First Report of Stenotrophomonas maltophilia from Canine Dermatological Infections: Unravelling Its Antimicrobial Resistance, Biofilm Formation, and Virulence Traits
Ria Rajeev, Porteen Kannan, Sureshkannan Sundaram, Sandhya Bhavani Mohan, Sivachandiran Radjendirane, Chaudhary Jeetendrakumar Harnathbhai, Anbazhagan Subbaiyan, Viswanathan Naveenkumar, Nithya Quintoil Mohanadasse, Wilfred Ruban Savariraj, Charley A. Cull

TL;DR
This study reports the first occurrence of S. maltophilia in dogs with skin infections, highlighting its antibiotic resistance, biofilm formation, and potential to spread to humans.
Contribution
First documentation of S. maltophilia in canine dermatological infections with detailed analysis of its resistance and virulence traits.
Findings
15 S. maltophilia isolates showed resistance to cefpodoxime and aztreonam but susceptibility to several other antibiotics.
High efflux activity and biofilm formation were observed in most isolates, indicating strong virulence potential.
Resistance and virulence genes were positively correlated, suggesting co-evolution of these traits.
Abstract
Background/Objectives: The present study was aimed at documenting S. maltophilia occurrence in dogs with skin ailments, investigating its virulence, biofilm-forming ability, antimicrobial susceptibility, and zoonotic potential to inform preventive and therapeutic strategies against multidrug resistant S. maltophilia infections. Methods: Skin swabs (n = 300) were collected from dogs with dermatological ailments. Isolation was performed using selective media and confirmed with molecular methods, validated by MALDI Biotyper. Antimicrobial susceptibility testing and efflux activity assessment were conducted. Resistance genes related to sulfonamides, quinolones, and β-lactams were screened. Virulence was assessed by biofilm formation, motility, and virulence gene profiling. Results: In total, 15 S. maltophilia (5%) isolates were identified. All 15 isolates were susceptible to…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
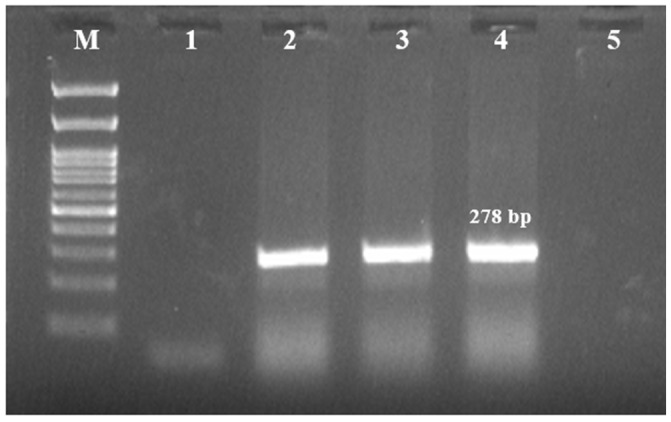 Figure 1
Figure 1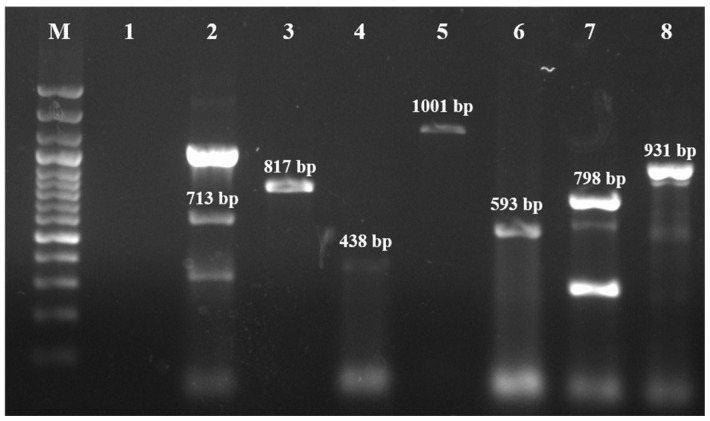 Figure 2
Figure 2 Figure 3
Figure 3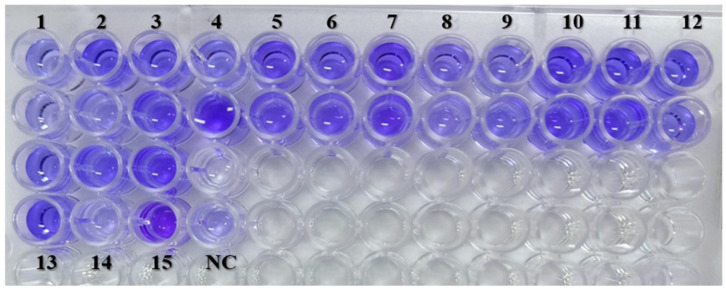 Figure 4
Figure 4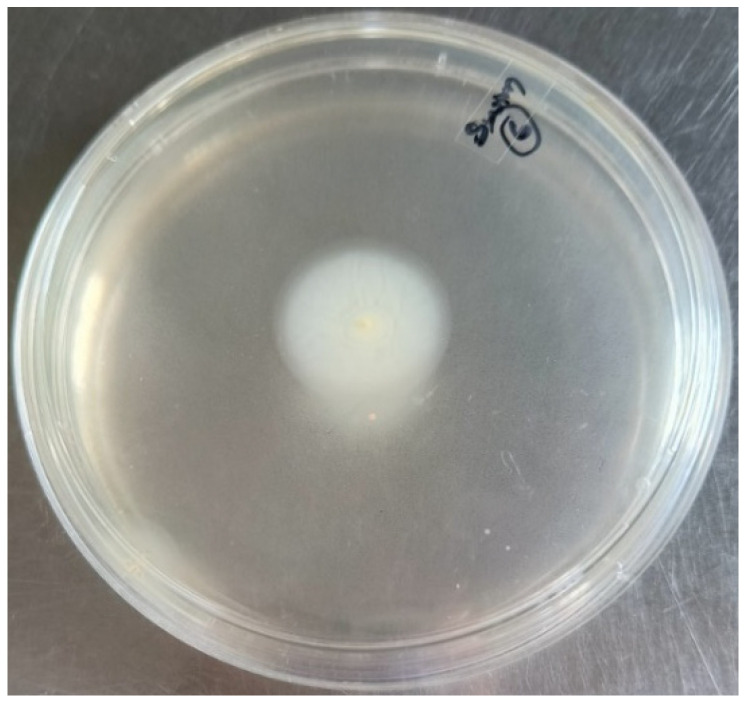 Figure 5
