A Subset of HOX Genes Negatively Correlates with HOX/PBX Inhibitor Target Gene Expression and Is Associated with Apoptosis, DNA Repair, and Metabolism in Prostate Cancer
Richard Morgan, Christopher Smith, Hardev Pandha

TL;DR
Some HOX genes are linked to cancer growth pathways and reduced cell adhesion in prostate cancer.
Contribution
Identifies a specific HOX gene subset correlated with cancer-related pathways and patient age in prostate cancer.
Findings
A subgroup of HOX genes negatively correlates with Fos, DUSP1, and ATF3 expression in prostate cancer.
These HOX genes correlate with DNA repair and aminoacyl tRNA biosynthesis pathways.
The HOX gene subset shows negative correlation with cell adhesion-promoting genes.
Abstract
Background/Objectives: The HOX genes encode a family of homeodomain-containing transcription factors that have important roles in defining cell and tissue identity in embryonic development, but which also show deregulated expression in many cancers and have been shown to have pro-oncogenic roles. Due to their functionally redundant nature, strategies to target HOX protein function in cancer have focused on their interaction with their PBX cofactor using competitive peptides such as HXR9. HOX/PBX inhibition triggers apoptosis through a sudden increase in target gene expression, including Fos, DUSP1, and ATF3, which are otherwise repressed by HOX/PBX binding. Methods: We analyzed publicly available transcriptomic data in the R2 platform. Results: We show that a specific subgroup of HOX genes is negatively correlated with Fos, DUSP1, and ATF3 expression in prostate cancer, and that this…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
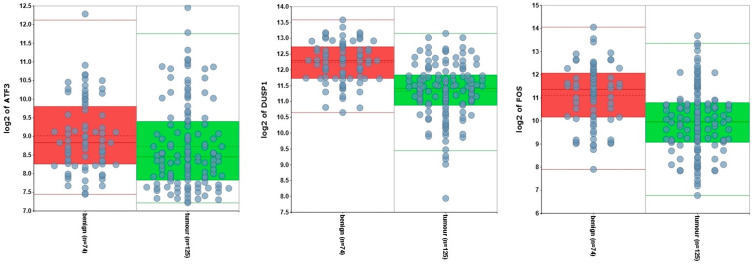 Figure 1
Figure 1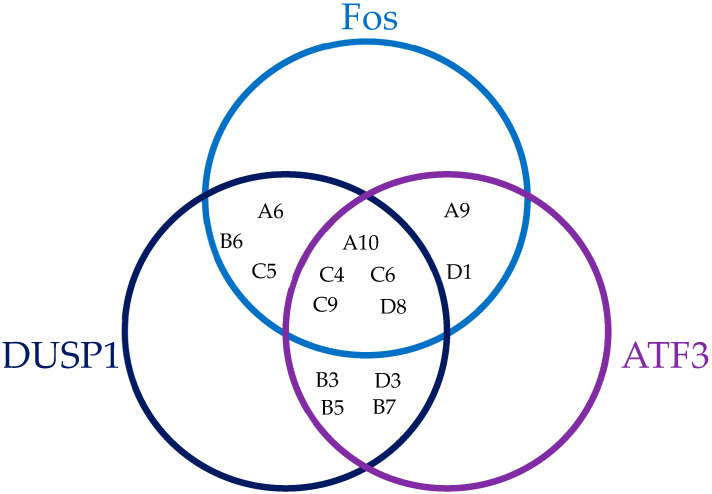 Figure 2
Figure 2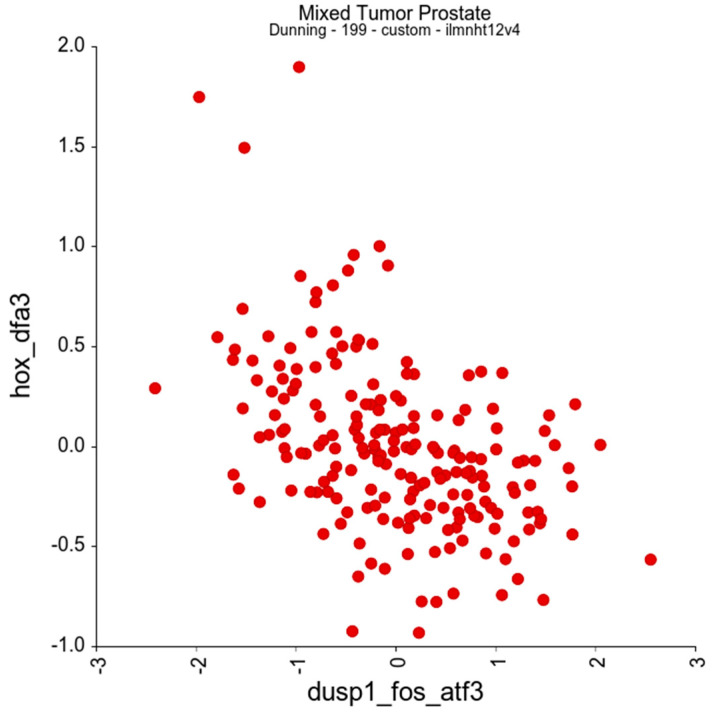 Figure 3
Figure 3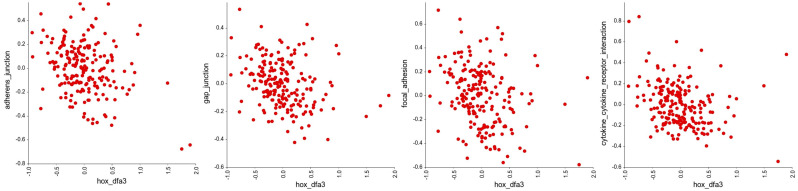 Figure 4
Figure 4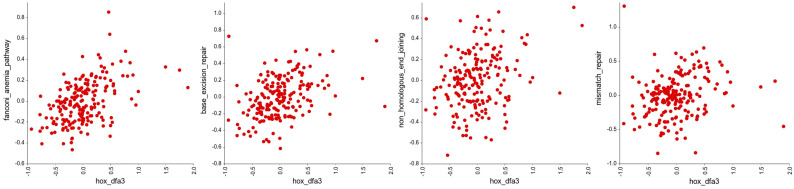 Figure 5
