Functional and Regulatory Effects of Factor V Leiden and Factor V rs6028 in Breast Cancer
Sara Marie Lind, Marit Sletten, Carola Elisabeth Henriksson, Mari Tinholt, Nina Iversen

TL;DR
This study explores how two genetic variants of the coagulation factor V gene may influence breast cancer progression by affecting gene expression and cell death.
Contribution
The study identifies novel regulatory effects of F5 rs6025 and rs6028 on FV expression and apoptosis in breast cancer cells.
Findings
F5 rs6025 and rs6028 variants increased FV mRNA, protein, coagulant activity, and apoptosis compared to wild type.
Cis-eQTL analysis showed F5 rs6028 has a regulatory effect on F5 expression.
Transcription factor binding sites overlap with F5 rs6025, suggesting a regulatory role.
Abstract
Background/Objectives: Cancer progression and the hemostatic system are closely linked. Coagulation factor V (FV) has a key function in coagulation, with both pro- and anticoagulant functions. FV gene (F5) expression and F5 variants have been linked to breast cancer progression. The direct impact of F5 variants on FV expression and functional effects in breast cancer are unknown. We aimed to investigate whether the F5 variants FV Leiden (F5 rs6025) and F5 rs6028 influenced FV expression, coagulant activity, and apoptosis in breast cancer cells. Methods: MDA-MB-231 cells were transfected with overexpression plasmids containing F5 wild type, F5 rs6025 or F5 rs6028. We investigated the functional impact of the F5 variants on F5 mRNA, FV protein, FV coagulant activity, and apoptosis in vitro, and examined the potential of the variants as transcriptional regulators of F5 expression in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
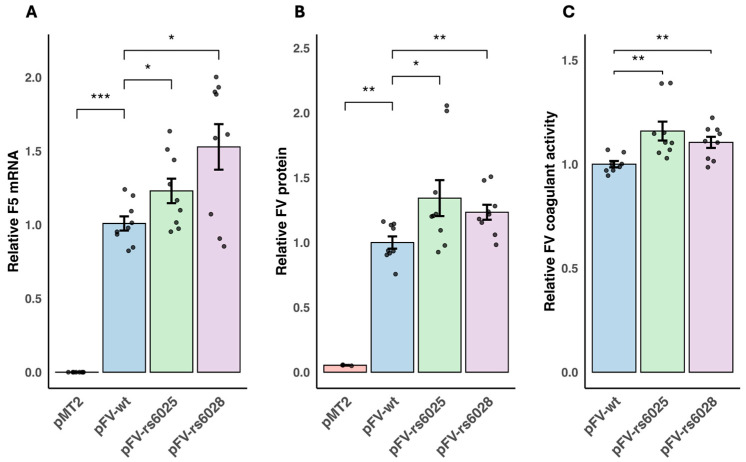 Figure 1
Figure 1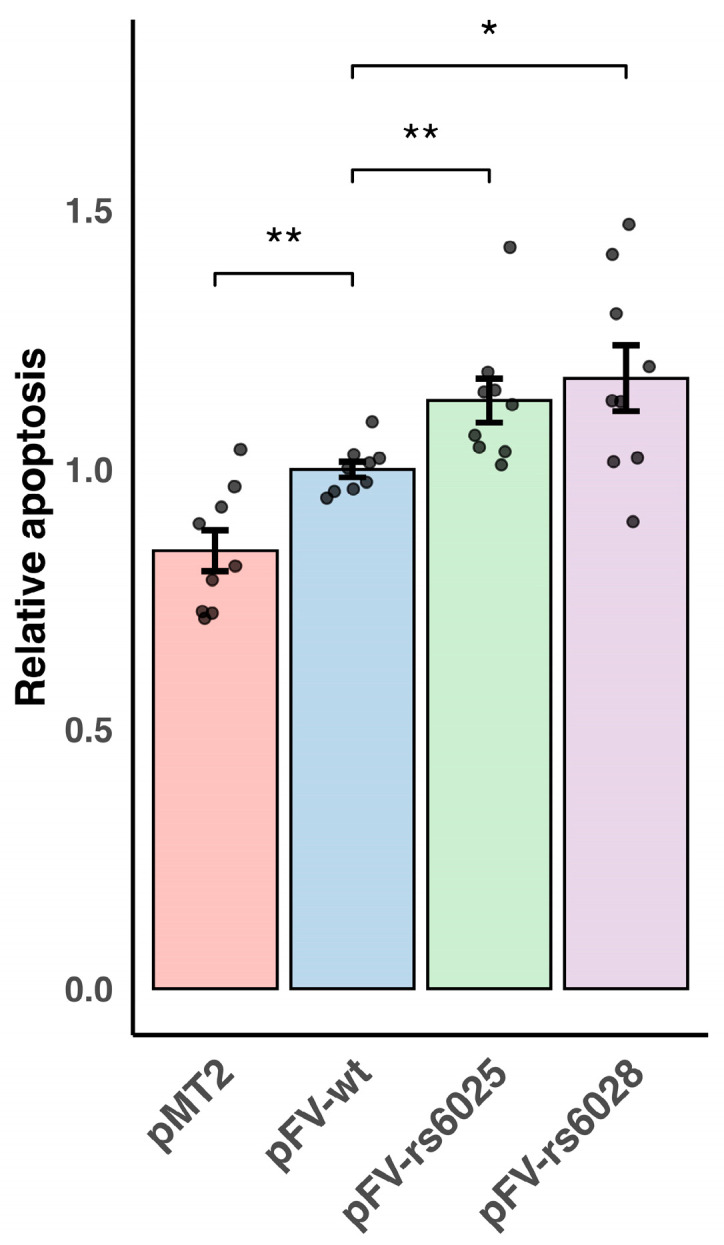 Figure 2
Figure 2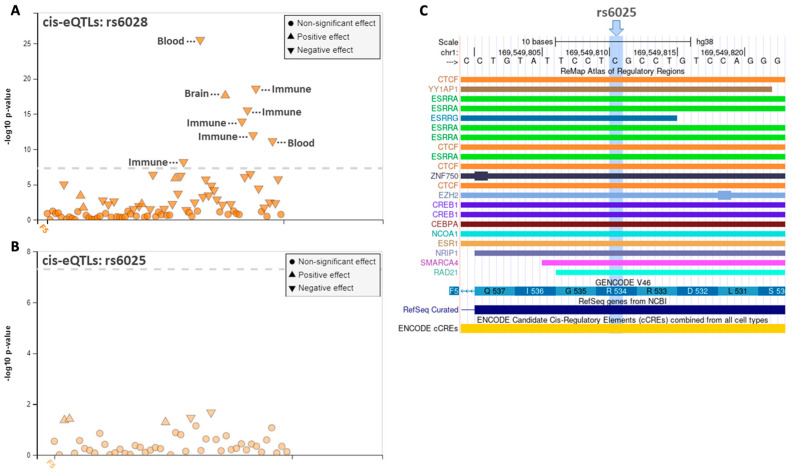 Figure 3
Figure 3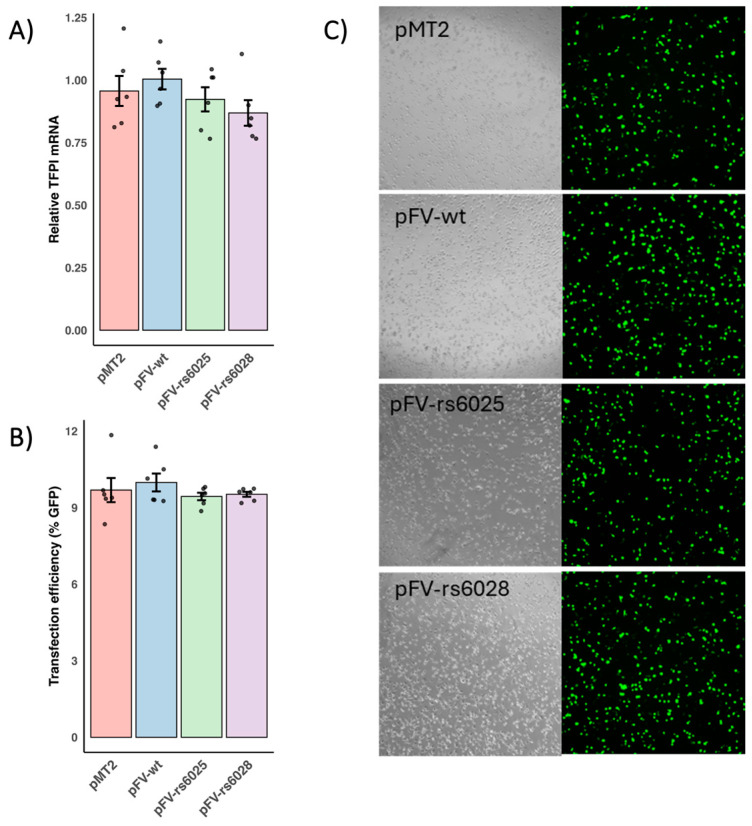 Figure 4
Figure 4- —Oslo University Hospital, Oslo, Norway