Figure 5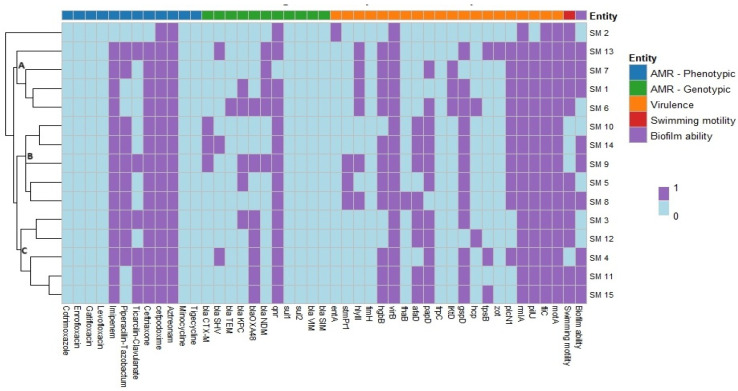 Figure 6
Figure 6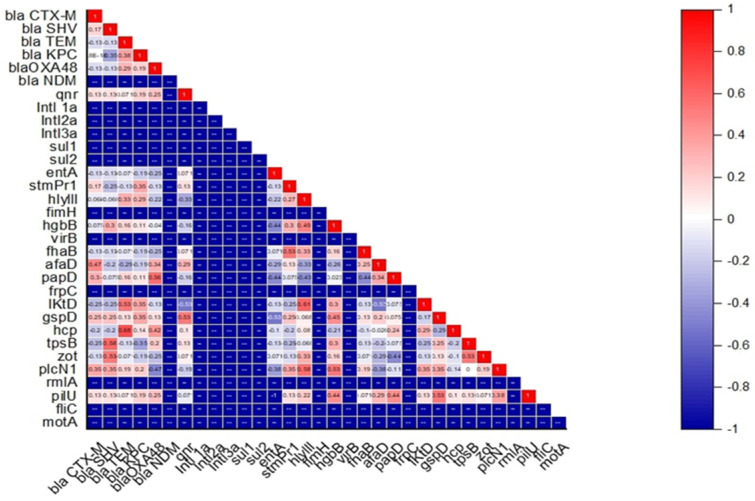 Figure 7
Figure 7Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInfections and bacterial resistance · Bacterial biofilms and quorum sensing · Antimicrobial Resistance in Staphylococcus
1. Introduction
The genus Stenotrophomonas, belonging to the class Gammaproteobacteria, consists of non-fermenting gram-negative bacilli (NFGNB) that occur ubiquitously in environments such as soil, water, plant rhizospheres, and the bodies of animals and humans [1]. Stenotrophomonas maltophilia is the primary pathogen within this genus, implicated in various infections in humans. This opportunistic pathogen causes nosocomial infections, including respiratory, wound, urinary tract, ocular, and soft tissue infections, and ranks as the third most frequently isolated unusual NFGNB after Pseudomonas aeruginosa and Acinetobacter baumannii [2].
S. maltophilia expresses numerous virulence factors that facilitate host colonization, such as adhesins (lipopolysaccharides, flagella, type 1 fimbriae, type IV pili) and extracellular enzymes (proteases, esterases, lipases, gelatinase, hemolysin, siderophores, and cytotoxins) [3]. Its biofilm-forming ability on abiotic surfaces and host tissues diminishes the efficacy of drugs used to treat hospital-acquired infections [3]. Additionally, quorum sensing via autoinducers regulates gene expression based on cell density [4].
S. maltophilia is intrinsically resistant to a wide range of antibiotics, including β-lactams, fluoroquinolones, tetracyclines, chloramphenicol, and aminoglycosides. This resistance is mediated by two β-lactamases, L1 (a metallo-β-lactamase) and L2 (a clavulanic acid-susceptible cephalosporinase) [1]. Aminoglycoside resistance is also supported by modifying enzymes [AAC (6′)-Iz, APH (3′)-Iic, AAC (6′)-Iak] and efflux pumps (SmeABC, SmeIJK, SmeYZ) [1]. Additionally, resistance genes such as sul1, sul2, and dfrA associated with Class 1 integrons confer resistance to trimethoprim/sulfamethoxazole (TMP/SMX). TMP/SMX is the drug of choice for treating S. maltophilia infections, with alternate options like levofloxacin, minocycline, tigecycline, and ceftazidime used for TMP/SMX-resistant cases [2].
