Epidemiological Surveillance, Variability, and Evolution of Isolates Belonging to the Spanish Clone of the 4,[5],12:i:- Monophasic Variant of Salmonella enterica Serovar Typhimurium
Xenia Vázquez, Patricia García, Javier Fernández, Víctor Ladero, Carlos Rodríguez-Lucas, Jürgen J. Heinisch, Rosaura Rodicio, M. Rosario Rodicio

TL;DR
This study tracks the evolution and resistance patterns of a Salmonella clone in Spain over two decades, linking changes to antibiotic use policies and genetic factors.
Contribution
The study reveals how IS26 transposition events and plasmid instability drive genetic variability and resistance loss in a Salmonella clone.
Findings
Isolates clustered into two clades, A and B, with distinct temporal and genetic characteristics.
Antibiotic resistance genes decreased over time, possibly due to EU regulations on antibiotic use in feed.
IS26 transposition events are linked to both clonal relationships and the monophasic phenotype via fljAB deletions.
Abstract
Background/Objective: The present study focused on the analysis of the Spanish clone belonging to the successful 4,[5],12:i:- monophasic variant of Salmonella enterica serovar Typhimurium. Methods: All isolates of the clone recovered in a Spanish region from human clinical samples between 2008 and 2018 (N = 14) were investigated using microbiological approaches and genome sequence analysis. In addition, they were compared with isolates from the years 2000 to 2003 (N = 21), which were previously characterized but had not yet been sequenced. Results: Phylogenetic analyses indicate that all isolates are closely related (differing by 1 to 103 SNPs) but belong to two clades termed A and B. With few exceptions, clade A comprised isolates of the first period, also including two “older” control strains, LSP 389/97 and LSP 272/98. Clade B only contained isolates from the second period. Isolates…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
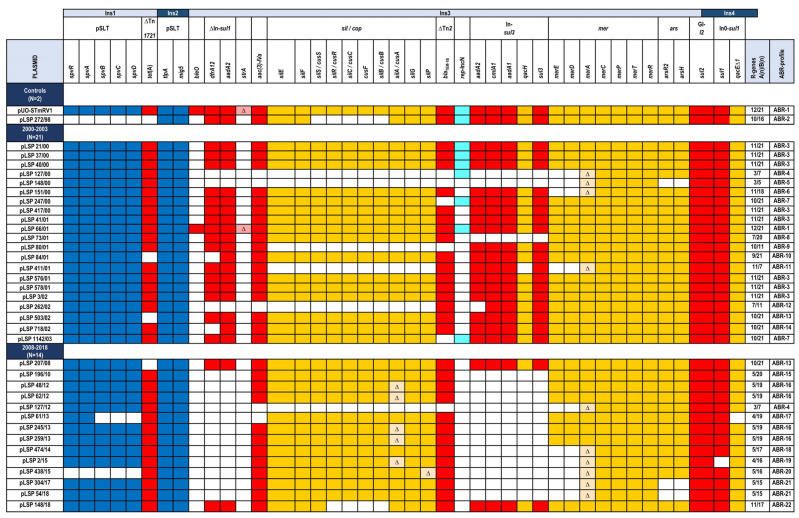 Figure 1
Figure 1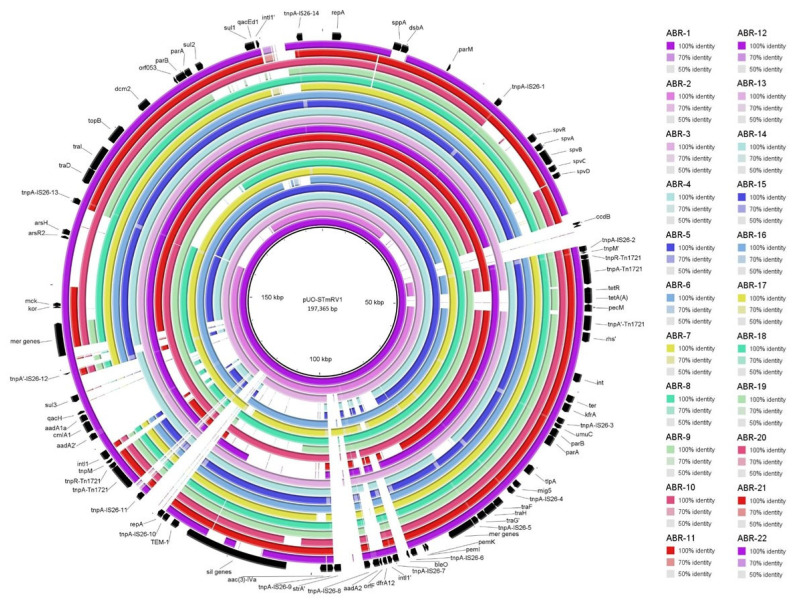 Figure 2
Figure 2 Figure 3
Figure 3 Figure 4
Figure 4- —“Fondo de Investigación Sanitaria, Instituto de Salud Carlos III, Ministerio de Economía y Competitividad”
- —European Regional Development Fund of the European Union
- —Program “Severo Ochoa”
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSalmonella and Campylobacter epidemiology · Antibiotic Resistance in Bacteria · Vibrio bacteria research studies
1. Introduction
The bacterial pathogen Salmonella enterica is one of the major causes of foodborne intestinal infections worldwide [1]. Although nearly 2600 serovars have been identified in this complex species [2], most cases of human salmonellosis worldwide can be attributed to just three of them, namely, S. Enteritidis, S. Typhimurium, and S. 4,[5],12:i:-. In fact, they accounted for 84.7% of all confirmed human infections in the European Union (EU) in 2022 [3]. Although S. 4,[5],12:i:- represents an independent serovar in terms of epidemiological surveillance [4], it actually is a monophasic variant of S. Typhimurium, which lacks the second-phase flagellar antigens (designated 1,2 in the antigenic formula of S. Typhimurium: 4,[5],12:i:1,2). This variant was responsible for 4.3% of the total cases of human salmonellosis confirmed in 2022 in the EU [5].