Figure 5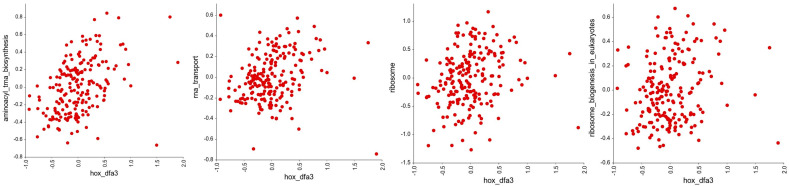 Figure 6
Figure 6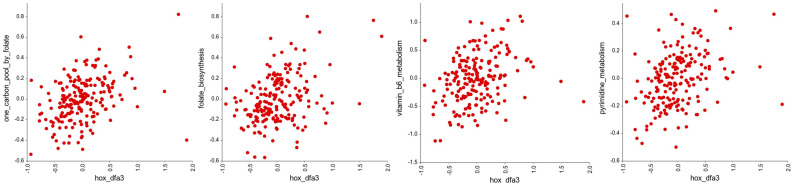 Figure 7
Figure 7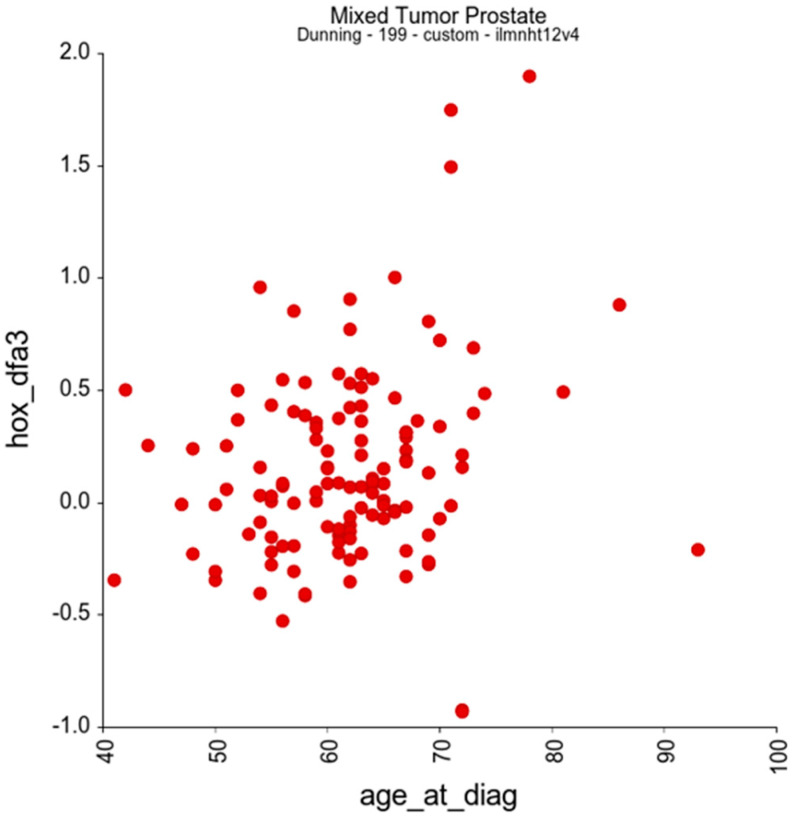 Figure 8
Figure 8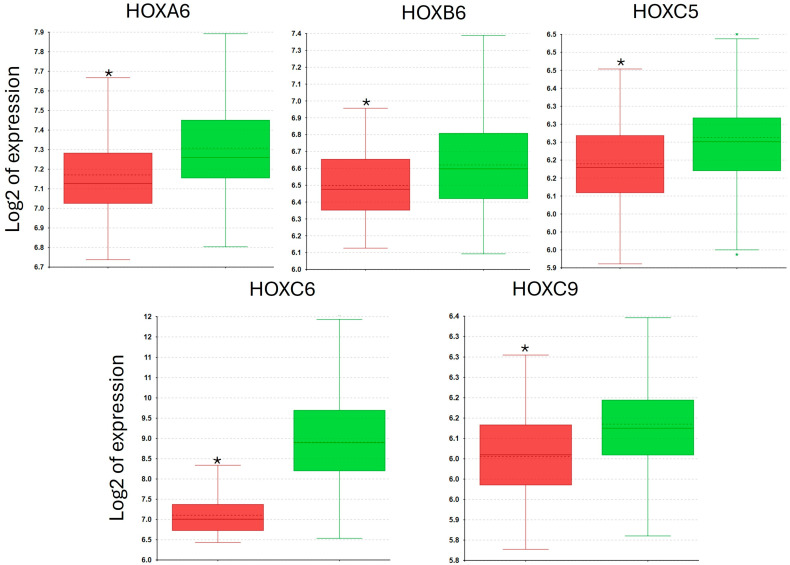 Figure 9
Figure 9Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsGenetic and Clinical Aspects of Sex Determination and Chromosomal Abnormalities · Prostate Cancer Treatment and Research · RNA Research and Splicing
1. Introduction
The HOX genes encode a family of transcription factors that have key roles in development [1] and in adult processes that involve the renewal of cells, including the proliferation of haematopoietic stem cells and the subsequent differentiation of blood cell lineages [2]. Central to this is the ability of HOX genes to confer specific identities on cells and tissues, which, combined with their expression in spatial and temporal gradients in the embryo, gives them a key role in embryonic patterning [1].