S. maltophilia has been isolated from animals with chronic respiratory disease and urinary infections, including horses, cats, dogs, and ball pythons [5,6]. However, only one study in India has reported its occurrence in animals, specifically in sarcoptic pig skin ulcers [7], with no prior studies in dogs. This study aimed to document S. maltophilia occurrence in dogs with skin conditions, evaluating its virulence, biofilm-forming capacity, and antimicrobial susceptibility to assess zoonotic potential and develop treatment strategies for biofilm-associated multidrug resistant S. maltophilia infections.
2. Results
2.1. Prevalence of S. maltophilia
A total of 15 isolates (5%, 15/300) showed characteristic colonies of S. maltophilia and were species-identified by morphological, biochemical, molecular (Figure 1), and MALDI-TOF MS methods (Table 1).
2.2. Characterization of Antimicrobial Resistance in S. maltophilia
All S. maltophilia isolates were susceptible to six antimicrobials: cotrimoxazole, enrofloxacin, gatifloxacin, levofloxacin, minocycline, and tigecycline. However, all isolates were resistant to cefpodoxime and aztreonam. Additionally, 14 isolates (93.33%) were resistant to imipenem and ceftriaxone, while 10 isolates (66.67%) exhibited resistance to piperacillin-tazobactum (Table 2). Molecular characterization revealed the presence of various ARGs in the isolates (Figure 2), with the qnr gene being most prevalent (93.33%). Other detected ARGs included blaOXA-48 (n = 7, 46.67%), blaKPC (n = 5, 33.33%), blaNDM (n = 5, 33.33%), blaCTX-M (n = 3, 20%), blaSHV (n = 3, 20%), and blaTEM (n = 1, 6.67%) (Table 2). In addition to this, evaluation of the S. maltophilia isolates for bacterial pump efflux activity by the Etbr agar cartwheel method showed no fluorescence on the agar plates indicating active efflux activity (Figure 3) (Table 2).
2.3. Virulence Characterization of S. maltophilia
The crystal violet staining assay revealed that 9 of the S. maltophilia isolates (60%) were strong biofilm producers, while 6 isolates (40%) were moderate biofilm producers (Figure 4) (Table 2). Biofilm strength was categorized based on OD values as Negative: < 0.044; Weak: 0.044 < A ≤ 0.088; Moderate: 0.088 < A ≤ 0.176 and Strong: A > 0.176, where A-OD Value of Sample.
Virulence gene analysis revealed all the isolates harbored virB (100 %), motA (100 %), rmlA (100 %), and fliC (100 %) genes followed by pilU (93.3 %), gspD (80 %), papD (73.3 %), hgbB (73.3 %), picNl (66.6 %), afaD (53.3 %), and hlyIII (40 %) genes, respectively. However, the frpC and fimH genes were not found in any of the isolates (Table 2).
Swimming motility was assessed on modified LB plates, revealing that all 15 isolates exhibited swimming motility. Based on the diameter of the motility zone, 11 isolates (73.3%) were classified as strong, while 4 isolates (26.67%) were classified as moderate (Figure 5).
2.4. Association Between Antimicrobial Resistance, Virulence, Motility Pattern, and Biofilm-Forming Ability in S. maltophilia
The link between antimicrobial resistance, virulence, motility pattern, and biofilm-forming ability of S. maltophilia isolated from skin swabs of dogs with dermatological ailments was established by hierarchical clustering (dendrogram). The heatmap showed the grouping of isolates into mainly three clusters (A, B, and C) (Figure 6). Cluster A consisted of 4 isolates namely SM 13, SM 7, SM 1, and SM 6 which harbored virB, motA, rmlA, fliC, pilU, hgbB, picNl, and hlyIII virulent genes; blaNDM ARG and resistant to cefpodoxime, aztreonam, imipenem, and ceftriaxone in common. Cluster B consisted of 5 isolates namely SM 10, SM 14, SM 9, SM 5, and SM 8 which harbored virB, motA, rmlA, fliC, pilU, picNl, and gspD virulent genes; qnr ARG and resistant to cefpodoxime, aztreonam, imipenem, piperacillin-tazobactum, and ceftriaxone in common. Cluster C consisted of 5 isolates namely SM 3, SM 12, SM 4, SM 11, and SM 15 which harbored virB, motA, rmlA, fliC, pilU, and papD virulent genes; qnr and blaO ARGs and resistant to cefpodoxime, aztreonam, imipenem, and ceftriaxone in common. Isolate SM 2 remained outgroup. Interestingly, among the 15 isolates, none of the isolates harbored the same pattern of antimicrobial resistance, virulence, motility pattern, and biofilm-forming ability. Further association between antimicrobial resistance and virulence gene was established. A moderate positive correlation was observed between blaSHV with zot and tpsB (+0.53 to +0.58), blaTEM with lktD and hcp (+0.53 to +0.68), blaOXA-48 with papD (+0.56) and qnr with gspD (+0.53) (Figure 7). On the other hand, a weak negative correlation was seen between blaKPC with tpsB (−0.35), blaOXA-48 with plcN1 (−0.47), and qnr with hylIII and lktD (−0.33 to −0.53) (Figure 7).
