples with RNA integrity number below 3.5 were removed. Sense-strand cDNA was synthesized from 10 ng of total RNA, followed by fragmentation and labelling using the GeneChip WT Pico terminal labelling kit, in accordance with the manufacturer’s protocol (Thermo Fisher Scientific, Saint-Laurent, QC, Canada). Subsequently, 5 mg of the labelled DNA target was hybridized on GeneChip Clariom^TM^ D human (Thermo Fisher Scientific, Saint-Laurent, QC, Canada) and incubated at 45 °C in the GeneChip Hybridization oven 640 (Affymetrix, Santa Clara, CA, USA) for 17 h at 60 rpm. Post-hybridization, the arrays were washed in a GeneChips Fluidics Station 450 (Thermo Fisher, Saint-Laurent, QC, Canada) using GeneChip Hybridization Wash and Stain kit according to manufacturer’s guidelines (Thermo Fisher, Saint-Laurent, QC, Canada). Finally, the microarrays were scanned on a GeneChip scanner 3000 (Thermo Fisher, Saint-Laurent, QC, Canada). Gene expressions were normalized using the Transcriptome Analysis Software from Thermo Fisher Scientific (Saint-Laurent, QC, Canada). SERPINE1 mRNA levels are presented on a log_2_ scale.
2.2. Study Participants from the Mayo Clinic/Banner Brain Bank Cohort
A total of 193 subjects from the Mayo Clinic Brain Bank and Banner Sun Health institute were included, comprising individuals with Alzheimer’s disease (AD; n = 82), progressive supranuclear palsy (PSP; n = 83) and cognitively unimpaired (CU; n = 28). Control subjects met the following criteria: Braak NFT stage of 3.0 or less, CERAD neuritic and cortical plaque densities of 0 (none) or 1 (sparse) and an absence of any pathological diagnoses, including AD, Parkinson’s disease, Lewy body disease, vascular dementia, progressive supranuclear palsy, motor neuron disease, corticobasal degeneration, Pick’s disease, Huntington’s disease, frontotemporal lobar degeneration, hippocampal sclerosis or dementia without distinctive histology. All participants were North American Caucasians who were aged 60 or older at the time of their death (refer to Table 1 for demographic details). All procedures complied with the ethical principles of the Declaration of Helsinki. Gene expression data were retrieved from AMP-AD Knowledge Portal https://adknowledgeportal.synapse.org (synapse ID: syn5550404, accessed on 1 November 2024). Expression profiles were generated using next-generation RNA-sequencing (RNAseq) from temporal cortex RNA samples collected for 193 individuals enrolled through the Mayo Clinic Brain Bank and Banner Sun Health research institute [10].
2.3. Study Participants from the Asymptomatic PREVENT-AD Cohort
The PRe-symptomatic EValuation of Experimental or Novel Treatments for Alzheimer’s Disease (PREVENT-AD) cohort consists of asymptomatic, “at-risk” individuals with a parental or multi-sibling history of sporadic AD [11]. Most participants were over the age of 60 at the time of recruitment (see Table 1 for demographics); however, individuals aged 55–59 years were eligible if they were within 15 years of the age of symptom onset of their youngest-affected relative. Cognitive normality was confirmed at the eligibility visit using the Clinical Dementia Rating (CDR) and the Montreal Cognitive Assessment (MoCA). PREVENT-AD participants were followed longitudinally, with annual visits involving cognitive evaluations, neurosensory testing, blood and CSF collection (for a subset of individuals), structural and functional magnetic resonance imaging (MRI) and positron emission tomography (PET) scans. Each participant and their study partner provided written informed consent. All study procedures were approved by the McGill University Faculty of Medicine Institutional Review Board and adhered to the ethical standards outlined in the Declaration of Helsinki. A detailed description of the PREVENT-AD cohort is available in previously published work [11].
2.3.1. Cerebrospinal Fluid Measurements of the PREVENT-AD Cohort
Lumbar punctures (LP) were performed in PREVENT-AD participants using a Sprotte 24-gauge atraumatic needle following an overnight fast. To remove cells and insoluble material, CSF samples were centrifuged (~2000× g) for 10 min at room temperature within 4 h of collection. The resulting supernatant was aliquoted into 0.5 mL polypropylene cryotubes and stored at −80 °C.
CSF levels of SERPINE1 were quantified in a subset of PREVENT-AD participants (n = 125) using the OLINK Cardiovascular III panel, which utilizes proximity extension assay (PEA) technology as per manufacturer’s instructions (Uppsala, Sweden). Results are reported in arbitrary normalized protein expression units (NPX).