In addition to their role in development, HOX genes are also significantly deregulated in cancer, and are frequently upregulated in most tumour types [3]. Their role in cancer is primarily pro-oncogenic, based on functional (rather than associative) studies, and includes the promotion of cell survival and proliferation, angiogenesis, metastasis, and immune invasion [3]. A major barrier to targeting HOX proteins is their functional redundancy: the four clusters of vertebrate HOX genes (A, B, C, and D) arose by the duplication of the ancestral HOX cluster found in insects, and paralog genes that share a common ancestor gene (for example, HOXA4, HOXB4, HOXC4, and HOXD4) have high levels of similarity, as do neighbouring genes within clusters (for example HOXB4, HOXB5, and HOXB6). Consequently, multiple HOX proteins must be knocked down or inhibited to result in a phenotypic effect [4]. One solution to this has been to target the interaction between HOX proteins encoded by HOX genes in paralog groups 1 through 10, and their common cofactor, PBX. PBX binding modifies the way in which HOX proteins bind to DNA and can change their intracellular distribution, causing them to relocate to the nucleus. This interaction can be inhibited using a peptide mimic of the conserved hexapeptide region in HOX proteins that mediates binding to PBX. The most frequently used inhibitor peptide, HXR9, contains a polyarginine sequence that facilitates cellular uptake, and has been shown to cause apoptosis in a wide range of solid malignancies [5].
HXR9-induced apoptosis is mediated by the loss of inhibition of three key genes—Fos, Dual Specificity Phosphatase 1 (DUSP1), and Activating Transcription Factor 3 (ATF3), all of which can trigger cell death when expressed at high levels [6,7,8,9,10]. In the case of Fos, this is primarily through the activation of the extracellular canonical apoptotic pathway via an increase in Fas Ligand (FASL) expression [11]. ATF3 also increases apoptosis by stabilising p53, leading to increased BAX expression [12]. DUSP1 inhibits Epidermal Growth Factor Receptor (EGFR) signalling through the dephosphorylation of MEK and ERK, reducing cell proliferation and survival [13]. A key role of HOX genes in cancer thus seems to be the repression of Fos, DUSP1, and ATF3 expression, which in turn inhibits apoptosis, and correspondingly DUSP1 [14] and ATF3 [15] have both previously been identified as potential tumour suppressor genes in prostate cancer, although the role of Fos in this context is less clear.
In this bioinformatic analysis, we have identified HOX genes that show a negative correlation with Fos, DUSP1, and ATF3 expression in prostate tumours. The expression of this subset of HOX genes is also strongly correlated with that of genes involved in DNA repair, metabolism, and motility, indicating that they have additional pro-oncogenic roles. Furthermore, they show a significant increase in tumour expression in older patients, reflecting previous findings that HOX genes become progressively more highly expressed with age in some individuals.
2. Methods
2.1. Microarray Data
The dataset used in this analysis is from the study by Ross-Adams et al., in which 103 samples from men with prostate cancer and 99 matched benign samples were profiled using the Illumina HT12v4 expression array to provide transcriptomic data [16]. The clinical characteristics of this cohort are described in detail elsewhere, but the key clinical parameters included are age at diagnosis (range, 41–93 years), biochemical relapse, clinical stage, Gleason grade, extra capsular extension, positive surgical margins, PSA at diagnosis, and total follow up time (range, 2–67 months). Bead-level data were processed to remove spatial artefacts and then log2-transformed and quantile-normalised. Probes that did not match their intended genomic location or which mapped to genomic areas that were not useful for transcription studies were excluded from the analysis. For validation, the analyses were repeated on a second, independent dataset [17]. This contains transcriptomes from 131 primary prostate tumours, 29 matched normal tissues, and 19 metastases from patients with a mean age of 58 years at diagnosis (range, 37–83 years).