3. Discussion
S. maltophilia has emerged as an important opportunistic pathogen which has been increasingly reported worldwide [1]. Although traditionally considered as a human pathogen, it has been reported in dogs, horses, cats, dogs, and ball pythons [5,6]. Earlier studies have reported that animal strains of S. maltophilia shared phylogenetic traits with some of the most successful human strains [8]. In this study, 15 S. maltophilia isolates were obtained from the skin swabs of dogs with dermatological ailments. These results indicate its involvement either as a primary pathogen or as part of a polymicrobial infection. Further studies would be necessary to confirm whether S. maltophilia is a direct cause of the dermatological problems or a secondary invader exploiting the diseased environment. This is the first study globally to document S. maltophilia from dogs with dermatological ailments.
An antibiotic susceptibility test revealed that the isolates were susceptible to cotrimoxazole, enrofloxacin, gatifloxacin, levofloxacin, minocycline, tigecycline, and ticarcillin-clavulanate. As fluoroquinolones and tetracyclines are commonly used in both human and veterinary medicine, the findings of this study support their potential use for treatment in cases involving S. maltophilia [2]. Furthermore, the isolates were found to be resistant to cefpodoxime, aztreonam, imipenem, ceftriaxone, and piperacillin-tazobactam. Carbapenem resistance is particularly worrisome, as they are considered a last-resort antibiotic for treating MDR infections [9]. A limitation of this study is the lack of specific CLSI breakpoints for S. maltophilia for several antibiotics, including enrofloxacin, gatifloxacin, imipenem, piperacillin-tazobactam, ceftriaxone, cefpodoxime, aztreonam, and tigecycline. In the absence of established breakpoints for S. maltophilia, we interpreted antimicrobial susceptibility results based on the available literature and expert recommendations, which may not fully reflect clinical susceptibility patterns. Further studies are needed to refine and standardize breakpoints for S. maltophilia to improve the accuracy and relevance of antimicrobial resistance data for this pathogen.
The resistance to β-lactams and carbapenems might be attributed to intrinsic resistance mechanisms typical of S. maltophilia or acquired resistance genes [10]. The molecular characterization of ARGs in S. maltophilia isolates demonstrates a high prevalence of the qnr gene (93.33 %) and a concerning presence of multiple β-lactamase genes, including blaOXA-48 (46.67 %), blaKPC (33.33%), blaNDM (33.33%), blaCTX-M (20%), blaSHV (20%), and blaTEM (6.67%). There was a broad spectrum of β-lactamase enzymes, capable of hydrolyzing a wide range of β-lactam antibiotics, including carbapenems, cephalosporins, and penicillins, which are often considered last-resort treatments for MDR infections. While this represents the first investigation of S. maltophilia specifically from canine dermatological sources, our findings can be meaningfully contextualized within the broader literature. Our detection of qnr (93.3%) aligns with previous studies reporting a high prevalence of quinolone resistance mechanisms in S. maltophilia clinical isolates [11,12]. The presence of multiple β-lactamase genes including blaOXA-48 (46.7%), blaKPC (33.3%), and blaNDM (33.3%) is consistent with emerging trends of carbapenemase-producing S. maltophilia in healthcare settings [13,14,15], though the co-occurrence of multiple resistance genes in our canine isolates suggests potential horizontal gene transfer mechanisms warranting further investigation.
Over-expression of the bacterial efflux pump systems is one of the mechanisms by which bacteria exhibit multi-drug resistance [16]. All the isolates showed high efflux activity resulting in MDR phenotype. These findings are in line with the AST results and presence of ARGs which showed that the 15 isolates were resistant to more than 3 antibiotics tested and 14 out of 15 isolates harbored multiple ARGs. These findings point to a complex resistance profile that complicates treatment options for infections caused by these isolates. Therefore, targeted therapeutic strategies are crucial for understanding the genetic basis of resistance and managing the spread of resistant bacteria in both animal and human populations.
Among S. maltophilia virulence factors, biofilm plays an important role in the survivability and virulence. In this study, all S. maltophilia isolates were able to produce biofilm and most of them were strong biofilm producers (60%) followed by moderate biofilm producers (40%) which is comparable to rates reported in clinical S. maltophilia isolates from human infections, where biofilm formation rates typically range from 70 to 90% [17,18,19]. The strong biofilm-forming isolates identified in this study are likely to exhibit increased resistance to standard treatments, as biofilms create a physical barrier that limits the penetration of antibiotics and reduces their efficacy. The slightly lower rates in our study may reflect source-specific variations or environmental factors affecting canine skin isolates. In the study of motility, most of the isolates exhibited strong or moderate levels of swimming patterns; however, none of the isolates were able to swarm which was in line with previous findings [20,21].
The screening of virulence genes in S. maltophilia showed a high prevalence of virB, motA, rmlA, and fliC in 100% of the isolates. Gene virB encodes components of the Type IV secretion system (T4SS), which is involved in the translocation of effector proteins into host cells, contributing to host cell manipulation and immune evasion [22]. Furthermore, motA and fliC are associated with motility, colonization, and biofilm formation, making them essential for bacterial persistence in diverse environments [3]. Also, rmlA is responsible for biosynthesis of rhamnose, a sugar required for the production of lipopolysaccharides (LPS) in the bacterial outer membrane, which plays a role in immune evasion and biofilm formation [23]. Moderate prevalence of pilU (93.3%) encoding for type IV pili, which is essential for bacterial adherence to host tissues and surfaces and translocation; gspD (80%) encoding a component of the Type II secretion system (T2SS); papD (73.3%) involved in the assembly of P pili associated with bacterial adherence; hgbB (73.3%) encoding for a protein involved in iron acquisition, which is crucial in iron-limited environments; and plcN1 (66.6%) responsible for bacterial immune evasion and tissue invasion, further emphasizing the bacterium’s ability to establish infections in various host environments were recorded [3,22,23]. The lower prevalence of virulence genes such as afaD (53.3%) and hlyIII (40%) encoding for adhesins and haemolysin protein were observed among the isolates.