The 4,[5],12:i:- isolates investigated thus far can be assigned to four main clonal lines known as the Spanish, the European, the Southern European, and the United States clones [4,6,7]. These clones primarily differ both in their resistance towards antimicrobial drugs, with all except the U.S. clone being multidrug-resistant (MDR), and in the location of the encoding genes (residing either in specific chromosomal resistance regions or on plasmids of distinct incompatibility groups). They also differ by variable alterations affecting the fljAB-hin region, which encodes the second phase flagellin (fljB gene) and is responsible for phase variation in biphasic S. Typhimurium (governed by hin and fljA) [8]. Together with other characteristics of the monophasic clones, such as the sequence type (ST), predominant phage types, PFGE profiles, and phylogenetic relationships, this indicates that the monophasic clones originated from independent evolutionary events and do not necessarily derive from a common ancestor [9].
The Spanish monophasic clone emerged in Spain in the 1990s, recovered from pigs and pork products, and has been circulating mostly in the Iberian Peninsula [10,11,12,13,14,15,16,17]. Isolates of this clone show ST19 and were mainly associated with phage type U302, although DT193, DT208, and NT (not typeable with the available phage library) have also been reported [9,12,14]. The monophasic feature of the Spanish clone results from deletions within the fljAB region and its surrounding genes. Several deletions have already been identified, which were probably caused by transposition of the insertion sequence IS26. They all start at the same site within STM2758 (according to the orf nomenclature used for the annotation of the S. Typhimurium LT2 genome; accession number AE006468.1) but end at distinct positions [15,18,19,20].
The Spanish clone is typically resistant to ampicillin, chloramphenicol, gentamicin, streptomycin, sulphonamides, tetracyclines, and trimethoprim, although variations of the main phenotype have been detected [14,16,17]. The genes responsible for these resistances [blaTEM-1B, cmlA1, aac(3)-IV, aadA1, aadA2, sul1, sul2, sul3, tet(A), and dfrA12] are located on large plasmids (of ca. 150 to 200 kb) of the IncC (initially reported as IncA/C) incompatibility group which occasionally harbour the IncN replicon. Most of the resistance genes are supplied by elements involved in DNA mobility, including two class 1 integrons of the sul1-type, a class 1 integron of the sul3-type, and several transposons, specifically truncated versions of Tn2, Tn21, and Tn1721. One of the sul1 integrons carries the dfrA12-aadA2 genes in the variable region but lacks the 3′-conserved segment (CS) where the sul1-qacEΔ1 genes reside; the other sul1 integron does not contain gene cassettes in the variable region but retains an intact 3′-CS. On the other hand, the resistance genes aadA2, cmlA1, aadA1, and qacH are located within the sul3 integron. The IncC plasmids of the Spanish monophasic clone also contain the spv operon (spvRABCD) along with other genes derived from pSLT, the virulence plasmid specific for S. Typhimurium [14,17]. pUO-STmRV1 (197,365 bp), the first resistance–virulence plasmid reported in the Spanish monophasic clone, was selected as a representative of the group and fully sequenced employing second- and third-generation technologies (i.e., Illumina and PacBio) [21].