2.2. Data Analysis
Data analysis was performed using the R2 platform (R2: Genomics Analysis and Visualization Platform (http://r2.amc.nl, accessed on 1 July 2025)). In order to generate the HOX_DFA3 gene set, individual gene vs. gene correlations were performed for each of the putative target genes (Fos, DUSP1, and ATF3) with each of the HOX genes in paralog groups 1–10 (30 HOX genes in total) using the “Correlate 2 genes” function, and HOX genes that negatively correlated with one or more of the target genes with p < 0.05 and r < −0.1 were included in the HOX_DFA3 group. For gene set vs. gene set correlations, R2 was used to perform a regression analysis for the mean value of the genes in the HOX_DFA3 gene set against other defined gene sets (KEGG) for each sample, with transformation based on the Z-score. For correlations between the HOX_DFA3 gene set and patient age at diagnosis, the Track (#) vs. Genesets Correlations function was used, again with the Z-score transformation.
For individual HOX genes, we compared expression with specific clinical and pathological criteria using the gene vs. track function and a log2 transformation, using a 1-way ANOVA to test for significance (p < 0.0033, applying the Bonferroni correction to account for the 14 different HOX genes in the HOX_DFA3 gene set).
3. Results
3.1. DUSP1 and FOS Are Both Significantly More Highly Expressed in Benign Prostate Tissue Compared to Tumour Tissue
Given the potential tumour suppressor roles of DUSP1, FOS, and ATF3, we compared the expression of these genes in prostate tumours and benign prostate tissue. This reveals that all three of these genes are expressed more highly in benign tumours (Figure 1), although only significantly so in the case of DUSP1 (p = 2.02 × 10^−12^) and Fos (p = 1.18 × 10^−7^).
3.2. A Subset of HOX Genes Negatively Correlate with DUSP1, Fos, and ATF3 Expression
As described above, DUSP1, Fos, and ATF3 are chosen as a starting point because previous studies showed that their upregulation mediated the apoptosis induced by the HOX/PBX antagonist HXR9. Hence, the first part of this study seeks to identify HOX genes that show a negative association with the expression of ATF3 and/or DUSP1 and/or Fos in order to determine a putative set of HOX genes that may be responsible for the cytotoxicity of HXR9. To establish which HOX genes showed an mRNA expression that was negatively correlated with Fos, DUSP1, or ATF3, we compared their individual expression to that of each HOX gene in paralog groups 1–10 through a regression analysis (Table 1), with a cutoff value of p < 0.05 and r < −0.1. This indicates that 14 of the 30 HOX genes in these paralog groups are significantly correlated with the expression of at least one of Fos, DUSP1, or ATF3, and that 5 HOX genes (HOXA10, HOXC4, HOXC6, HOXC9, and HOXD8) are significantly correlated with the expression of all three (Figure 2).
On this basis, we can define a subset of HOX genes which are negatively correlated with DUSP1 and/or Fos and/or ATF3—HOXA6, HOXA9, HOXA10, HOXB3, HOXB5, HOXB6, HOXB7, HOXC4, HOXC5, HOXC6, HOXC9, HOXD1, HOXD3, and HOXD8. For subsequent analyses, these are treated as a single gene set, referred to as DFA3_HOX. We then performed a correlation analysis between DFA3_HOX and DUSP1, Fos, and ATF3 as a single gene set, referred to as DUSP1_FOS_ATF3 subsequently. This reveals a very strong negative association between DFA3_HOX and their putative target genes, DUSP1_FOS_ATF3, with an R-value of −0.469 and a p-value of 2.70 × 10^−12^ (Figure 3).