The presence of multiple virulence genes, particularly those related to secretion systems (T4SS and T2SS), motility, adhesion, and iron acquisition, paints a picture of S. maltophilia as a highly adaptable pathogen capable of surviving in diverse environments and host tissues. These virulence factors likely contribute to the bacterium’s ability to persist in chronic infections, form biofilms, and resist host immune responses. The high prevalence of motA and fliC suggests that motility is a critical trait for environmental colonization and infection initiation, while the presence of secretion systems (virB and gspD) underscores the bacterium’s capacity to secrete effector molecules that manipulate host cells and degrade tissues [22].
A heat map was established to study the link between antimicrobial resistance, virulence, motility pattern, and biofilm-forming ability of S. maltophilia which showed that the isolates have a clonal connection. The hierarchical clustering exhibited diverse profiles with no two isolates sharing the same pattern of traits. This could be attributable to the adaptive versatility of S. maltophilia, which enables it to thrive in different environmental niches and cause various infections. These clusters suggests that the pathogenic potential of S. maltophilia may vary significantly between isolates, even when they are from same source.
The correlation matrix analysis revealed interesting associations between ARGs and virulence factors, suggesting potential co-regulation or functional linkages between these traits. The moderate positive correlation between blaSHV, a β-lactamase gene conferring resistance to β-lactam antibiotics, and the virulence genes tpsB (Type VI secretion system) and zot (zonula occludens toxin) (+0.53 to +0.58), suggests that some resistance mechanisms may be linked with the bacterium’s ability to deliver virulence factors to host cells. The association between blaTEM and lktD (leukotoxin D) and hcp (Type VI secretion system protein) (+0.53 to +0.68) points to a possible synergy between resistance to β-lactams and the secretion of toxins or immune-modulating proteins. Similarly, blaOXA-48 a carbapenemase gene, correlates positively with papD (+0.56), a gene involved in the assembly of pili, which is important for bacterial adhesion to host surfaces suggesting that increased resistance to carbapenems may enhance the bacterium’s capacity to adhere to host tissues, potentially making infections harder to treat. The positive correlation between the quinolone resistance gene qnr and gspD (Type II secretion system) suggests that resistance to fluoroquinolones may be linked with the bacterium’s ability to secrete enzymes and toxins, further complicating treatment. A weak negative correlation was seen between blaKPC with tpsB (−0.35), blaOXA-48 with plcN1 (−0.47) and qnr with lktD and hylIII (−0.33 to −0.53) suggesting that resistance of said genes are less dependent on certain virulence mechanisms. The correlation between antimicrobial resistance and virulence genes observed in our study echoes findings from human clinical isolates, where similar associations between resistance mechanisms and pathogenicity factors have been documented [24,25]. This suggests that the pathogenic potential and resistance profiles of S. maltophilia may be conserved across different host species, supporting the zoonotic concern we highlight.
4. Materials and Methods
4.1. Isolation and Characterization of S. maltophilia
4.1.1. Sample Collection
The present study was carried out from January 2024 to March 2024 in the Teaching Veterinary Clinical Complex, Madras Veterinary College, Chennai. A total of 300 skin swabs were collected aseptically from dogs presented with skin ailments and transported to the Bacteriology Laboratory of the Department of Veterinary Public Health and Epidemiology for further processing.
4.1.2. Bacterial Isolation, Identification, and Molecular Confirmation
Skin swabs were inoculated in 5 mL Luria Bertani Broth and incubated at 37 °C for 24 h. The enriched inoculum was then streaked onto Stenotrophomonas selective agar with vancomycin (5 mg), imipenem (32 mg), and amphotericin B (2.5 mg) and incubated overnight at 37 °C. Characteristic S. maltophilia colonies appeared as dark green with a blue halo. Presumptive colonies underwent gram staining, oxidase, catalase, and glucose fermentation tests for confirmation. PCR targeting species-specific 23S rRNA [26] was used for molecular characterization, following a 25 µL reaction protocol with a specific thermal cycle (Table 3). Optimization of PCR was carried out using reference strain S. maltophilia (MCC2083) obtained from the National Centre for Microbial Resource, Pune, India.