In the same subset (n = 125), CSF concentrations of AD biomarkers Aβ42, phosphorylated tau (pTau181) and total tau (tTau) were measured using the validated Innotest enzyme-linked immunosorbent assay (ELISA) kit (Fujirebio, Ghent, Belgium) following the standardized protocols established by the BIOMARKAPD consortium (Aβ42 Cat.# 81583, pTau181 Cat.# 81581 and tTau Cat.# 81579).
CSF GAP43, SNAP25 and SYT1 were immunoprecipitated and their concentrations were measured by mass spectrometry, as previously described [12]. Mass spectrometry results are expressed in arbitrary units.
2.3.2. Hippocampus Volume of the PREVENT-AD Cohort
MRI acquisition protocols have been previously described in detail [13]. In brief, T_1_-weighted images for each participant were segmented into gray matter, white matter and CSF using Statistical Parametric Mapping 12 v.6225. running on MATLAB version 2012a [14]. Total intracranial volume was computed as the sum of the three segmentation volumes. Subject-specific templates were generated using the DARTEL toolbox [15] and all scans were non-linearly normalized to these templates. The resulting templates were then registered to the MNI-ICBM152 standard space using non-linear transformations with modulation for both linear and non-linear deformations. These transformation parameters were applied to each subject’s T_1_ image to align it with the template space. Following segmentation and normalization, all images underwent visual quality control. Gray matter probability (c1) maps in template space were smoothed with an 8 mm^3^ isotropic Gaussian kernel to produce grey matter volume images. These were then masked using a maximum probability grey matter mask derived from the group-average image. For comparative analyses, hippocampal volume measures were extracted in native space for all participants using a previously validated method [16].
2.3.3. APOE Genotyping
The QIAsymphony apparatus and DNA Blood Mini QIA Kit (Qiagen, Valencia, CA, USA) were used to isolate DNA from brains or whole blood. The standard QIASymphony protocol was used in accordance with the manufacturer’s guidelines. The PyroMark Q96 pyrosequencer (Qiagen, Toronto, ON, Canada) was used to determine APOE genotype, with the following primers purchased from Eurofins genomics (Toronto, ON, Canada): rs429358 amplification forward 5’-ACGGCTGTCCAAGGAGCTG-3’, rs429358 amplification reverse biotinylated 5’-CACCTCGCCGCGGTACTG-3’, rs429358 sequencing 5’CGGACATGGAGGACG-3’, rs7412 amplification forward 5’-CTCCGCGATGCCGATGAC-3’, rs7412 amplification reverse biotinylated 5’-CCCCGGCCTGGTACACTG-3’ and rs7412 sequencing 5’-CGATGACCTGCAGAAG-3’.
2.4. Statistical Analysis
All statistical analysis including linear and logistic regressions were performed using JMP Pro 16.
2.5. Data Availability
Data collected from the Douglas-Bell Canada Brain Bank are not publicly available; however, data can be provided by the corresponding author upon reasonable request. Gene expression data from the Mayo Clinic were downloaded from AMP-AD Knowledge Portal https://adknowledgeportal.synapse.org (synapse ID: syn5550404, accessed on 1 November 2024). CSF, genetic and clinical data from the ADNI cohort were downloaded from the ADNI website (http://adni.loni.usc.edu/, accessed on 1 November 2024) following registration.
The PREVENT-AD multimodal data, including subject characteristics, CSF biomarkers levels, tau and amyloid-PET SUVR, APOE genotypes and RBANS scores, have been summarized in the supporting data files of the PREVENT-AD data release 7, which are available from the corresponding authors upon reasonable request at https://openpreventad.loris.ca/ and following registration.
3. Results
3.1. SERPINE1 mRNA Levels Correlate with AD Pathology
Figure 1 shows SERPINE1 mRNA levels measured in frontal cortices of age-matched controls (n = 30) and AD (n = 55) subjects from the Douglas-Bell Canada Brain Bank. Stratified by APOE4 status, Figure 1A shows a near-significant difference (p = 0.0863) in APOE4-negative subjects, whereas Figure 1B shows a significant increase in AD males versus CTL males (p = 0.03). When stratified by APOE4 status (Figure 1C), a near-significant positive correlation (p = 0.06) emerged between SERPINE1 mRNA levels and senile plaque density. Despite the gender stratification shown in Figure 1D, no significant correlations were found with senile plaque density. When comparing with neurofibrillary tangles density, no significant correlations appeared after stratifying by APOE4 status (Figure 1E). After sex-specific analysis (Figure 1F), a significant correlation (p = 0.05) between SERPINE1 mRNA levels and NFT density was found only in males.