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBlood Coagulation and Thrombosis Mechanisms · Cancer-related gene regulation · Myeloproliferative Neoplasms: Diagnosis and Treatment
1. Introduction
The close relationship between cancer and the hemostatic system is well established [1]. Cancer patients are at increased risk of developing thrombosis [2]. Moreover, the hemostatic system promotes cancer pathogenesis [1,3]. Coagulation factor V (FV) is a non-enzymatic coagulation factor which is synthesized in the liver and present in blood circulation as an inactive precursor. FV is crucial in secondary hemostasis, exhibiting both pro- and anticoagulant properties. The procoagulant state occurs when activated FV (FVa) acts as a cofactor for coagulation factor X, accelerating thrombin generation. Conversely, acting as an anticoagulant cofactor to activated protein C (APC) and protein S, FV is able to inactivate activated coagulation factor VIII [4]. Alternative roles of FV are emerging. We have reported that FV was elevated in breast cancer tumors, and that high F5 mRNA levels were associated with improved overall survival and treatment response to chemotherapy, as well as induced apoptosis and reduced proliferation in breast cancer cells [5,6].
Single nucleotide polymorphisms (SNPs) and mutations in the FV gene (F5) can influence the properties of FV and lead to thrombosis or bleeding [4]. Factor V Leiden (F5 rs6025) is an extensively studied point mutation (c.1601G>A; p.Arg534Gln) resulting in the substitution of glutamine with arginine at position 506 of FV [4]. F5 rs6025 affects a critical cleavage site for APC, leading to APC resistance [4], and weakens the interaction between tissue factor pathway inhibitor α and FV [7]. These mechanisms contribute to an elevated risk of thrombosis, with heterozygous carriers showing an approximately seven-fold increased risk and homozygous carriers an approximately twenty-fold increase [8]. Cancer patients with F5 rs6025 have a two-fold increased risk of thrombosis compared to cancer patients without F5 rs6025 [9]. Research exploring associations between F5 rs6025 and cancer risk has shown diverse results [10,11,12,13,14,15,16]. Of note, we have previously demonstrated an association between four intronic F5 SNPs and risk of breast cancer [16].