4.1.3. Matrix-Assisted Laser Desorption Ionization–Time of Flight Mass Spectrometry (MALDI-TOF MS) Identification
Bacterial isolates were prepared for MALDI-TOF MS following the ethanol-formic acid extraction method according to the manufacturer’s instructions. A loopful of bacteria was suspended in 300 µL distilled water with 900 µL ethanol, centrifuged at 17,000× g for 2 min, and the supernatant discarded. After repeated centrifugation and ethanol removal, the pellet was air-dried and resuspended in 5–50 µL of formic acid–water (70:30). An equal volume of acetonitrile was added, centrifuged, and 1 µL of the supernatant was placed on the MALDI target plate. After drying, it was overlaid with 1 µL matrix solution (α-Cyano-4-hydroxycinnamic acid). Mass spectra were analyzed on a Microflex LT mass spectrometer (Bruker) with Biotyper software (V 3.0) using Bruker bacterial test standard for identification. antibiotics-14-00639-t003_Table 3Table 3List of primers used for the detection of S. maltophilia.GenePrimer SequenceCyclic ConditionNo. of CyclesAmplicon Size (bp)ReferenceStepTemperature and TimeSpecies specific PCR****23S rRNAF-GCTGGATTGGTTCTAGGAAAACGCR-ACGCAGTCACTCCTTGCGInitial denaturation94 °C–5 min 278[26]Denaturation94 °C–45 s30Annealing68 °C–45 sExtension72 °C–45 sFinal extension72 °C–10 min Antimicrobial resistance genesbla_SIM_F-GTACAAGGGATTCGGCATCGR-TGGCCTGTTCCCATGTGAGInitial denaturation 95 °C–4 min 569[27]Denaturation95 °C–45 s30Annealing58 °C–60 sbla_VIM_F-GTTTGGTCGCATATCGCAACR-AATGCGCAGCACCAGGATAG382Extension72 °C–40 sFinal Extension72 °C–5 min blaTEMF-TCCGCTCATGAGACAATAACCR-TTGGTCTGACAGTTACCAATGCInitial denaturation 94 °C–5 min 931[28]Denaturation 94 °C–30 s35Annealing60 °C–15 sblaCTX-MF-ATGTGCAGYACCAGTAARGTKATGGCR-TGGGTRAARTARGTSACCAGAAYCAGCGGExtension72 °C–30 s593Final Extension72 °C–5 min blaSHVF-AGCCGCTTGAGCAAATTAAACR-ATCCCGCAGATAAATCACCACInitial denaturation 95 °C–5 min 713[29]Denaturation 94 °C–45 s35Annealing53 °C–45 sExtension72 °C–1 minFinal Extension72 °C–10 min blaKPCF-CGTCTAGTTCTGCTGTCTTGR-CTTGTCATCCTTGTTAGGCGInitial denaturation 94 °C–10 min 798[30]Denaturation 94 °C–30 s30Annealing52 °C–40 sExtension72 °C–50 sFinal Extension72 °C–5 min blaNDMF-TCGCATAAAACGCCTCTGR-GAAACTGTCGCACCTCATInitial denaturation 95 °C–6 min 1001[31]Denaturation 95 °C–45 s32Annealing55 °C–45 sExtension72 °C–60 sFinal Extension72 °C–7 min qnrF-ACACAGAACGGCTGGACTGCR-TTCAACGACGTGGAGCTGTInitial denaturation 95 °C–5 min 817[32]Denaturation 95 °C–60 s30Annealing55 °C–60 sExtension68 °C–60 sFinal Extension68 °C–5 min Sul1F-TAGCGAGGGCTTTACTAAGCR-ATTCAGAATGCCGAACACCGInitial denaturation95 °C–5 min 437[33]Denaturation95 °C–1 min35Annealing55 °C–60 sSul2F-CCTGTTTCGTCCGACACAGAR-GAAGCGCAGCCGCAATTCAT956Extension72 °C–1 minFinal Extension72 °C–10 min blaOXA-48F-GCGTGGTTAAGGATGAACACR-CATCAAGTTCAACCCAACCGInitial denaturation 95 °C–5 min 438[34]Denaturation 95 °C–45 s35Annealing60 °C–45 sExtension72 °C–1 minFinal Extension72 °C–8 min Virulence genesentAF: CGTTCGCACTCGACGTGACR: CGAACTGACGGTAACGATCACGInitial denaturation 94 °C–5 min 251[35]Denaturation 94 °C–30 s34Annealing60 °C–30 sExtension72 °C–30 sFinal Extension72 °C–5 min stmPr1F: TGAAAGCAAATGCGCCGTTGR: GTGATGGCGTCGGTGATGTCInitial denaturation 94 °C–5 min 852Denaturation 94 °C–30 s34Annealing60 °C–30 sExtension72 °C–30 sFinal Extension72 °C–5 min hlyIII F: CGTCCATTGCTTCGATCCGTGR: GACGAAGTGGCAGACGCTG Initial denaturation 94 °C–5 min 607Denaturation 94 °C–30 s34Annealing60 °C–30 sExtension72 °C–30 sFinal Extension72 °C–5 min fimH F: GATCCGCCTGAACTGCCAGR: CTGGCAGTTCAGGCGGATC Initial denaturation 94 °C–5 min 576Denaturation 94 °C–30 s34Annealing60 °C–30 sExtension72 °C–30 sFinal Extension72 °C–5 min hgbBF: GGACATCCAGAACATGGGTGCR: GGATCGATCGTGTACGGACCInitial denaturation 94 °C–5 min 1239Denaturation 94 °C–30 s34Annealing60 °C–30 sExtension72 °C–30 sFinal Extension72 °C–5 min virBF: GCATCATGCAGAACGAGCTGR: GACGGCTCGTACTTCTGCACInitial denaturation 