To broaden our knowledge of the Spanish monophasic clone and determine its variability, all isolates of this clone recovered from human clinical samples in Asturias between 2008 and 2018 were sequenced with Illumina technology and experimentally characterized in the present study. In addition, the results were compared with those from clinical isolates obtained in the same region between the years 2000 and 2003, which were experimentally investigated in previous studies [14,15] but not sequenced until now. The obtained results were discussed in the context of the EU policies on the use of antibiotics and heavy metals in animal food production. It is worth noting that the province of Asturias in Northern Spain was the location where IncC plasmids of the Spanish monophasic clone were first identified [17].
2. Results
2.1. Incidence and General Properties of Monophasic Isolates of the Spanish Clone During the 2008–2018 Period
A total of 15 out of 615 monophasic isolates (2.4%) recovered from clinical samples in Asturias during 2008 to 2018 were assigned to the Spanish clone and initially selected for the present study (Table 1). The assignment was based on a resistance profile (phenotype and responsible genes) compatible with that typically associated with the Spanish clone or its variants, together with the presence of a plasmid of the IncC incompatibility group. Specifically, fourteen isolates were obtained from the faeces of different patients, and one was also present in the urine of one of those patients. For all fifteen isolates, the antigenic formula was experimentally determined to be 4,5,12:i:-.
2.2. Genome Sequence Analysis
The genomes of the 14 epidemiologically unrelated isolates were sequenced with short-read Illumina technology. Of the two isolates obtained from faeces and urine from the same patient, only the former was sequenced, as preliminary characterization indicated that both belonged to the same strain (Table 1). In addition, 21 clinical isolates from the 2000 to 2003 period and the old LSP 272/98 isolate [17] were also sequenced. The assembly size of the genomes ranged from 4,984,459 to 5,156,906 kb, built from 50 to 79 contigs (Table S1). All isolates could be assigned to ST19 by MLST performed in silico, as expected for the Spanish monophasic clone. Genome sequence analyses also led to the identification of genes related to both antibiotic and biocide resistance (Section 2.3). Moreover, the information gained for antibiotic resistance genes was consistent with the antibiogram results and with the presence of the relevant genes confirmed by PCR amplification, performed herein for the 2008–2018 isolates and previously reported for the older ones [14,17]. New information on the presence of biocide resistance genes and plasmid content was obtained by in silico analyses of all genome sequences.
The results from the plasmid analyses, as revealed by PLACNETw reconstruction, PBRT, PlasmidFinder, and pMLST, are summarized in Table 1. Apart from the IncC plasmid common to all isolates (Section 2.3), two of them (LSP 1142/03 and LSP 2/15) carried an IncI1-I(α) plasmid of ca. 96.4 and 116.2 kb, assigned to ST154 and ST259, respectively. In silico analysis of the genomes also showed the presence of a 54.8 kb plasmid in one isolate (LSP 54/18), whose incompatibility group could not be determined, and of smaller plasmids (ranging in size from 1.9 to 14.5 kb) in all isolates, except LSP 389/97. The latter plasmids were identified as OriColE, Col(BS512), Col8282, and Col(pHAD28) or had an unknown replicon. With regard to the phenotypes conferred by the identified plasmids, all were cryptic, except for the IncI1-I(α) plasmids, which contained the strA, strB, and sul2 genes encoding resistance towards streptomycin and sulphonamides, and the IncC plasmids, which carried all other resistance genes found in the isolates, including antibiotic and biocide resistance genes, as well as virulence genes of pSLT.