We identified the SNPs in high linkage disequilibrium (LD) with the four target SNPs and found strong LD with the synonymous SNP F5 rs6028, which was the only SNP in LD that was located in an exon. Although synonymous SNPs do not change the encoded amino acid, they can influence mRNA splicing, stability, and structure, as well as protein folding [17]. This positions F5 rs6028 as a promising candidate for investigating potential functional effects. To our knowledge, this SNP has not yet been studied in cancer, and the functional impact of F5 variants in general is scarcely investigated.
In this study, we aimed to gain a better understanding of functional and regulatory effects of F5 rs6025 and F5 rs6028 in breast cancer cells.
2. Materials and Methods
2.1. Cell Culture
The triple-negative human breast epithelial adenocarcinoma cell line MDA-MB-231 (American Type Culture Collection) was cultured in Dulbecco’s Modified Eagle Medium (Gibco, Waltham, MA, USA) containing 4.5 g/L glucose and L-glutamine and 10% fetal bovine serum (Biowest, Nuaillé, France). The cells were maintained at 37 °C in a humidified atmosphere with 5% CO_2_.
2.2. Expression Vector and Mutagenesis
The pMT2-FV wild type (pFV-wt) [5] was used as a template to create plasmids with F5 rs6025 or F5 rs6028 using the QuikChange II XL Site-Directed Mutagenesis Kit (Agilent Technologies, Santa Clara, CA, USA). F5 rs6025 was introduced in a plasmid named pMT2-FV-rs6025 (pFV-rs6025) with the following primers: forward 5′-GCAGATCCCTGGACAGGCAAGGAATACAGAGGGCAGC-3′ and reverse 5′-GCTGCCCTCTGTATTCCTTGCCTGTCCAGGGATCTGC-3′. F5 rs6028 was introduced in a plasmid named pMT2-FV-rs6028 (pFV-rs6028) with the following primers: forward 5′-GAAAAACCACAGTCTACCATTTCAGGACTTCTTGGGCC-3′ and reverse 5′-GGCCCAAGAAGTCCTGAAATGGTAGACTGTGGTTTTTC-3′. The mutagenesis was verified with Sanger sequencing.