Figure 2 shows SERPINE1 mRNA levels measured by RNAseq in frontal cortices obtained from the Mayo Clinic/Banner brain bank cohort. Samples include 111 non-AD subjects (28 CTLs and 83 PSP) and 82 AD subjects for which Braak stages were split into three groups. Figure 2A shows a week relationship with APOE4 status (p = 0.05); Figure 2B shows a correlation between SERPINE1 mRNA levels and Braak stages (p = 0.0008) only in females.
3.2. CSF SERPINE1 Protein Levels Correlate Positively with AD Biomarkers
In a cohort of 125 “at-risk” but cognitively unaffected PREVENT-AD individuals, CSF samples were used to measure protein levels of SERPINE1 and the AD biomarkers Aβ42, pTau181 and tTau. Positive correlations with Aβ42 (Figure 3A, p = 0.01), pTau181 (Figure 3B, p = 0.0002) and tTau (Figure 3C, p = 0.0002) were found exclusively in females. APOE4-negative subjects display positive correlations with pTau181 (Figure 3B, p = 0.001) and tTau (Figure 3C, p = 0.0002). In APOE4-positive subjects, positive correlations were found with pTau181 (Figure 3B, p = 0.01) and tTau (Figure 3C, p = 0.03).
3.3. SERPINE1 Protein Levels Correlate Negatively with Hippocampus Volume
In a subset of participants from PREVENT-AD undergoing MRI (n = 92), hippocampus volume was measured and contrasted with CSF SERPINE1 protein levels. In APOE4-negative subjects, a strong negative correlation was found with both left (Figure 4A, p = 0.006) and right (Figure 4B, p = 0.0002) hippocampus volume. Analysis by gender revealed a significant inverse correlation between hippocampus volume and the outcome measure only in females (Figure 4A, left: p = 0.01; and Figure 4B, right: p < 0.0001).
3.4. SERPINE1 Protein Levels Correlate Positively with Synaptic Markers
Figure 5 shows significant positive correlations between CSF SERPINE1 protein levels and three synaptic markers: GAP43 (Figure 5A, APOE4 neg, p = 0.016), SNAP25 (Figure 5B. APOE4 neg, p = 0.03; APOE4 pos, p = 0.013) and SYT1 (Figure 5C, APOE4 neg, p = 0.015). Figure 5 (lower panels) reveals significant gender-specific correlations between SERPINE1 and GAP43 (Figure 5A, females, p = 0.002), SNAP25 (Figure 5B, females, p = 0.0036) and SYT1 (Figure 5C, males, p = 0.05; females, p = 0.009).
3.5. Quantitative Trait Loci Analysis of CSF SERPINE1 Levels Identifies Naturally Occurring Polymorphisms in the SERPINE1 and VAT1L Genes
Using trans-pQTL analysis, we identified a genome-wide significant polymorphism (rs17705051 downstream of the VAT1L gene, Supplemental Figure S4) that affects SERPINE1 levels in the CSF of cognitively unaffected PREVENT-AD subjects. Rs6092, a coding SNP, was identified as the most significant cis-pQTL influencing polymorphism using CSF SERPINE1 protein levels in the ADNI cohort (Supplemental Figure S5). While the presence of these variants has no impact on AD risk levels (Table 2) in three cohorts (ADNI, MAYO, DBCBB), they significantly affect protein concentrations of SERPINE1 in the PREVENT-AD and ADNI cohorts and the mRNA levels in the MAYO cohort. As for AD biomarkers, only the presymptomatic PREVENT-AD cohort display significant association between the vesicle amine transport 1 like (VAT1L) variant and pTau181 (p = 0.006) as well as total tau levels in the CSF (p = 0.005).
4. Discussion
SERPINE1 was originally identified as a negative regulator of fibrinolysis by preventing the dissolution of blood clots [17]. Increased concentration of SERPINE1 in plasma can cause venous thrombosis and myocardial infarction [18]. More recently, myocardial infarction was proposed to be a predisposing factor for AD development [19]. Mounting evidence suggests that the ageing of the neurovasculature is a critical determinant of brain ageing and AD risk [20]. In neurodegenerative conditions, such as Alzheimer’s disease, Parkinson’s disease and frontotemporal lobar degeneration, plasma levels of SERPINE1 were found in higher concentrations [21,22]. These associations prompted us to examine SERPINE1 neurobiology throughout AD spectrum, i.e., before and after AD onset in multiple living and autopsied cohorts.