2.3. Comparison of IncC Plasmids from the 2000 to 2003 and 2008 to 2018 Periods
The IncC plasmids of all sequenced clinical isolates were comprehensively characterized by means of bioinformatic analyses, and those belonging to older and more recent isolates were compared. The plasmids ranged in size between ca. 126 and 197.4 kb. The pMLST scheme for IncC plasmids includes the conserved repA, parA, parB, and orf053 genes involved in plasmid replication and maintenance [22]. In silico pMLST performed for the IncC plasmids of the Spanish clone identified the repA_4*, parA_5*, parB_6, and A053_2 alleles, yielding an allelic profile different from those recorded for other plasmids of the group. The new ST was termed ST10* by the CGE tool since it was more closely related to ST10, differing by the alleles marked with an asterisk.
As indicated before, IncC plasmids of the older isolates were previously characterized using a wide variety of experimental approaches [14,15,17], but they had not been previously sequenced. Sequence analysis performed herein provided additional information on virulence and resistance genes carried by them and allowed their comparison with those circulating in more recent years. Figure 1 shows the relevant genes in the order they appear in pUO-STmRV1, the plasmid of LSP 389/97, distributed along the four regions of foreign DNA (termed Ins1 to Ins4) inserted within the highly reduced backbone of the control plasmid [21].
With regard to virulence genes, the spv locus (located within Ins1) was found to be intact in all plasmids, except pLSP 272/98 and pLSP 438/15, which lacked the entire locus, and pLSP 61/13, in which three out of the five spv genes were missing. This is in line with the original description of pLSP 272/98 as a resistance plasmid (formerly known as pUO-SR4 and pUO-STmR1), not as a resistance–virulence plasmid, based on the absence of the spv locus [14,17]. Yet, two other virulence genes of pSLT, tlpA and mig5, belonging to Ins2, were detected in all IncC plasmids under study, including pLSP 272/98. In relation to resistance profiles, the pUO-STmRV1-like plasmids of the Spanish monophasic clone displayed a great diversity (Figure 1). This is, for instance, reflected in the highly variable number of antibiotic resistance genes, which ranged between three and twelve, with only sul2 shared by all plasmids. Variability was also observed for genes associated with resistance to heavy metals (mercury, silver, copper, and organo-arsenicals) and quaternary ammonium compounds. Taken together, resistances could be categorized into 22 ABR (antibiotic biocide resistance) gene profiles. A BRIG comparison of 22 plasmids selected as representatives of the different ABR profiles is shown in Figure 2.
Fourteen and ten profiles were associated with plasmids belonging to older and newer isolates, respectively, with only two of them (ABR4 and ABR13) being common to plasmids from both periods. The highest number of genes (33) was carried by the control pUO-STmRV1 and by pLSP 66/01, which shared the same profile (ABR-1) and were the only plasmids containing the bleO gene for bleomycin resistance. In contrast, the profile of pLSP 272/98 (ABR-2), also used as a control, was not shared by any other plasmid. The most frequent profile was ABR-3, which comprised all resistance genes present in pUO-STmRV1, except bleO, and was associated with eight plasmids, all found in isolates from the 2000 to 2003 period. The second most common profile was ABR-16, which was shared by four plasmids of recent isolates. The remaining profiles were associated either with one (fourteen profiles) or two (four profiles) isolates.
Variability concerning the antibiotic resistance mainly affected the class 1 integron of the sul3 type and the defective class 1 integron of the sul1 type with the 1600 bp/dfrA12-aad2 variable region, which lacks the 3′-conserved segment where the sul1 gene is located. In fact, both integrons were absent in all but two plasmids of the second period, as well as in two plasmids of the first period. Smaller deletions, which removed dfrA12 or aadA2, appeared in the integrons of another three plasmids of the first period. Deletions affecting most other antibiotic resistance genes, including tet(A) and blaTEM-1B (supplied by defective Tn1721 and Tn2 transposons, respectively) and sul1 (carried by a second class 1 integron of the sul1 type termed In0, which lacks the variable region), as well as aac(3)-IVa for gentamicin and tobramycin resistance, were also observed, though to a lesser extent. As indicated before, the only antibiotic resistance gene conserved in all plasmids appears to be sul2, which belongs to the GIsul2 integrative element [23]. Similar to the antibiotic resistance loci, deletions that removed all or part of those associated with resistance to quaternary ammonium compounds and heavy metals, including copper, silver, mercury, and organo-arsenical compounds, were also detected.
2.4. Genetic Basis of the Monophasic Phenotype
To precisely determine the genetic basis of the monophasic phenotype, the fljAB chromosomal region of the isolates from the 2008 to 2018 period was compared with the corresponding region of Typhimurium LT2 (Figure 3).