3.3. DFA3_HOX Expression Correlates with the Genes Involved in Cell Adhesion, DNA Repair, Cell Metabolism, and Translation
Next, we looked to see if the HOX genes in DFA3_HOX had significant associations with other gene groups in the KEGG pathways of defined cellular processes using an R-value cutoff of <−0.2 and >0.2. This reveals a significant negative correlation with the expression of genes involved in cell adhesion (Figure 4, Supplementary Figure S1), and a significant positive correlation with the expression of genes involved in DNA repair (Figure 5, Supplementary Figure S2), translation (Figure 6, Supplementary Figure S3), and cellular metabolism (Figure 7, Supplementary Figure S4). Amongst these, the strongest correlations were between DFA3_HOX and genes involved in the Fanconi Anemia Pathway (r = 0.407, p = 2.49 × 10^−9^), base excision repair (r = 0.382, p = 2.48 × 10^−8^), and aminoacyl tRNA synthesis (r = 0.425, p = 4.02 × 10^−10^).
In addition to the relationships between gene sets, we looked to see if there was a relationship between the DFA3_HOX gene set and clinical characteristics of the patient cohort. This revealed only one potentially significant positive correlation, which was between DFA_HOX3 expression and patient age at diagnosis, with an R-value of 0.22 and p-value of 0.014 (Figure 8).
3.4. Individual HOX_DFA3 Gene Expression in Tumour vs. Normal Tissue
We also assessed whether any of the individual genes in the HOX_DFA3 gene set showed a significantly different level of expression between normal prostate tissue and tumours, with a cutoff p-value of 0.0033 (applying the Bonferroni correction with respect to the 14 HOX genes in this group). Comparing benign to tumour samples, HOXA6 (p = 4.26 × 10^−5^), HOXB6 (p = 0.001), HOXC5 (p = 4.69 × 10^−5^), HOXC6 (p = 3.02 × 10^−28^), and HOXC9 (p = 0.0001) all showed significantly greater expression in tumour samples, with HOXC6 having by far the greater fold difference (Figure 9). In addition, we performed a heat map analysis for all the HOX genes in HOX_DFA3, which reveals that most normal prostate tissue samples have significantly lower expression of these genes compared to tumour tissue (Supplementary Figure S5).
3.5. Validation with Independent Datasets
To determine whether the findings from our analysis are applicable beyond the Ross-Adams dataset, we repeated the analyses on data from a second, independent study. This second dataset contains transcriptomes from a total of 131 primary prostate tumours, 19 metastases, and 29 normal prostate tissue samples [17]. This revealed similarly significant relationships between HOX_DFA3 and the other gene sets described above (Table 2).
4. Discussion
In this analysis of HOX gene expression in prostate tumours and matched benign tissue, we have identified a subset of 14 HOX genes, DFA3_HOX, the expression of which is negatively correlated with ATF3, DUSP1, and Fos. These three genes have previously been identified as targets of HOX/PBX-mediated transcriptional repression, which in turn is a key survival mechanism in at least some types of cancer cells, as they promote apoptosis through both intrinsic and extrinsic pathways. This strong negative correlation between DFA3_HOX and ATF3, DUSP1, and Fos, therefore, supports the hypothesis based on in vitro data and mouse models that HOX genes have an important role in preventing apoptosis.
Our analysis also revealed that this subset of HOX genes has a strong negative correlation with genes involved in cell adhesion, specifically those that encode proteins in gap, focal adhesion, and adherens junctions. This concurs with previous studies that have shown a relationship between HOX genes and cell migration, and, in particular, with the epithelial to mesenchymal transition (EMT) [3]. Multiple HOX genes are involved in EMT, and there is evidence of redundancy in their function—for example, HOXB7, HOXB8, HOXC8, HOXC10, and HOXD3 all repress E-cadherin expression [18,19,20,21,22,23,24,25], and similarly, HOXB8 and HOXD3 promote N-cadherin expression [20,21,26,27].
The DFA3_HOX genes also show a strong positive correlation with genes involved in pro-oncogenic processes—DNA repair, translation, and cell metabolism. Multiple HOX genes have previously been shown to have roles in DNA repair, for example, HOXB7 directly interacts with components involved in double-stranded break repair (DSB) [28], and HOXB9 [29] and HOXA10 [30] also promote DSB. Conversely, there do not seem to be any previous studies identifying HOX transcription factors as regulators of genes involved in translation and cell metabolism, although the generally reported pro-oncogenic role for HOX genes is consistent with the association reported here [3].