Using autopsy-confirmed AD brains, we showed that cortical SERPINE1 mRNA levels are significantly higher compared to CTL brains (Figure 1B, p = 0.03). This association is also slightly affected by both APOE4 status and gender. These results are consistent with the previous literature reporting the upregulation of SERPINE1 mRNA levels in the superior frontal gyrus, entorhinal cortex and hippocampus of AD subjects compared to age-matched controls [8]. Only AD patients without APOE4 in the ADNI CSF cohort showed increased SERPINE1 (Figure S1).
Plasmin, when inhibited by SERPINE1, is known to facilitate the degradation of Aβ fibrils [5] and is found in low concentrations in AD versus control brains [6]. In this context, it was tempting to propose that SERPINE1 could affect senile plaque density in a regional and concentration-dependent manner. In Figure 1C, we found such interaction at a trend level but only in APOE4-negative subjects. In the same vein, we examined the influence of SERPINE1 on neurofibrillary tangles density and found a weak correlation between the two, but only in males (Figure 1F).
Using a replication cohort from the Mayo brain bank with a larger sample size, we investigated the influence of SERPINE1 on tangle spreading using Braak staging [23]. A positive association was found between cortical SERPINE1 mRNA levels and Braak stages in females only (Figure 2B). It is of note that the proportion of females was higher in the AD group compared to non-AD, which could explain, at least in part, those results. Nevertheless, a positive correlation with both NFT (Figure 1F) and Braak (Figure 2) suggests a possible influence on Tau metabolism and/or deposition.
To determine if SERPINE1 could be used as an early, presymptomatic, AD biomarker, we turned to the PREVENT-AD cohort, comprising asymptomatic “at-risk” individuals. Figure 3 shows strong correlations between CSF SERPINE1 protein levels and pTau181 and tTau, especially in females and APOE4-negative subjects (see also Figure S2 for ADNI results). These results are consistent with the tangles/SERPINE1 association described earlier in fully established AD cases. Note that for Aβ, the expected negative correlation with SERPINE1 was not established (Figure 3A and Figure S2A).
Using proteomic quantitative trait loci analysis, we identified genome-wide and locus-wide significant (naturally occurring) polymorphisms that display significant association with SERPINE1 protein levels in the CSF of our asymptomatic cohort of subjects (Supplemental Figures S4 and S5). The two variants that modulates CSF SERPINE1 concentration, rs17705051 (VAT1L) and rs6092 (SERPINE1) failed to affect AD risk levels when assessed in our three distinct cohorts (Table 2). Using the larger publicly available IGAP GWAS database (https://www.nature.com/articles/ng.2802, accessed on 1 November 2024), we performed additional screening for AD risk in this large dataset (n = 74,046) and found no significant genome-wide association with sporadic AD for rs17705051 (p = 0.1779) nor for rs6092 (p = 0.002).
A previous study involving the ADNI cohort identified an intronic SNP in VAT1L in association with cognitive decline measured by ADAS-cog [24]. In our asymptomatic cohort, the SNP affecting CSF SERPINE1 protein levels was found downstream of the VAT1L gene. Further studies will be necessary to establish the link between VAT1L and SERPINE1. The cis-acting SNP rs6092 is a coding SNP associated with impaired secretion of SERPINE1 and hemorrhage problem [25]. Other SNPs in SERPINE1 were shown to affect AD risk [26,27].
In the early stages of AD, there is a swift degeneration of hippocampal tissue, which can be measured by magnetic resonance imaging [28]. In the PREVENT-AD cohort, we show that CSF SERPINE1 correlates inversely with hippocampus volume, particularly in females and APOE4-negative subjects (Figure 4 and Supplemental Figure S3 for ADNI results). These results suggest that in the absence of symptoms in at-risk individuals, hippocampal loss can be predicted by the presence of high levels of SERPINE1 in the CSF.
In preclinical stages of AD, changes in synaptic proteins were shown to precede CSF markers of neurodegeneration [29]. Increased concentrations of the presynaptic markers GAP43, SNAP25 and SYT1 were observed in the CSF during neurodegeneration [12,30,31]. In the asymptomatic PREVENT-AD cohort, we found that CSF SERPINE1 positively correlates with those three pre-synaptic markers in both APOE4-negative and -positive subjects (Figure 5, upper panels) as well as in all females (Figure 5, lower panels). Previous studies suggest that cognitive deficit is more closely associated with abnormalities in synaptic structure and function than with the quantity of plaques and tangles [21,32].