2.3. Transient Transfection and Harvesting of mRNA and Protein Lysates
MDA-MB-231 cells seeded in 6-well plates were transiently transfected using Lipofectamine™ 3000 (Thermo Fisher Scientific, Waltham, MA, USA) with 2.5 µg pMT2 empty vector, pFV-wt, pFV-rs6025, or pFV-rs6028. Transfection efficiency for the plasmids was tested by analyzing green fluorescent protein and tissue factor pathway inhibitor mRNA expression. Cells and media were harvested 48 h post transfection. Total RNA was isolated from cell lysates using the RNAqueous Total RNA Isolation Kit (Thermo Fisher Scientific). Cell lysates for protein analyses were harvested using RIPA buffer (Sigma-Aldrich, St. Louis, MO, USA) with 1xHalt Proteinase and Phosphatase Inhibitor (Thermo Fisher Scientific).
2.4. Quantitative Measurement of F5 mRNA
Total RNA was quantified with NanoDrop^®^ND-1000 and cDNA was synthesized using the High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific). Quantification of mRNA was performed with quantitative RT-PCR (qPCR) using the QuantStudio 12k Flex Real-Time system with the TaqMan Gene Expression Master Mix (Thermo Fisher Scientific) and TaqMan assays for F5 (Hs00914120_m1; Thermo Fisher Scientific) and Phosphomannomutase 1 (PMM1; in house) as endogenous control. Relative mRNA quantification (RQ) values for the F5 variants compared to pFV-wt was calculated using the comparative CT (ΔΔCT) method.
2.5. Measurement of FV Protein and FV Coagulant Activity
In cell media, FV protein was measured using ZYMUTEST Factor V (Hyphen-BioMed, Neuville-sur-Oise, France). The absorbance was measured with a VersaMax microplate reader (Emerson, St. Louis, MO, USA), and the SoftMax Pro 6.4 software was used to calculate protein concentrations. FV protein values in cell media were corrected for total protein concentrations in cell lysates, which was measured using the PIERCE BCA Protein Assay Kit (Thermo Fisher Scientific). In cell media, measurement of FV coagulant activity was performed as a one-stage clotting assay with prothrombin RecombiPlasTin^®^2G (Werfen, Barcelona, Spain) as initiating reagent on an ACL TOP 700 (Instrumentation Laboratory, Bedford, MA, USA). The values for the F5 variants were calculated relative to pFV-wt.
2.6. Detection of Apoptosis
Apoptosis was determined by quantification of nucleosomes in fresh cell lysates using the Cell Death Detection ELISA^PLUS^ kit (Roche Applied Science, Penzberg, Germany). The absorbance was measured using a VersaMax microplate reader. The values for the F5 variants were calculated relative to pFV-wt.
2.7. In Silico Analyses
Putative transcription factor binding sites overlapping with the positions of the F5 variants were studied using Remap 2022 [18] in the UCSC Genome Browser [19]. Remap provides data from ChIP-Seq experiments. Expression quantitative trait locus (eQTL) analysis (±1M base pair) was performed using FIVEx [20]. FIVEx is an interactive eQTL browser comprising the Gene-Tissue Expression (GTEx) data for visualization and functional interpretation of gene variants. FIVEx offers an intuitive interface that visualizes p-values alongside negative and positive effect sizes, representing decreased and increased expression for the alternative allele, respectively.
2.8. Statistical Analyses
Normality of the data was tested using the Shapiro–Wilks test. Statistics were calculated using the Kruskal–Wallis test followed by the Wilcoxon rank sum test. p-values < 0.05 were considered statistically significant.
3. Results and Discussion
F5 expression and F5 variants have been linked to breast cancer pathogenesis [6]. However, conflicting results are reported for the association between F5 rs6025 and cancer risk. Vossen et al. reported that homozygous carriers of F5 rs6025 showed an increased risk of colorectal cancer, whereas heterozygous carriers showed a decreased risk [10]. However, several studies with F5 rs6025 heterozygotes have failed to find associations to colorectal, gastric, gastrointestinal, oral, and gynecological cancers [11,12,13,14,15]. Similarly, we were not able to find an association between F5 rs6025 heterozygotes and breast cancer risk [16]. It is challenging to perform studies with sufficient statistical power when the frequency of a variant is low. Of note, only about 0.02% of the Caucasian population is F5 rs6025 homozygotes [21]. In vitro studies in cell lines are an alternative strategy to study direct effects of F5 variants in cancer. In this study, we aimed to explore functional and potential regulatory effects of the F5 rs6025 and F5 rs6028 variants in breast cancer.