All monophasic isolates had a large deletion, which starts at the same point within STM2758 (according to the annotation of the LT2 genome) and extends into iroB (STM2773), therefore removing the fljA, fljB, and hin genes. In all cases, only 100 bp out of the 1542 bp of the intact STM2758 was retained, while three different deletions, which eliminated 633, 808, and 683 bp out of 1116 bp of iroB, were identified. The observed deletions were designated as ΔfljAB-1, ΔfljAB-3_AS_, and ΔfljAB-4_AS_, following the numbering of previously published deletions for isolates from our region [15]. ΔfljAB-1 was detected in eleven out of the fourteen isolates studied herein (78.6%), while ΔfljAB-3_AS_ and ΔfljAB-4_AS_, which were newly discovered, were represented in only one and two isolates, respectively. The subscript AS (Asturias) was added to distinguish them from deletions with the same numbers previously identified in Spain outside Asturias [20]. ΔfljAB-2, reported in older isolates from our region [15], was not found in the more recent isolates. Yet, the same deletion was detected in other Spanish isolates, together with ΔfljAB-3 and ΔfljAB-4, which are different from ΔfljAB-3AS and ΔfljAB-4AS [20]. Supporting previous findings, all isolates carried a copy of IS26 between the start and end points of the observed deletions [15,20]. Interestingly, PacBio sequencing of the control isolate LSP 389/97 revealed a very large inversion encompassing ca. 2605 kb of chromosomal DNA, placed between truncated rtcB and STM2758 genes. This inversion was not present in the first analysis of this strain [15] and must have occurred at a later time during the cultivation in the laboratory (further elaborated on in Section 3).
2.5. Genomic Relationships Between the Isolates
To establish the genomic relationships between clinical isolates belonging to the Spanish clone, an SNP (single nucleotide polymorphism)-based phylogenetic tree was constructed, using 35 clinical isolates, 21 from the 2000 to 2003 period and 14 from the 2008 to 2018 period, as well as the control isolates LSP 389/97 and LSP 272/98. Two other isolates belonging to the Spanish monophasic clone and recovered from food samples in our region (LSP 87/13 and LSP 195/13) were also included in the tree. As shown in Figure 4, the 39 isolates were closely related, differing by a minimum of one (LSP 48/12 versus LSP 62/12) and a maximum of 103 (LSP 54/18 versus LSP 148/18) SNPs (Supplementary Table S2). Nevertheless, they formed two clades, termed A and B. Clade A comprised 27 isolates, varying by four to eighty-five SNPs, while the remaining twelve isolates clustered within clade B with one to fifty-seven SNPs. With two exceptions, clade A grouped isolates of the first period, including the “old” control strains LSP 389/97 and LSP 272/98, while clade B only contained isolates from the second period. To facilitate the categorization, Figure 4 also includes the phage type, the type of fljAB deletion, and the ABR profile and the spv genes contributed by the IncC plasmids.
Overall, 59.5% of the clinical isolates tested for phage type belonged to U302 (22/37, including LSP 389/97 and LSP 272/98). Others, like DT193 (8), U210 (1), and NT (4), which could not be typed with the phage library employed, were also detected (note that phage typing in Spain was discontinued in 2017). With regard to the fljAB deletions, ΔfljAB-1 was the most common, shown by 86.5% of the clinical isolates (32/37), while ΔfljAB-3_AS_ and ΔfljAB-4_AS_ were only shown by a limited number of isolates from the second period (Section 2.4), and ΔfljAB-2 was exclusively found in isolates from the first period [15]. Interestingly, the resistance profiles of the two isolates from the second period, which grouped in clade A (LSP 207/08 and LSP 148/18), comprised the highest number of antibiotic resistance genes (ABR-13 and ABR-22), with ten and eleven, respectively, versus three to five in the remaining profiles of isolates from the same period grouped into clade B.