Our research indicates a connection between SERPINE1 and the advancement of AD, most notably in the disease’s asymptomatic period. More specifically, SERPINE1 correlates with early signs of neurodegeneration, namely with elevated levels of tTau and a reduction in hippocampus volume. In parallel, SERPINE1 is strongly linked to pTau181 tau alterations in the CSF and the presence of high concentration of synaptic markers. Over time, when AD pathology sets in, SERPINE1 mRNA levels are elevated and show a weak correlation with plaques and NFT densities but a more significant correlation with tangle spreading monitored by Braak staging.
Our results support the notion that SERPINE1 can be used as an early biomarker for the detection of AD-associated pathological changes in asymptomatic subjects with a parental history of AD. Interestingly, most of the significant results are observed in APOE4-negative subjects, who are usually at lower risk of developing AD. SERPINE1’s strongest association is with tau pathology, a surprising finding considering the plasminogen activator system’s role in Aβ fibril degradation. SERPINE1’s biological network includes proteins such as PLAT, TP53 and IL6 that could possibly have an indirect effect on Tau phosphorylation via the main kinase GSK3B (Figure S6).
Further investigations are necessary to determine if the pharmacomodulation of SERPINE1 could be used to modulate disease progression.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cesari M. Pahor M. Incalzi R.A. Plasminogen activator inhibitor-1 (PAI-1): A key factor linking fibrinolysis and age-related subclinical and clinical conditions Cardiovasc. Ther.201028 e 72e 9110.1111/j.1755-5922.2010.00171.x 20626406 PMC 2958211 · doi ↗ · pubmed ↗
- 2Zhang L. Seiffert D. Fowler B.J. Jenkins G.R. Thinnes T.C. Loskutoff D.J. Parmer R.J. Miles L.A. Plasminogen has a broad extrahepatic distribution Thromb. Haemost.20028749350111916082 · pubmed ↗
- 3Kjoller L. Kanse S.M. Kirkegaard T. Rodenburg K.W. Ronne E. Goodman SL Preissner K.T. Ossowski L. Andreasen P.A. Plasminogen activator inhibitor-1 represses integrin- and vitronectin-mediated cell migration independently of its function as an inhibitor of plasminogen activation Exp. Cell Res.199723242042910.1006/excr.1997.35409168821 · doi ↗ · pubmed ↗
- 4Ruzha Y. Ni J. Quan Z. Li H. Qing H. Role of Vitronectin and Its Receptors in Neuronal Function and Neurodegenerative Diseases Int. J. Mol. Sci.2022231238710.3390/ijms 23201238736293243 PMC 9604229 · doi ↗ · pubmed ↗
- 5Tucker H.M. Kihiko M. Caldwell J.N. Wright S. Kawarabayashi T. Price D. Walker D. Scheff S. Mc Gillis J.P. Rydel R.E. The plasmin system is induced by and degrades amyloid-beta aggregates J. Neurosci.2000203937394610.1523/JNEUROSCI.20-11-03937.200010818128 PMC 6772619 · doi ↗ · pubmed ↗
- 6Ledesma M.D. Da Silva J.S. Crassaerts K. Delacourte A. De Strooper B. Dotti C.G. Brain plasmin enhances APP alpha-cleavage and Abeta degradation and is reduced in Alzheimer’s disease brains EMBO Rep.2000153053510.1093/embo-reports/kvd 10711263499 PMC 1083779 · doi ↗ · pubmed ↗
- 7Oh J. Lee H.J. Song J.H. Park S.I. Kim H. Plasminogen activator inhibitor-1 as an early potential diagnostic marker for Alzheimer’s disease Exp. Gerontol.201460879110.1016/j.exger.2014.10.00425304332 · doi ↗ · pubmed ↗
- 8Magistri M. Velmeshev D. Makhmutova M. Faghihi M.A. Transcriptomics Profiling of Alzheimer’s Disease Reveal Neurovascular Defects, Altered Amyloid-beta Homeostasis, and Deregulated Expression of Long Noncoding RN As J. Alzheimer’s Dis.20154864766510.3233/JAD-15039826402107 PMC 4698155 · doi ↗ · pubmed ↗