95 °C–5 min 1075Denaturation 95 °C–45 s35Annealing60 °C–45 sExtension72 °C–1 minFinal Extension72 °C–8 min frpCF: CCAGTTCAACCTGTCGATGCTGR: CACCGAACAGGTTGTCCCAGInitial denaturation 95 °C–5 min 653Denaturation 95 °C–45 s35Annealing60 °C–45 sExtension72 °C–1 minFinal Extension72 °C–8 min afaDF: GAAGCGCCTGACTGCCTTTTGR: GATCACGTTGTAAGGCCGCCInitial denaturation 95 °C–5 min 328Denaturation 95 °C–45 s35Annealing60 °C–45 sExtension72 °C–1 minFinal Extension72 °C–8 min fhaBF: GTATCGCACAACCGCTTCCAGR: CGTCGTTGATGACCTTCTGCACInitial denaturation 95 °C–5 min 1744Denaturation 95 °C–45 s35Annealing60 °C–45 sExtension72 °C–1 minFinal Extension72 °C–8 min papDF: CACGCGAGTGATCTATCCGGR: GTGATGAAGCGCACCTGGTCInitial denaturation 95 °C–5 min 579Denaturation 95 °C–45 s35Annealing60 °C–45 sExtension72 °C–1 minFinal Extension72 °C–8 min gspDF: GTCGACACCGATATCGGTGGR: GGTAGACCACATGCAGGTTGCInitial denaturation 94 °C–5 min 694Denaturation 94 °C–30 s32Annealing60 °C–15 sExtension72 °C–30 sFinal Extension72 °C–5 min hcpF: GACGGCAACGCGATCAATTACR: GTTCTTGGTTGCACTCCACTGInitial denaturation 94 °C–5 min 201Denaturation 94 °C–30 s32Annealing60 °C–15 sExtension72 °C–30 sFinal Extension72 °C–5 min zotF: GCGTCAGTACACCGATGGTTGR: GCAGGCAGTGTCCAGCATGInitial denaturation 94 °C–5 min 431Denaturation 94 °C–30 s32Annealing60 °C–15 sExtension72 °C–30 sFinal Extension72 °C–5 min plcN1F: GTGACCGATATCGGCCGACR: CTGGAAGTGGCGGTGGAAGInitial denaturation 94 °C–5 min 1779Denaturation 94 °C–30 s34Annealing62 °C–30 sExtension72 °C–30 sFinal Extension72 °C–5 min pilUF: CGACCACCATCGATTTCACTTCGR: GACAGGTCCATCAGCAGCTGInitial denaturation 94 °C–5 min 778Denaturation 94 °C–30 s34Annealing60 °C–30 sExtension72 °C–30 sFinal Extension72 °C–5 min fliCF: CGATCTCCGAGCGCTTCGR: GAACAGCTGGCTGGAGAACGInitial denaturation 94 °C–5 min 296Denaturation 94 °C–30 s34Annealing60 °C–30 sExtension72 °C–30 sFinal Extension72 °C–5 min rmlAF: CTCAGCGTGCTGATGCTGGR: GATGAAGTTGGAGGCTTCCAGCInitial denaturation 94 °C–5 min 600Denaturation 94 °C–30 s34Annealing60 °C–30 sExtension72 °C–30 sFinal Extension72 °C–5 min tpsBF: GTGGACATCGTGATGAAGCGCR: CTTGCCGATGAAGTGACGGTGInitial denaturation 94 °C–5 min 822Denaturation 94 °C–30 s34Annealing54 °C–30 sExtension72 °C–30 sFinal Extension72 °C–5 min motAF: CGTTGGATTCCTGGTCGTCATCR: GAGCCCATGGTGATGACGATGInitial denaturation 94 °C–5 min 558Denaturation 94 °C–30 s34Annealing54 °C–30 sExtension72 °C–30 sFinal Extension72 °C–5 min lktDF: GCACATCCGTGATGCAGTCGR: CGAGATTCTCGTCCTGCATGGInitial denaturation94 °C–5 min 1235Denaturation 94 °C–30 s34Annealing54 °C–30 sExtension72° C–30 sFinal Extension72 °C–5 min
4.2. Screening of the Isolates for the Presence of Antimicrobial Resistance
4.2.1. Antimicrobial Susceptibility Testing (AST)
All isolates underwent antimicrobial sensitivity testing using the Kirby–Bauer disc diffusion method against 12 antibiotics: trimethoprim-sulfamethoxazole (23.75 µg/1.25 µg), enrofloxacin (5 µg), gatifloxacin (5 µg), levofloxacin (5 µg), imipenem (30 µg), piperacillin-tazobactam (100 µg/10 µg), ticarcillin-clavulanate (75 µg/10 µg), ceftriaxone (30 µg), cefpodoxime (10 µg), aztreonam (30 µg), minocycline (30 µg), and tigecycline (15 µg). Isolates were grown in Luria Bertani Broth at 37 °C for 16 h, spread on Mueller–Hinton agar plates, and allowed to dry. Antibiotic discs were placed on the plates within 15 min, and plates were incubated overnight at 37 °C. Zones of inhibition were measured and classified as sensitive, intermediate, or resistant per CLSI guidelines [36].
4.2.2. Assessment of Bacterial Efflux Pump Activity in S. maltophilia
Efflux pump activity in S. maltophilia was assessed using the ethidium bromide agar cartwheel method [16]. Trypticase Soy agar plates with ethidium bromide (0.0–2.5 mg/L) were divided into 12 radial sectors. Bacterial cultures (0.5 McFarland) were swabbed from center to edge and incubated at 37 °C for 16 h. Fluorescence was then observed under a gel documentation system.