3. Discussion
The present study focused on the incidence, variability, and evolution of the Spanish monophasic clone of S. Typhimurium. For this, all isolates of the clone recovered from human clinical samples between 2008 and 2018 in the northern Spanish region of Asturias were thoroughly characterized by conventional techniques combined with genome sequence analyses and compared with older isolates from the same region. During the 2008–2018 period, the Spanish clone accounted for 2.4% of the total monophasic isolates of clinical origin detected in Asturias. Other isolates were mostly assigned to the European ST34 clone, which is nowadays one of the most common foodborne pathogens involved in human infections worldwide [6]. In contrast, the Spanish clone belongs to ST19 and has a more restricted geographical distribution, being mainly detected in the Iberian Peninsula [10,11,12,13,14,15,16,17]. Moreover, its frequency has declined over time, from being the fourth most common serovar in 1998 in Spain [12]. The rise and decline of multidrug-resistant clones, which occur in successive waves, have determined the epidemiology of S. Typhimurium and its monophasic variant in Europe over the past decades [25,26,27]. Though the forces driving clone succession remain largely elusive, the early success of the Spanish clone and its subsequent replacement by the European clone in Spain have been proposed to be associated with changes in the European Union regulations for the use of antibiotics and heavy metals in food animal production [28]. In the present study, isolates from the second period were recovered soon after the implementation of the ban on the use of antibiotics as growth promoters for food animals, which became compulsory in 2006 [29]. This could explain the observed decrease in the number of antibiotic resistance genes, which dropped from an average of 9.6 during the first period, when the use of antibiotics as feed additives was still permitted, to an average of 5.5 during the second period. Compliance in Asturias with the EU regulations and Spanish legislation concerning the prudent use of antibiotics to combat resistance, both in veterinary and human medicine, has probably contributed to the observed reduction. Similarly, the widespread application of heavy metal compounds in food–animal production may have contributed to the selection and expansion of S. 4,[5],12:i:- isolates [28,30,31,32]. This could apply to the Spanish and the European clone since both contain genes conferring resistance to copper, silver, mercury, and arsenic compounds. However, while the MICs to mercury and arsenic were similar, MICs to silver and copper were much lower in the Spanish than in the European clone [28]. Mercury and arsenic compounds are increasingly regarded as undesirable contaminants in animal feed, with attempts to reduce their concentration and hence diminish the selective pressure for resistance mechanisms [33]. However, this does not concern copper and silver derivatives, which are still employed in food animal production. The increased resistance to these compounds due to the acquisition of SGI-4 by the European clone could explain the reduced appearance of the Spanish clone and its replacement by the European clone, which in Spain was estimated to occur around 2008 [28].
In the context of changing regulations on antibiotic and heavy metal usage in the EU, the structure of the IncC plasmids of the Spanish monophasic clone has a direct impact on the evolution of resistance. Thus, the presence of multiple copies of IS26 (14 in pUO-STmRV1), together with many other potentially mobile genetic elements and repeated DNA [21], cause an intrinsic instability of these plasmids. This is consistent with the high number of variants identified herein and reported previously [14,16,17]. Interestingly, earlier attempts to cure the entire IncC plasmids from the Spanish clone failed but led to the identification of smaller variants originating from the loss of genes conferring resistance to ampicillin, chloramphenicol, gentamicin, streptomycin, tetracycline, and/or trimethoprim [16,17]. As shown in the present study, multiple variants lacking one or more of these and other resistance genes were detected in naturally occurring isolates infecting humans in our region. Accordingly, the IncC plasmids of the Spanish monophasic clone appear to combine a highly stable backbone (though largely degenerated [21]) with remarkably unstable accessory DNA. Together with efficient replication and partition mechanisms, vertical transmission of the plasmids is guaranteed by the toxin–antitoxin system encoded by the ant/tox (also known as ata/tad) genes within the IncC backbone, reinforced by the ccdAB genes of pSLT located within Ins1 [21]. As these plasmids are incapable of conjugative transfer due to extensive deletions within the tra1 and tra2 regions involved in conjugation, horizontal spread into non-clonal isolates does not occur [14,21]. Moreover, the accessory DNA is well-suited to respond to changes in selective pressure. In fact, the imprint of IS26 on the evolution of pUO-STmRV1 was revealed by sequence analysis of the whole plasmid genome [21], and this insertion sequence may well be playing a major role in the generation of variants.