3.1. Functional Effects of F5 Variants on FV Production, Secretion, and Coagulant Activity
We studied the effects of the F5 variants on F5 mRNA, FV protein, and FV coagulant activity. Transfection efficiency was equal for all plasmids (Figure A1). F5 mRNA and protein were significantly elevated in cells transfected with pFV-wt compared to the empty vector pMT2 (P = 8.2 × 10^−5^ and P = 9.1 × 10^−3^, respectively). Compared to pFV-wt, we observed a 23% and 53% increase in F5 mRNA levels for the pFV-rs6025 (P = 4.0 × 10^−2^) and pFV-rs6028 (P = 4.0 × 10^−2^) transfected cells, respectively (Figure 1A). Supporting this, FV protein levels in cell media were increased by 34% and 23% for pFV-rs6025 (P = 1.9 × 10^−2^) and pFV-rs6028 (P = 4.0 × 10^−3^), respectively (Figure 1B). These results showed that the elevated F5 mRNA expression for the F5 variants resulted in increased secretion of FV to cell media. Furthermore, coagulant activity of FV in cell media was elevated by 16% and 10% in cells transfected with pFV-rs6025 (P = 2.8 × 10^−3^) and pFV-rs6028 (P = 9.2 × 10^−3^), respectively, compared to pFV-wt (Figure 1C), which show that the increase in secreted FV protein was coagulant active in both variants. Supporting this result, we have, of note, observed elevated plasma D-dimer levels in F5 rs6025 carriers compared to non-carriers in breast cancer patients, which indicated increased coagulant activity in F5 rs6025 carriers with breast cancer. Differences in the transfection efficiency could correlate with increased mRNA and protein expression observed for the F5 variants compared to F5 wild type. We conducted control experiments with co-transfection of the F5 variant plasmids and a control plasmid with equal amounts of either GFP or TFPI plasmids. The results reflected equal transfection efficiency across the F5 plasmids (Figure A1). We can then assume that the differences we observed between the F5 variants are not because of differences in transfection efficiency, but specifically because of the F5 variants.
3.2. Functional Effects of F5 Variants on Apoptosis
Apoptosis was significantly elevated in cells transfected with pFV-wt compared to the empty vector pMT2 (P = 7.8 × 10^−3^). Compared to pFV-wt, we observed a modest 13% increase in apoptosis in cells transfected with pFV-rs6025 (P = 1.9 × 10^−3^) and an 18% increase in cells transfected with pFV-rs6028 (P = 2.4 × 10^−2^) (Figure 2). We have previously reported that overexpression of the FV wild type induced apoptosis in MDA-MB-231 cells [5]. The increased effect on apoptosis observed in cells transfected with pFV-rs6025 and pFV-rs6028, compared to pFV-wt, may therefore be due to the higher levels of FV per se, rather than a novel function caused by the F5 variants. The results show that the F5 variants enhances tumor suppressor functions, possibly via increasing the FV expression, which might be relevant in breast cancer pathology. Interestingly, we have previously reported that F5 rs9332542, the SNP in strong LD with F5 rs6028, was associated with decreased risk of breast cancer which might support these results. While this study focused on a triple-negative cell line, the F5 variants may exert different apoptotic effects in other breast cancer cell types, such as ER-positive tumors, which typically express lower levels of FV. Further research is needed to determine whether these effects vary across different breast cancer subtypes.
3.3. Regulatory Annotation Analysis of the F5 Variants
Considering the elevated expression observed for the F5 variants compared to pFV-wt, we explored the possible regulatory effect of the F5 variants by analyzing the associations between the F5 variants and known eQTLs. We searched for eQTLs located near the gene of origin (cis-eQTLs) using the FIVEx tool [20]. F5 rs6028 was identified as a cis-eQTL for F5 expression with the minor allele having an opposite effect on the gene expression in brain and blood/immune cells, which might indicate a tissue specific effect of the SNP (Figure 3A). The cis-eQTL associations for F5 rs6025 and F5 expression were not significant after adjusting for multiple testing (Figure 3B).