In addition to these associations, we also identified a significant positive correlation between patient age at diagnosis and expression of DFA3_HOX genes with a regression value of 0.22 (p = 0.014). A potential relationship between HOXA9 expression in normal peripheral blood cells and age has previously been reported [31], in which HOXA9 expression was relatively low until the age of 60 years, at which point there was a significant increase in mean expression (p < 0.0001). Subsequent studies have indicated that promoter methylation may play a role in age-related changes in HOX gene expression. For example, HOXA5 (which is considered to have a tumour-suppressor rather than pro-oncogenic role) undergoes increasing promoter methylation with age and a corresponding loss of expression at both the RNA and protein level [32], and in a study of muscle cells from young (27 ± 4.4 years) and old (83 ± 4 years) healthy volunteers, significantly increased methylation of regulatory regions was shown for multiple HOX genes [33].
Taken together, these findings support the previously reported pro-oncogenic role for HOX genes [3], which includes, for the DFA3_HOX subgroup, the suppression of pro-apoptotic genes and genes that inhibit cell movement, and the transcriptional activation of the genes involved in DNA repair, translation, and metabolism. Several of these potential regulatory interactions, especially with respect to translation and metabolism, have not previously been reported, and their mechanistic basis requires further investigation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Mallo M. Wellik D.M. Deschamps J. Hox Genes and Regional Patterning of the Vertebrate Body Plan Dev. Biol.201034471510.1016/j.ydbio.2010.04.02420435029 PMC 2909379 · doi ↗ · pubmed ↗
- 2Alharbi R.A. Pettengell R. Pandha H.S. Morgan R. The Role of HOX Genes in Normal Hematopoiesis and Acute Leukemia Leukemia 2013271000100810.1038/leu.2012.35623212154 · doi ↗ · pubmed ↗
- 3Morgan R. Hunter K. Pandha H.S. Downstream of the HOX Genes: Explaining Conflicting Tumour Suppressor and Oncogenic Functions in Cancer Int. J. Cancer 20221501919193210.1002/ijc.3394935080776 PMC 9304284 · doi ↗ · pubmed ↗
- 4Drake K.A. Adam M. Mahoney R. Potter S.S. Disruption of Hox 9,10,11 Function Results in Cellular Level Lineage Infidelity in the Kidney Sci. Rep.20188630610.1038/s 41598-018-24782-529679048 PMC 5910417 · doi ↗ · pubmed ↗
- 5Morgan R. El-Tanani M. Hunter K.D. Harrington K.J. Pandha H.S. Targeting HOX/PBX Dimers in Cancer Oncotarget 20178323223233110.18632/oncotarget.1597128423659 PMC 5458287 · doi ↗ · pubmed ↗
- 6Morgan R. Pirard P.M. Shears L. Sohal J. Pettengell R. Pandha H.S. Antagonism of HOX/PBX Dimer Formation Blocks the in Vivo Proliferation of Melanoma Cancer Res.2007675806581310.1158/0008-5472.CAN-06-423117575148 · doi ↗ · pubmed ↗
- 7Kelly Z.L. Michael A. Butler-Manuel S. Pandha H.S. Morgan R.G. HOX Genes in Ovarian Cancer J. Ovarian Res.201141610.1186/1757-2215-4-1621906307 PMC 3179923 · doi ↗ · pubmed ↗
- 8Morgan R. Boxall A. Harrington K.J. Simpson G.R. Michael A. Pandha H.S. Targeting HOX Transcription Factors in Prostate Cancer BMC Urol.2014141710.1186/1471-2490-14-1724499138 PMC 3942264 · doi ↗ · pubmed ↗