4.2.3. Antimicrobial Resistance Genes (ARGs)
Screening of antimicrobial genes in S. maltophilia was carried out by PCR with cyclic conditions as described in Table 3. The ARGs targeted were blaCTX-M and blaTEM [28], blaSHV [29], blaOXA-48 [34], blaKPC [30], blaNDM [31], blaVIM, and blaSIM [27] encoding for β-lactam resistance, qnr [32] encoding for quinolone resistance, and sul-1 and sul-2 [33] encoding for sulphonamide resistance.
4.3. Characterization of the Virulence Properties of S. maltophilia
4.3.1. Evaluation of Biofilm-Forming Ability
Biofilm-forming ability of isolates was assessed by crystal violet staining [37,38]. Cultures (1.0 McFarland, diluted 1:100) were incubated in microtiter plates at 37 °C for 24 h. After washing and fixing, biofilms were stained, destained, and quantified by OD at 492 nm. Biofilm strength was categorized based on OD values as Negative: < Ac; Weak: Ac < A ≤ 2Ac; Moderate: 2Ac < A ≤ 4Ac, and Strong: A > 4Ac, where Ac-OD Value of Negative control; A-OD Value of Sample.
4.3.2. Motility Assay
Swimming motility of S. maltophilia was assessed on modified LB agar (0.3% w/v) plates [20]. In total, 10 µL of cultures (0.5 McFarland) were inoculated at the center and incubated at 37 °C for 24 h. Halo diameters were recorded as high (>10 mm), moderate (>7 to ≤10 mm), or low motility (> 6 to ≤7 mm) [35].
4.3.3. Molecular Characterization of Virulence Genes
The screening of 20 virulence genes in S. maltophilia isolates was conducted through multiplex PCR which included set I (entA, stmPr1, hlyIII, fimH, hgbB), set II (virB, frpC, afaD, fhaB, papD), set III (gspD, hcp, zot), set IV (pilU, fliC, rmlA), set V (tpsB, motA, lktD), and picN1 as described by [35] (Table 3).
4.4. Statistical Analysis
A heatmap with hierarchical clustering was generated to visualize the distribution of antimicrobial resistance, virulence, motility, and biofilm formation in S. maltophilia isolates [39]. Spearman’s correlation between resistance and virulence genes was analyzed in R (Version 4.3.1) using pheatmap and rcorr packages, with significance at p < 0.05. Data visualization employed Origin 2024 (trial version).
5. Conclusions
In summary, our study documented the presence of multidrug-resistant virulent S. maltophilia isolates in dogs with dermatological ailments at the Teaching Veterinary Hospital, Madras Veterinary College, India. It highlights significant genetic diversity among isolates and is the first report of S. maltophilia in dogs from India. The varying profiles of antimicrobial resistance, virulence gene expression, motility, and biofilm-forming ability emphasize the need for vigilant antimicrobial stewardship in veterinary settings. The potential for zoonotic transmission from infected pets necessitates a One Health approach to mitigate risks associated with this opportunistic pathogen and promote health in both humans and animals.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Brooke J.S. Advances in the microbiology of Stenotrophomonas maltophilia Clin. Microbiol. Rev.202134 e 000301910.1128/CMR.00030-1934043457 PMC 8262804 · doi ↗ · pubmed ↗
- 2Gibb J. Wong D.W. Antimicrobial treatment strategies for Stenotrophomonas maltophilia: A focus on novel therapies Antibiotics 202110122610.3390/antibiotics 1010122634680807 PMC 8532924 · doi ↗ · pubmed ↗
- 3Mikhailovich V. Heydarov R. Zimenkov D. Chebotar I. Stenotrophomonas maltophilia virulence: A current view Front. Microbiol.202415138563110.3389/fmicb.2024.138563138741741 PMC 11089167 · doi ↗ · pubmed ↗
- 4Bhaumik R. Aungkur N.Z. Anderson G.G. A guide to Stenotrophomonas maltophilia virulence capabilities, as we currently understand them Front. Cell. Infect. Microbiol.202313132285310.3389/fcimb.2023.132285338274738 PMC 10808757 · doi ↗ · pubmed ↗
- 5Albini S. Abril C. Franchini M. Hüssy D. Filioussis G. Stenotrophomonas maltophilia isolated from the airways of animals with chronic respiratory disease Schweiz. Arch. Tierheilkd.200915132332810.1024/0036-7281.151.7.32319565454 · doi ↗ · pubmed ↗
- 6Kralova-Kovarikova S. Husnik R. Honzak D. Kohout P. Fictum P. Stenotrophomonas maltophilia urinary tract infections in three dogs: A case report Vet. Med.20125738038310.17221/6268-VETMED · doi ↗
- 7Thenissery A. Chandran R. Ravindran R. Ravishankar C. Piriyath J.H. Isolation and identification of emergent multidrug resistant Stenotrophomonas maltophilia from skin ulcers of Sarcoptes infested pigs Vet. Rec. Case Rep.202210 e 31210.1002/vrc 2.312 · doi ↗
- 8Jayol A. Corlouer C. Haenni M. Darty M. Maillard K. Desroches M. Lamy B. Jumas-Bilak E. Madec J.Y. Decousser J.W. Are animals a source of Stenotrophomonas maltophilia in human infections? Contributions of a nationwide molecular study Eur. J. Clin. Microbiol. Infect. Dis.2018371039104510.1007/s 10096-018-3203-029488120 · doi ↗ · pubmed ↗