The functional annotation analyses indicated that F5 rs6028 may act directly or indirectly as a transcriptional regulator for F5 in several tissues. This might explain the increased FV expression observed for pFV-rs6028 compared to pFV-wt. Supporting this, we have previously reported that F5 rs9332542, the SNP in strong LD with F5 rs6028, was correlated to FV mRNA and protein expression in plasma from breast cancer patients [6].
Since we observed a differential F5 expression for the F5 variants relative to the wild type, our hypothesis is that these variants might be located in transcription factor binding sites. This might influence putative binding sites for transcription which potentially can influence functional effects on F5 expression and apoptosis. To explore this, we used data from ChIP-Seq experimental data from Remap [18]. No ChIP-Seq peaks were observed for the location of F5 rs6028. In the location of F5 rs6025, the most profound ChIP-Seq peaks were for the transcription factors Estrogen Related Receptor Alpha (ESRRA) (n = 5) and CCCTC-binding factor (CTCF) (n = 4) (Figure 3C). The ChIP-Seq peaks for ESRRA were observed in three different breast cancer cell lines and lymphoblasts, whereas the ChIP-Seq peaks for CTCF were observed in B-cells, keratinocytes, cardiac cells, and epithelial cells. Stein et al. have reported that Estrogen-related receptor alpha (ERRα) (encoded by the ESRRA gene) is critical for growth and migration of estrogen receptor-negative cells [22], and it has been reported that CTCF upregulation reduces apoptosis in breast cancer cells [23,24]. The direct effects of F5 rs6025 on these putative transcription factor binding sites remain to be proven. However, the results suggest that F5 rs6025 might play a role in transcription regulation, potentially influencing processes like cell growth, migration, or apoptosis.
In this study, we used transient transfection of breast cancer cell lines. Using this approach, we can study a high overexpression of the different variants we want to study, but we can only study short-time effects. The efficiency of transfection between the different variant plasmids may influence the measured outcomes. We conducted co-transfection experiments with the F5 variant plasmids together with GFP and TFPI plasmids, and the results from these experiments show no indirect differences in the efficiency between the F5 variant plasmids, although an absolute normalization of the transfection efficiency could not be obtained by this method.
Collectively, our study demonstrated functional and regulatory effects of F5 rs6025 and F5 rs6028 in breast cancer cells. Our findings suggest that these F5 variants may enhance the tumor suppressor functions in breast cancer, potentially due to increased FV expression.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Falanga A. Marchetti M. Vignoli A. Coagulation and cancer: Biological and clinical aspects J. Thromb. Haemost.20131122323310.1111/jth.1207523279708 · doi ↗ · pubmed ↗
- 2Soff G. Thrombosis and Hemostasis in Cancer Springer International Publishing Cham, Switzerland 2019
- 3Hisada Y. Mackman N. Tissue factor and cancer: Regulation, tumor growth, and metastasis Seminars in Thrombosis and Hemostasis Thieme Medical Publishers New York, NY, USA 201910.1055/s-0039-1687894 PMC 654651931096306 · doi ↗ · pubmed ↗
- 4Cramer T.J. Griffin J.H. Gale A.J. Factor V is an anticoagulant cofactor for activated protein C during inactivation of factor Va Pathophysiol. Haemost. Thromb.201037172310.1159/00031514120501981 PMC 2974842 · doi ↗ · pubmed ↗
- 5Lind S.M. Sletten M. Hellenes M. Mathelier A. Tekpli X. Tinholt M. Iversen N. Coagulation factor V in breast cancer: A p 53 regulated tumor suppressor and predictive marker for treatment response to chemotherapy J. Thromb. Haemost.2024221569158210.1016/j.jtha.2024.02.00838382738 · doi ↗ · pubmed ↗
- 6Tinholt M. GarredØ. Borgen E. Beraki E. Schlichting E. Kristensen V. Sahlberg K. Iversen N. Subtype-specific clinical and prognostic relevance of tumor-expressed F 5 and regulatory F 5 variants in breast cancer: The Co Ca V study J. Thromb. Haemost.2018161347135610.1111/jth.1415129766637 · doi ↗ · pubmed ↗
- 7Mast A.E. Ruf W. Regulation of coagulation by tissue factor pathway inhibitor: Implications for hemophilia therapy J. Thromb. Haemost.2022201290130010.1111/jth.1569735279938 PMC 9314982 · doi ↗ · pubmed ↗
- 8Albagoush S.A. Koya S. Chakraborty R.K. Schmidt A.E. Factor V Leiden Mutation, in Stat Pearls Stat Pearls Publishing St. Petersburg, FL, USA 202330521223 · pubmed ↗