Apart from enabling readjustments in antimicrobial drug resistance, IS26 is also responsible for the monophasic phenotype of the Spanish clone. Four different deletions were identified in our region (∆fljAB-1, ∆fljAB-2, ∆fljAB-3_AS_, and ∆fljAB-4_AS_), all removing the fljAB operon and its flanking DNA. The start point of these deletions within ST2758, common to all of them, and the variable end point located within the iroB gene, are separated by a single copy of IS26. This observation can be explained by two consecutive transposition events [34,35]. The first one would have been an intermolecular transposition of IS26 into the chromosomal STM2758 gene of S. Typhimurium (marked with “1” in Figure 3). A pUO-STmRV1-like plasmid, already acquired by a biphasic isolate likely of phage type U302, probably acted as donor in this event. Although pU302L, found in S. Typhimurium U302 and carrying four copies of IS26, was suggested as the possible donor of the element [20], to the best of our knowledge this plasmid has not been detected in the Spanish clone. The second event would have been an intramolecular transposition of IS26 from ST2758 into iroB (“2” in Figure 3) using the cis-pathway. This would remove the intervening DNA leading to the monophasic phenotype, and it must have occurred independently more than once to account for the different deletions observed. A similar event would explain deletions extending beyond iroB, which were reported in Spain by other authors [18,20]. Of note, after acquiring the ΔfljAB-1 deletion, IS26 transposition into the rtcB gene (“3” in Figure 3) of the control LSP 389/97 isolate using the trans-pathway led to the inversion of a large DNA segment, spanning between ∆STM2758 and ΔrtcB. This view is substantiated by the fact that TSDs (target site duplications) of 8 bp, originally present in rtcB, were observed, although the position and orientation of one of the repeats changed upon inversion (5′-GCATGGCG-3′ and 5′-CGCCATGC-3′). Since this inversion did not yet occur when LSP 389/98 was first tested for the fljAB deletion [15], this is a good example of the “hyperactivity” of IS26, which once incorporated into a new DNA molecule will continue to act as an efficient motor for further evolution.
Apart from explaining the monophasic phenotype, the proposed scenario is consistent with the close relationship existing between the monophasic isolates of the Spanish clone, as previously revealed by traditional typing techniques, like phage typing, PFGE, MLVA, and microarray analysis [14,15] and corroborated by SNP-based phylogenetic analysis performed herein. In fact, all isolates of the Spanish clone may derive from a single biphasic ancestor of S. Typhimurium U302 that either acquired pUO-STmRV1 or in which the resistance–virulence plasmid was first assembled. After transposition of IS26 from the plasmid into STM2758, intra-clonal evolution would not only explain the various deletions leading to the monophasic phenotype but also the differences in phage type, resistance profiles, and the limited number of SNPs detected within the genomes.
4. Materials and Methods
4.1. Isolate Selection and Preliminary Characterization
All S. 4,[5],12:i:- isolates recovered from human clinical samples in Asturias (a Northern Spanish region) during the 2008–2018 period (615) were experimentally tested for antibiotic resistance properties, resistance genes, and plasmid content. Fifteen of them, displaying properties consistent with the Spanish clone, were selected for the present study (Table 1). They were obtained from the faeces of different patients with gastroenteritis at four hospitals in the region, and also from the urine of one of the patients. For all of them, the serotype was experimentally determined, and for those recovered before 2017, the phage type was also established. The information was provided by the Laboratory of Public Health (LSP) of Asturias. Antibiotic susceptibility testing and detection of the resistance genes were experimentally performed as in [36]. Screening for the IncC and IncN replicons was carried out by the PBRT (PCR-based replicon typing) method [37]. LSP 389/97 (the isolate carrying pUO-STmRV1), LSP 272/98, which are two of the oldest monophasic isolates of the Spanish clone recovered in our region [17], and S. Typhimurium LT2, the type strain of the genus Salmonella [24], were used as controls in different experiments.
4.2. Whole Genome Sequencing and Bioinformatics Analysis
Fourteen epidemiologically unrelated isolates belonging to the Spanish clone and recovered from clinical samples during the 2008–2018 period were sequenced using short-read Illumina technology, as described [36]. Another 21 isolates of the same clone, obtained between 2000 and 2003 and characterized in previous studies [14,15], as well as LSP 272/98, were additionally sequenced. The obtained raw reads were assembled into contigs with the VelvetOptimiser.pl script implemented in the online version of PLACNETw (https://castillo.dicom.unican.es/upload/; last accessed on 12 April 2021). The assembled genomes were annotated by the NCBI Prokaryotic Genome Annotation Pipeline (PGAP; https://www.ncbi.nlm.nih.gov/genome/annotation_prok/; last accessed on 28 June 2021) and deposited in GenBank under the accession numbers provided in Table S1. Bioinformatic analyses were performed both with PLACNETw and several tools available at the Center for Genomic Epidemiology (CGE; https://www.genomicepidemiology.org/services/; last accessed on 12 June 2025) of the Technical University of Denmark (DTU), including SeqSero, MLST, ResFinder, PlasmidFinder, and pMLST. Heavy metal resistance genes were identified by MEGares 2.0 and with a customized database containing heavy metal resistance genes previously detected in S. enterica, which was screened with MyDbFinder (CGE, DTU). Analysis of fljAB regions was performed with the aid of BLASTn and CLONE Manager (ClonSuit9), and also with a second in-house customized database comprising all open reading frames located between STM2752 and the iroC gene in the chromosome of biphasic S. Typhimurium LT2 (accession number AE006468.1). The entire sequence of LSP 389/97, including the chromosome (accession no. CP018219) and plasmid pUO-STmRV1 (accession no. CP018220), has been previously generated by combining short-read Illumina and long-read PacBio technologies [21]. Both sequences were used as controls for the in silico analyses. Graphic representations derived from IncC plasmids were generated with BRIG (Blast Ring Image Generator; https://sourceforge.net/projects/brig/; last accessed on 10 July 2025), using pUO-STmRV1 as the reference.
4.3. Phylogenetic Analysis
The phylogenetic relationships between isolates of the Spanish monophasic clone sequenced in the present study (a total of 36) were inferred with the CSI tool (version 1.4) available at the CGE website (last accessed on 17 September 2021). LSP 389/97 and two isolates of the same clone recovered from food samples in our region (LSP 87/13 and LSP 195/13) were also included in the tree. The pipeline was run with default parameters, using the genome of LSP 389/97 as reference for SNP calling, and the resulting SNP matrix is shown in Table S2. Bootstrap support for the consensus phylogenetic tree was based on 1000 replicates [38].
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Majowicz S.E. Musto J. Scallan E. Angulo F.J. Kirk M. O’Brien S.J. Jones T.F. Fazil A. Hoekstra R.M. International Collaboration on Enteric Disease ‘Burden of Illness Studies’ The global burden of nontyphoidal Salmonella gastroenteritis Clin. Infect. Dis.20105088288910.1086/65073320158401 · doi ↗ · pubmed ↗
- 2Issenhuth-Jeanjean S. Roggentin P. Mikoleit M. Guibourdenche M. de Pinna E. Nair S. Fields P.I. Weill F.X. Supplement 2008-2010 (no. 48) to the White-Kauffmann-Le Minor scheme Res. Microbiol.201416552653010.1016/j.resmic.2014.07.00425049166 · doi ↗ · pubmed ↗
- 3ECDC (European Food Safety Authority and European Centre for Disease, Prevention and Control) The European Union One Health 2022 Zoonoses Report EFSA J.202321 e 84423808947110.2903/j.efsa.2023.8442 PMC 10714251 · doi ↗ · pubmed ↗
- 4EFSA (European Food Safety Authority) Panel on Biological Hazards (BIOHAZ) Scientific Opinion on monitoring and assessment of the public health risk of “Salmonella typhimurium-like” strains EFSA J.20108182610.2903/j.efsa.2010.1826 · doi ↗
- 5ECDC (European Food Safety Authority and European Centre for Disease, Prevention and Control) The European Union One Health 2021 Zoonoses Report EFSA J.202220 e 0766610.2903/j.efsa.2022.766636524203 PMC 9745727 · doi ↗ · pubmed ↗
- 6Sun H. Wan Y. Du P. Bai L. The epidemiology of monophasic Salmonella Typhimurium Foodborne Pathog. Dis.202017879710.1089/fpd.2019.267631532231 · doi ↗ · pubmed ↗
- 7Switt A.I. Soyer Y. Warnick L.D. Wiedmann M. Emergence, distribution, and molecular and phenotypic characteristics of Salmonella enterica serotype 4,5,12:i Foodborne Pathog. Dis.2009640741510.1089/fpd.2008.021319292687 PMC 3186709 · doi ↗ · pubmed ↗
- 8Silverman M. Simon M. Phase variation: Genetic analysis of switching mutants Cell 19801984585410.1016/0092-8674(80)90075-66247071 · doi ↗ · pubmed ↗